基于糞便DNA的賀蘭山巖羊親權鑒定和婚配制研究
2019-12-31 08:02:58祝招玲劉振生張致榮姚緒新韓興志王繼飛滕麗微
生態學報 2019年22期
祝招玲,劉振生,高 惠,張致榮,姚緒新,劉 鵬,韓興志,王繼飛,4,滕麗微,*
1 東北林業大學野生動物與自然保護地學院,哈爾濱 150040 2 佳木斯大學經濟與管理學院,佳木斯 154007 3 國家林業局野生動物保護學重點開放實驗室,哈爾濱 150040 4 寧夏賀蘭山國家級自然保護區管理局,銀川 750021
巖羊(Pseudoisnayaur)屬偶蹄目牛科羊亞科巖羊屬,是典型的高山動物,棲息于高原丘原和高山裸巖與山谷間的草地,被世界自然保護聯盟物種生存委員會(IUCN/SSC)收錄為低危種,在我國重點保護野生動物名錄中,被列為國家二級保護動物。目前對巖羊的研究主要集中在種群生態學[1]、行為生態學[2]和種群遺傳學[3]等領域。伴隨著非損傷性取樣技術的應用,利用糞便DNA結合微衛星標記對羊亞科動物進行個體識別、親權鑒定和遺傳多樣性等方面開展了豐富的研究[4-5],同時對動物的婚配制也進行了一定的研究,如對倭黑猩猩(Panpaniscus)[6]、黑麂(Muntiacuscrinifrons)[7]、小麂(Muntiacusreevesi)[8]和黑犀牛(Dicerosbicornis)[9]等婚配制的研究,這使得分子生物學手段成為研究巖羊親權鑒定和婚配制的新視角。
本研究采用非損傷性取樣技術,通過微衛星標記進行個體識別和親權鑒定,并分析雌雄巖羊的交配策略,探討巖羊的婚配制度,為進一步開展巖羊種群分子行為學研究而完成基礎性技術研發工作,同時也為該物種的保護、利用和管理提供科學依據。
1 材料和方法
1.1 研究地概況及樣本采集
本研究采樣地點為賀蘭山巖畫風景區(N 38°44′,E 106°01′),位于賀蘭山東麓,北起拜寺口,南至三關口,南北長30 km,東西寬4 km,海拔1448 m,是典型的大陸性氣候特征,全年干旱少雨,年均降水量200—400 mm之間,年平均蒸發量2000 mm[1]。植被以山楊(Populusdavidiana)、灰榆(Ulmuspumila)、酸棗(Zizyphusjujubevar.spinosa)、冰草(Agropyroncristatum)、針茅(Stipacapillata)、早熟禾(Poaannua)等為主。此處巖羊數量較多,具有一定的代表性。在2017年7—8月和11—12月采用樣線法進行巖羊新鮮糞便樣本的采集,依據賀蘭山實際溝道走向和地形地貌特征布設了7條樣線,樣線長度為2—6 km[10]。在樣線行走過程中,搜尋巖羊的新鮮糞便,若發現巖羊實體,等待巖羊自行離開后,再進行新鮮糞便樣本的收集。為了避免樣本交叉污染,戴口罩和一次性PE手套,用已消毒的鑷子采集糞便樣本,在新鮮的糞便中選取20粒左右放入50 ml的管中,貼上標簽,每采集完一次樣本,更換PE手套和鑷子。樣本采集后帶回實驗室,放入-80℃冰箱保存。兩次共獲得369份巖羊糞便樣本,巖羊皮張1份。
1.2 基因組DNA的提取和PCR擴增
1.2.1DNA提取和質量檢測
巖羊糞便用試劑盒(QIAamp Fast DNA Stool Mini Kit)提取DNA,巖羊皮張用血液/細胞/組織基因組DNA提取試劑盒提取DNA,按照說明書進行操作。1.0%瓊脂糖凝膠電泳檢測巖羊糞便DNA質量。
1.2.2微衛星位點的選取和PCR擴增
本研究通過查閱文獻得到31對賀蘭山巖羊的微衛星位點[5,11-12],并委托生工生物工程(上海)股份有限公司(Sangon)合成引物,經多次實驗后篩選出擴增穩定且多態信息含量高的10對引物(見表1),在上游引物5′端分別標記FAM、HEX、TAMRA三色熒光重新合成。PCR反應體系:總體積12 μL,2×PCR Buffer for KOD FX Neo 6 μL;2 mM dNTPs 2.4 μL;10 pmol/μL的上、下游引物各0.36 μL;100 ng/μL的DNA模板1.5 μL;1 U/50 μL KOD FX Neo 0.24 μL;ddH2O 1.14 μL。PCR反應程序:94℃預變性2 min,98℃變性10 s,54℃—60℃退火30 s,68℃延伸30 s,反應35個循環,最后68℃延伸7 min。
實驗采用Huber等[13]提供的2對引物SRY12、BMCl009和牙釉蛋白基因PCR法[14]進行性別鑒定(見表2)。對實驗的所有樣本進行3次PCR 擴增,并用實驗室已知性別的巖羊皮張DNA樣本作為對照。性別鑒定多重PCR擴增:出現雙條帶(180 bp和300 bp)判定為雄性;出現一條帶(300 bp)則判定為雌性。牙釉蛋白基因PCR擴增法:出現雙條帶(218 bp和263 bp)判定為雄性;出現一條帶(263bp)則判定為雌性。最后比較兩套PCR體系的結果來判斷樣本的性別。PCR反應體系同上。
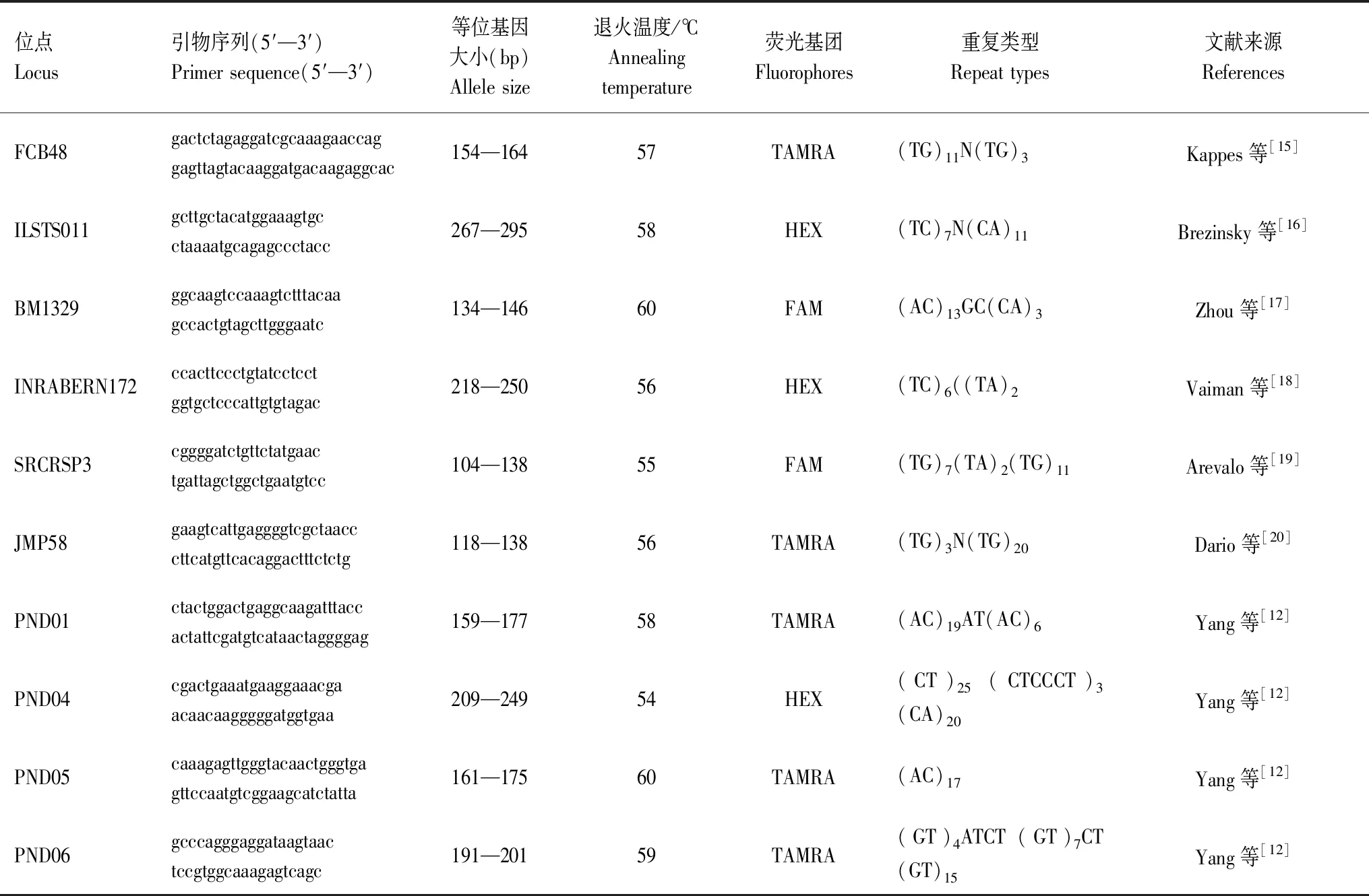
表1 10對微衛星引物序列Table 1 Ten microsatellite primer sequences

表2 性別鑒定引物序列Table 2 Primer sequences of sex determination
1.3 數據分析
微衛星位點的擴增產物送上海生工用ABI測序測儀(3730xl DNA Analyzer)進行分型,用Excel整理微衛星數據,之后用Cervus 3.0[15](Allele Frequency Analysis)功能,計算等位基因數目、雜合度、多態信息含量、親權排除概率、無效等位基因頻率及Hardy-Weinberg平衡檢測,同時對巖羊進行個體識別,鑒定個體時設定以下原則:所有微衛星座位上的基因型都相同或只有一個座位上的一個等位基因存在差異則為同一個個體[16]。運行Cervus 3.0的 Simulation of Parentage Analysis 和 Parentage A nalysis 功能,由于父本、母本均未知,所以將所有雄性和雌性分別作為候選父本和候選母本,取樣概率均設為1.0(取樣概率為候選父占取樣中雄性比例及候選母占取樣中雌性比例),其他選項均為系統默認值,進行親權鑒定。實驗中為了保證鑒定結果的準確性,所有位點全匹配的才確認親子關系。使用Kingroup V2[17]計算個體間的親緣系數(r),作為親權鑒定的補充,親緣系數范圍在-1到1之間,表示親緣相近的程度。理論上,當種群處于Hardy-Weinberg平衡時,0.5≤r<1時為同父同母的兄弟姐妹關系,0.25≤r<0.5時為同父異母或同母異父的兄弟姐妹關系[18]。通過Genepop[19]對10個位點組成的45組進行連鎖不平衡檢驗,采用馬爾科夫鏈法(Markov chain method)分析各位點間的連鎖關系,參數均設為5000 dememorization、100 bateh和5000 iteration[20]得到無偏估計P值,當P<0.05表明連鎖不平衡具有顯著性。運用POPGENE 32[21]計算種群的近交系數Fis值,其值范圍為-1—1,Fis值為顯著正值時,表示種群內存在較嚴重的近交現象,Fis值為顯著負值時,表示種群內存在遠交[22]。
2 結果
2.1 個體識別和親權鑒定
2.1.1基因組提取、性別鑒定和個體識別
369份巖羊糞便全部成功提取到DNA,經1.0%瓊脂糖凝膠電泳檢測,巖羊糞便中提取的DNA片段完整,質量較高,可以直接用于PCR擴增,共鑒定出282只巖羊個體,其中雄性152只,雌性130只。性別鑒定部分結果見圖1、圖2。
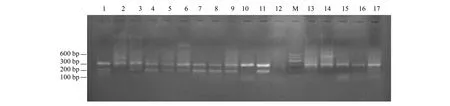
圖1 巖羊SRY12和BMCl009位點的擴增結果Fig.1 The amplification results of SRY12 and BMCl009 loci of Pseudois nayaur1—9,13—17:巖羊糞便DNA;10:雌性巖羊皮張DNA;11:雄性巖羊皮張DNA;12:陰性對照
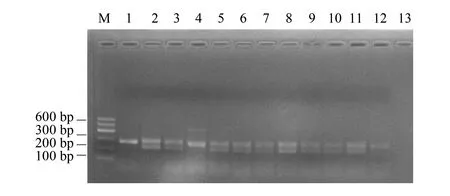
圖2 牙釉蛋白基因(AMELX和AMELY)擴増結果Fig.2 The amplification results of amelogenin gene (AMELX &AMELY)1:雌性巖羊皮張DNA;2:雄性巖羊皮張DNA;3—12:巖羊糞便DNA;13:空白對照
2.1.2微衛星位點的親權排除率
10個微衛星位點共檢測到101個等位基因,平均等位基因(Na)為10.1,PIC在0.452—0.825之間,平均多態信息0.6129,為高度多態位點(PIC﹥0.5),平均觀測雜合度(HObs)為0.7737,平均期望雜合度(HExp)為0.6512。Fis平均值為-0.1836。無效等位基因(F(Null))頻率極低,平均值為-0.0790。Hardy-Weinberg平衡檢測顯示,8個位點完全符合哈溫平衡,位點SRCRSP3和ILSTS011則顯著偏離平衡。累積單親排除概率為0.9615(累積單親排除概率是根據多個位點累積排除任意一個可疑父親或母親為假父親或假母親的概率),累積雙親排除概率為0.9975(累積雙親排除概率是根據多個位點累積排除任意兩個可疑父親和母親為假父親和假母親的概率)(見表3)。
通過Genepop對10個位點組成的45組進行連鎖不平衡檢驗(見表4),由此得到11組連鎖不平衡具有顯著性的組合。經Bonferroni[23]校正后,仍連鎖不平衡(P<0.05/45=0.0011)的組合有2組,PND01與INRABERN172,FCB48與PND05。
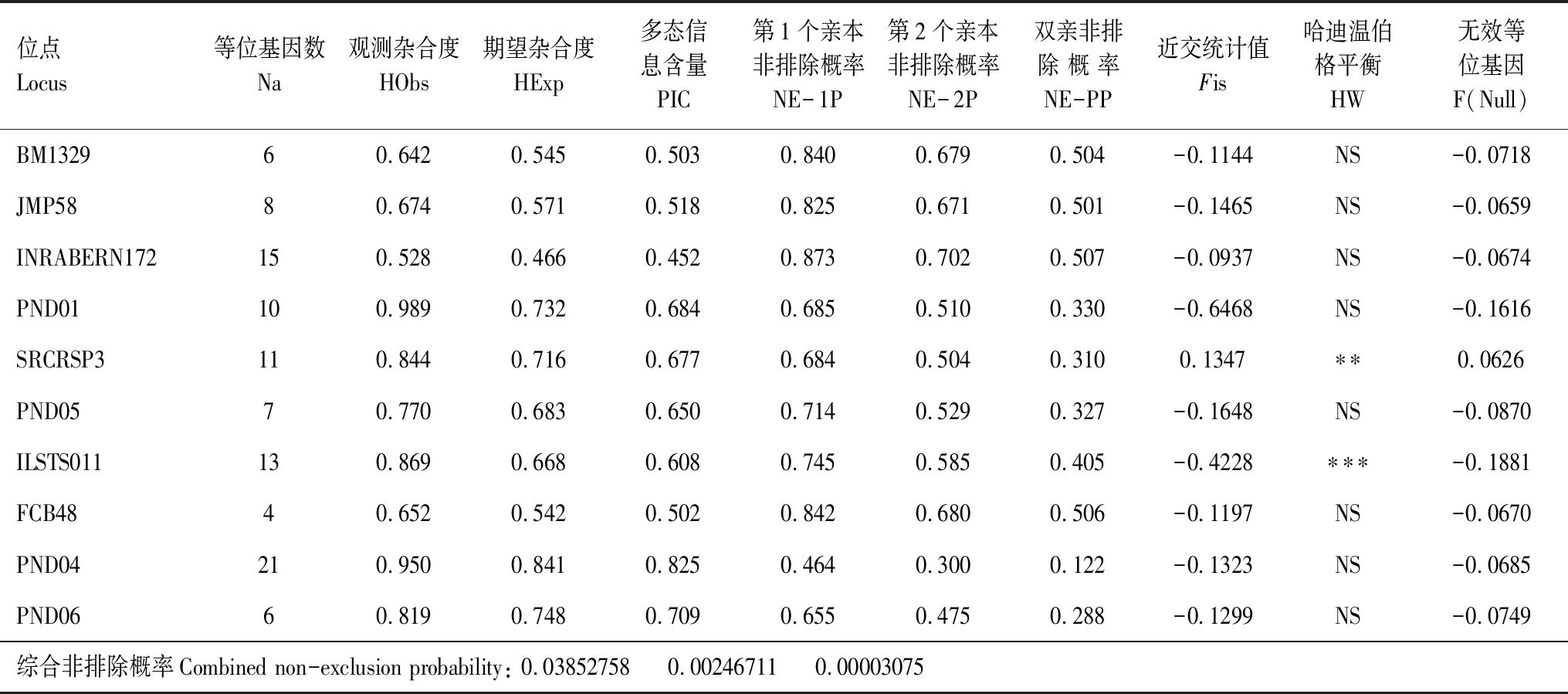
表3 10個微衛星位點的親權排除概率Table 3 Parental exclusion probability of 10 microsatellite locis
Na:位點等位基因數,allele number of 1oci;HObs:位點觀測雜合度,observed heterozygosity;HExp:位點期望雜合度,Expected heterozygosity;PIC:位點多態信息含量,Polymorphic information content;NE-1P:第1個親本非排除概率,Combined non-exclusion probability (first parent);NE-2P:第2個親本非排除概率,Combined non-exclusion probability (second parent);NE-PP:雙親非排除概率,Combined non-exclusion probability (parent pair);Fis:近交系數的F-統計量檢測值,inbreeding coefficient withF-statistics;HW:哈迪溫伯格平衡Hardy-Weinberg equilibrium;F(Null):無效等位基因頻率,frequency with null;NS:不顯著偏離,no significant deviation

表4 10個微衛星位點的連鎖不平衡檢驗結果Table 4 Linkage disequilibrium test result of 10 microsatellites locis
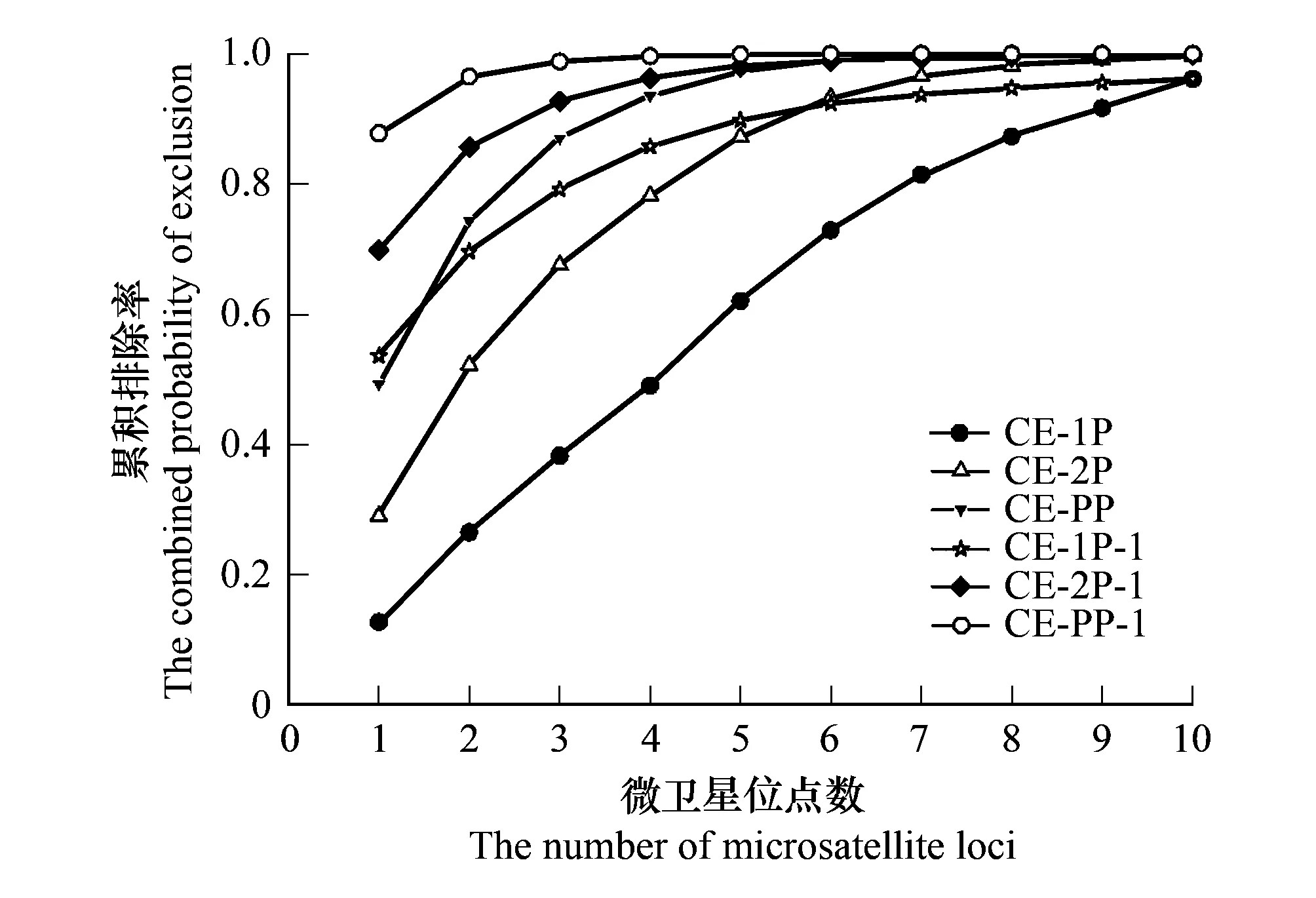
圖3 微衛星位點數與累積排除率之間的關系Fig.3 Relationship between the combined probability of exclusion and the number of microsatellite lociCE-1P,CE-2P,CE-PP:PIC由低到高排列的累積排除概率;CE-1P-1,CE-2P-1,CE-PP-1:PIC由高到低排列的累積排除概率
按照多態信息含量(PIC)由低到高和由高到低的順序依次增加微衛星位點數進行累積排除率分析,結果由圖3可知,隨著微衛星位點數的增加,累積排除率逐漸增加,當微衛星位點達到7個(PIC由高到低)和9個(PIC由低到高)時,累積親權排除概率大于0.9900,說明所篩選的10個微衛星衛點能進行準確的親權判定。
2.1.3親權鑒定
運用 Cervus3.0 軟件的Parent pair(Sexes known)功能進行親權鑒定,共鑒定出父-母-子7對,父-子21對,母-子19對。28對父-子的親緣系數(r)在0.5000—0.8462之間,26對母-子的親緣系數(r)在0.5000—0.8000之間,所有父子間、母子間、同父同母個體間的親緣系數平均分別為0.6164、0.6251和0.5426。實驗中運用10個微衛星位點進行親子鑒定,親權排除概率為99.75%。
2.2 婚配制度
由表5、表6可知,X162(♂)、X173(♂)各自3個后代間的親緣系數在0.25≤r<0.5之間,說明X162(♂)、X173(♂)均與3個雌性交配過;D31(♀)、D33(♀)、D18(♀)、X168(♀)、D148(♀)各自后代間的親緣系數在0.5≤r<1之間,說明它們只與1個雄性交配過;D73(♀)、D78(♀)、D89(♀)各自后代間的親緣系數在0.25≤r<0.5之間,說明它們與2個雄性交配過;X163(♂)、X3(♂)、D145(♂)、X160(♂)、X112(♂)、D15(♂)各自后代間的親緣系數在0.25≤r<0.5之間,說明它們與2個雌交配過;D45(♂)、D70(♂)各自后代間的親緣系數在0.5≤r<1之間,說明它們只與1個雌性交配過。研究中多次出現1個雄性與2個或2個以上的雌性交配并產下后代,可知巖羊的婚配制屬于一雄多雌。
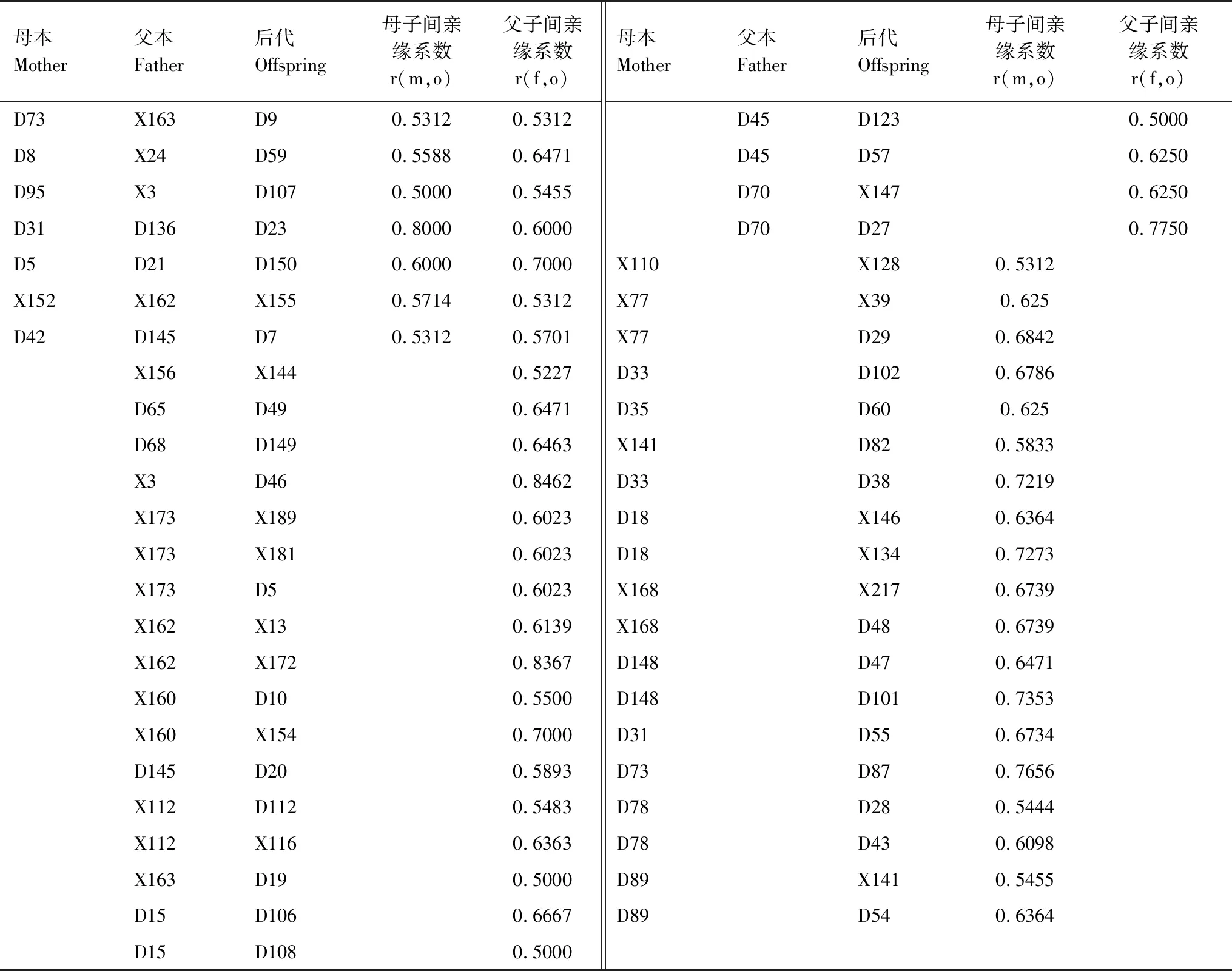
表5 巖羊的親子關系和親緣系數Table 5 The coefficient of parentage and genetic relationship in Pseudois nayaur
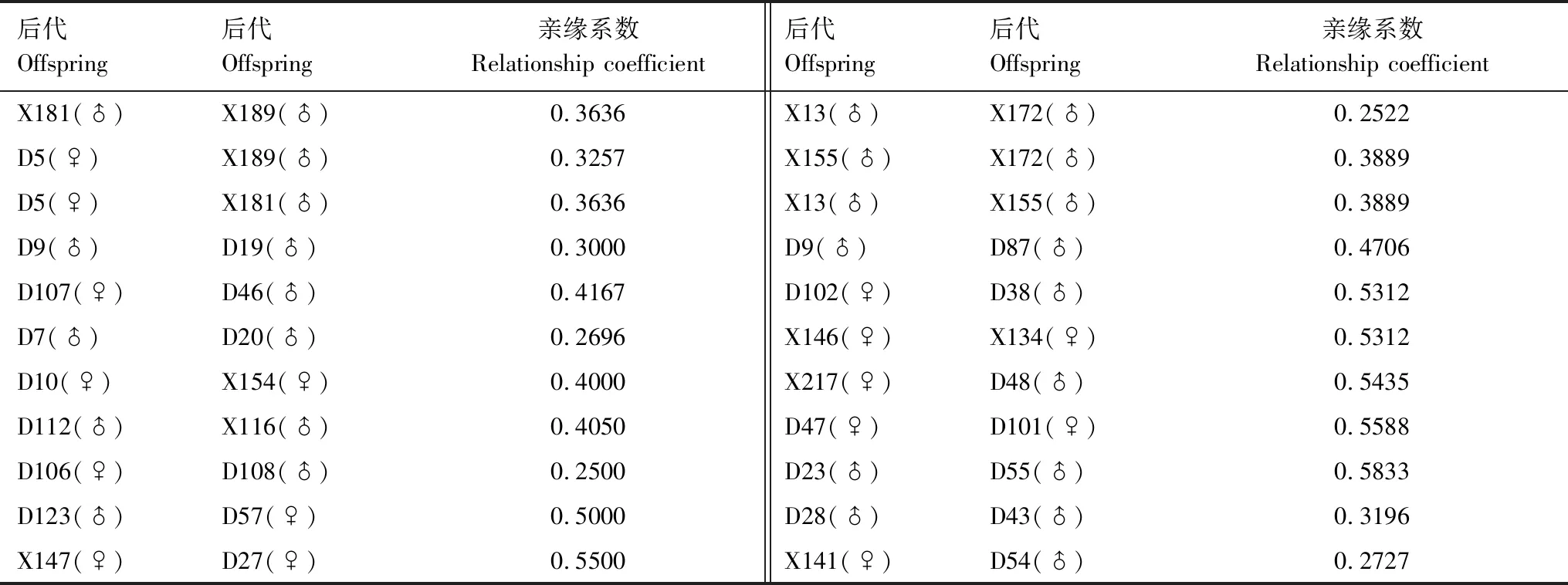
表6 后代之間的親緣系數Table 6 The relationship coefficient among the offsprings
3 討論
3.1 親權鑒定
目前普遍采用STR來鑒定個體間的親權關系,其排除非親本能力依賴于微衛星位點數和等位基因的多樣性[24]。一般8—10個及以上微衛星位點即可確定較高的親權鑒定準確率[15]。本研究選用10個高度多態性(PIC>0.5)微衛星位點(見表3),進行親權鑒定是可行的。
無效等位基因是影響親權鑒定準確性的基礎因素,本研究使用的10個微衛星位點,雜合子豐富,無效等位基因頻率極低,而Cervus3.0軟件使用似然率(似然率即某一可疑父親的似然值相對于理想父親似然值的比)容許這些錯誤的存在,在統計處理上可以有效降低錯配的敏感性,使親權鑒定結果更加科學化,同時還可以充分使用所占有的遺傳資料[25-26]。Hardy-Weinberg平衡檢驗結果發現共有2個微衛星位點出現偏離平衡的現象,其余8個位點處于平衡狀態,Cervus 中一兩個座位的Hardy-Weinberg不平衡并不影響親權分析中似然比的計算。Fis為負值,說明種群內不存在近交。賀蘭山巖羊是生活在特殊環境下的一個邊緣種群,集小群,隨著季節的變化,其集群類型、集群大小均會發生一定的變化,這種邊緣種群在適應性進化過程中可能會導致不平衡選擇,使得基因組中的某些位點偏離Hardy-Weinberg平衡[5]。10個位點經校正后有2組連鎖不平衡具有顯著性,可能是因為無效等位基因的存在導致了這些組合連鎖不平衡。
一般情況下,參照國際常用標準,RCP>95%,傾向有親子關系;RCP>99.73%,認定有親子關系[25];對于人類的親權判定,累積排除概率99.0%—99.8%即可認定為極其成立的親子關系[27];王躍峰利用5個微衛星位點對白頭葉猴進行個體識別和親緣鑒定,得到累積非父排除概率為99.70%[28];程文科對144頭豬進行個體親子鑒定時得到累積非父排除率為98.35%,與系譜記錄結果完全相同[4];郭立平利用8個微衛星位點在3種情況下為西門塔爾牛進行親子鑒定,累積排除概率分別為99%、99.95%和99.99%,鑒定結果與系譜一致[29]。
實驗中親權排除概率為99.75%,并用Kingroup V2確定親緣系數,進一步驗證了親權鑒定的準確性。因此,實驗中采用的10對微衛星位點完全可以達到親子鑒定的要求。
3.2 巖羊的婚配制度
陳小南[7]用70個黑麂個體糞便樣本、任鵬[8]用177個小麂個體糞便樣本、Garnier[9]用35只野生黑犀牛的糞便樣本在親權鑒定的基礎上,通過分析雌雄間的交配關系確定婚配制。本研究利用282只巖羊個體糞便樣本,根據巖羊雌雄間的交配關系研究婚配制度,在樣本數量上完全滿足實驗要求。
通常把婚配制度分為單配制(monogamy)和多配制(polygamy),多配制又包括一雄多雌(polygyny)、一雌多雄(polyandry)和混交制(promiscuity),動物選擇哪一種婚配制度受個體之間的相互作用和對外界環境適應的影響[30]。研究動物的婚配制度,需要注意在一個繁殖季節里,個體以什么樣的方式獲得配偶,獲得配偶的數量以及雌雄個體的育幼行為等因素[31]。
新疆天山北山羊(Caprasibirica)傾向于組成“一雄多雌”的小群完成交配[32]。捻角山羊(Caprafalconeri)是一雄多雌的交配系統[33]。赤羊(Ovisvignei)在交配時期,雄性赤羊會由獨居轉為與4—5頭雌性赤羊居住,是一雄多雌的交配系統[33]。盤羊(Ovisammon)在交配期間,活動區域選擇地勢平坦、避風暖和的低海拔區域,雄羊和雌羊開始集群,5—6 只雄盤羊和數十只雌盤羊一起活動,雄性盤羊間爭偶激烈,相互劇烈格斗,勝者獲取這群盤羊的交配權,交配季節結束后,雌雄又分開活動[34]。叉角羚(Antilocapraamericana)在交配季節占據了不設防的領地,雄性保護雌性的后宮[35]。阿爾卑斯羱羊(Capraibex)似乎能夠通過堅持它們之間預先建立的穩定的優勢關系來減少因爭斗而消耗的能量,年長的雄性通常采取撫育策略壟斷與雌性交配的機會,而年輕的、處于從屬地位的雄性通過悄悄接近雌性的策略獲得交配,但成功率總體上較低[36]。賀蘭山巖羊集小群,發情期雌雄群、混合群和獨羊比例顯著上升,雌性巖羊相對分散,部分雄性巖羊在不同群之間遷移尋找發情的雌性巖羊,并通過爭斗獲得優勢地位,進一步獲得交配權,盡可能增加繁殖機會[37],這一現象說明了實驗中1只雌性與2只雄性巖羊交配并產下后代的現象是在雌性不同的繁殖期之間,同時也證實了實驗中1只雄性與多只雌性巖羊交配并產下后代的結果,推測巖羊是一雄多雌的婚配制度。
哺乳動物具有較長的妊娠期和哺乳期,雄性動物在此期間不能提供更多的幫助,而是傾向于盡可能多地與多個雌性交配,這是提高自身適合度的一種進化選擇[38]。巖羊發情交配期在每年的11月末到12月,在12月中旬時達到高峰,母羊經過大約160 d的妊娠期后在次年5月中旬到6月中旬產仔,哺乳期3個月[39],這期間雄性巖羊不參與育幼活動,由雌性巖羊哺育下一代,觀察發現母仔群出現的頻率在一年的四個季節中均最高[10]。由于巖羊雌雄兩性在婚配中投資的不平衡性,促使雄性巖羊有更多的時間和精力參與到繁殖行為中,獲得與更多雌性交配的機會,這也驗證了實驗研究的結果,雄性巖羊與多個雌性巖羊交配并產下后代,表明巖羊是一雄多雌制的婚配制度。
實驗從分子生物學角度對賀蘭山巖羊進行親權鑒定和婚配制研究,填補了這一領域的研究空白,并為進一步開展巖羊種群分子行為學研究積累基礎資料。同時對采用非損傷性取樣技術研究有蹄類動物的個體識別、性別鑒定、親權鑒定和婚配制等具有重要意義,特別是對瀕危有蹄類動物的保護和利用提供了新的研究方法和視角。此外,希望通過實驗的研究成果,可以為有關部門和科研單位提供有效數據,促進形成一套更加完整的賀蘭山巖羊保護、利用和管理體系。
致謝:寧夏賀蘭山國家級自然保護區管理局胡天華高級工程師在野外調查中給予幫助,特此致謝。
