4-羥基他莫昔芬依賴的內含肽調控綠色熒光蛋白功能研究
2019-12-25 08:43:50趙拯浩王步森王玉東朱丹妮董銘心侯利華陳薇
生物技術通訊 2019年5期
關鍵詞:效率
趙拯浩,王步森,王玉東,朱丹妮,董銘心,侯利華,陳薇
軍事科學院軍事醫(yī)學研究院生物工程研究所,北京100071
內含肽(intein)是位于宿主蛋白質中的一段插入序列。經典內含肽結構由催化蛋白剪接的N端和C 端剪接結構域以及位于中央的歸巢內切酶結構域構成一個馬蹄鐵狀的結構[1],其對應的核苷酸序列嵌合在宿主蛋白對應的核酸序列中,與宿主蛋白基因存在于同一開放讀框內,并與宿主蛋白基因進行同步轉錄和翻譯。當翻譯形成蛋白質前體后,內含肽從蛋白質前體中切除,從而產生成熟的宿主蛋白[2-3]。在此過程中,內含肽序列被準確切除,而其兩側被稱為外顯肽(extein)的2 段宿主蛋白質序列則通過正常的肽鍵連接起來。內含肽在多種環(huán)境下形成和清除肽鍵的能力,使得其發(fā)展成為分子生物學強有力的工具,應用于蛋白質修飾、純化、生物傳感器等領域[4-6]。
天然內含肽的自剪接不需要ATP、酶及小分子的參與,但這種自剪接的功能是不利于外部調控的。Liu 等[7]基于天然內含肽結構,開發(fā)了一種基于內含肽的分子開關,這個分子開關可將小分子的結合轉化成目標蛋白質的激活。這種可以利用小分子4-羥基他莫昔芬(4-hydroxytamoxifen,4-HT)調控的內含肽最早來源于結核分枝桿菌RecA 內含肽結構中的部分序列,在融合了304~551 氨基酸殘基的人雌激素受體配體結合域(estrogen receptor-ligand binding domain,ER-LBD)后,得到了含有424 個氨基酸殘基的內含肽融合蛋白。為了使這一內含肽融合蛋白在哺乳動物細胞中也能高效應用,Peck 等[8]通過反復誘變和選擇,產生了高度依賴4-HT 的具有強剪接活性的內含肽,得到一套低背景、快速剪切、適用哺乳動物細胞的內含肽系統(tǒng)。
目前國外文獻報道利用這種分子開關能夠實現對包括Cas9[9-10]、增強型綠色熒光蛋白(eGFP)[7]和卡那霉素抗性蛋白[11]在內的多種蛋白的調控作用,國內尚無利用這種分子開關的相關報道。本實驗旨在驗證小分子調控的內含肽剪接技術在eGFP 上應用的效率,針對eGFP 的結構與功能在其基因上篩選出剪接效率更高的位點,并基于篩選結果,摸索小分子的最適工作條件。
1 材料與方法
1.1 材料
pVax-Intein 質粒由上海生工公司合成;質粒提取試劑盒(Promega,A1223);膠回收試劑盒(TaKaRa,9762);DNA 片段回收試劑盒(TaKaRa,9761);PremixTaq預混液(TaKaRa,RR902);2×Q5 PCR 聚合酶(NEB,M0494S);高保真DNA 組裝預混液(NEB,E2621L);六孔細胞培養(yǎng)板(Corning);凝膠成像儀(Clinx Science,GenoSens 1560);超微量分光光度計(GE,NanoVue Plus);PCR 擴增儀(Eppendorf AG 22331 Hamburg);熒光顯微鏡(BioTek,CY71V);流式細胞儀(BD FACSCantoⅡ);電熱恒溫培養(yǎng)箱(泰斯特,DH5000BⅡ);恒溫振蕩器(Crystal,IS-RDV1)。
1.2 擴增內含肽片段與線性化載體pMD-eGFP
根據改構的4-HT 可調控的內含肽與eGFP 基因(GenBank:NC025025.1)序列[8],用Vector NTI 設計PCR 引物(表1)。以Vax-Intein、pMD-eGFP 質粒(實驗室保存)為模板進行PCR 擴增(擴增條件:94℃5 min;94℃30 s,56℃30 s,72℃30 s/kb,32 個循環(huán);72℃5 min)。50 μL 擴增體系包括模板10 ng,上、下游引物各0.5 μmol/L,2×Q5熱啟動PCR 聚合酶25 μL,加22 μL 超純水補齊50 μL。PCR 結束后4℃保存產物,行1%瓊脂糖凝膠電泳,電泳畢,用凝膠成像系統(tǒng)對膠進行觀察,對符合預期的條帶用膠回收試劑盒回收。
1.3 重組載體pMD-eGFP-Intein 的構建和陽性克隆鑒定
將1.2 中回收的內含肽片段與線性化載體片段引物以摩爾比2∶1 的比例總計3 μL 加入0.25 mL EP 管中,添加NEBuilder 2×Mix 5 μL,去離子水補齊10 μL,混勻后在50℃金屬浴上重組反應15 min。反應結束后,立即放入冰中冷卻,后放入4℃暫存。將重組質粒轉入TOP10 感受態(tài)大腸桿菌,菌液涂布于含50 μg/mg 氨芐青霉素的LB固體培養(yǎng)基上,37℃培養(yǎng)過夜,從轉化平板中挑取單菌落各4 個,劃線培養(yǎng),并進行菌落PCR,引物為InteinF/R。PCR 反應體系(20 μL)包括模板(菌落),正、反向引物各0.4 μL,PremixTaq預混液10 μL,加水補齊20 μL。擴增條件:94℃ 5 min;94℃ 30 s,56℃ 30 s,72℃ 2 min,32 循環(huán);72℃ 5 min。用1%瓊脂糖凝膠電泳檢測。從平板上挑取鑒定陽性菌落在5 mL 含氨芐青霉素的LB 培養(yǎng)基中37℃振蕩培養(yǎng)14~16 h,提取質粒后測序確定目的序列。
1.4 質粒轉染HEK293 細胞系
293 T 細胞以3.5×105~5×105/孔接種至12 孔板,于37℃、5% CO2細胞培養(yǎng)箱中培育14~16 h,使細胞密度達到60%~70%。在1.5 mL 離心管中加入100 μL Opti-MEM 培養(yǎng)基,隨后按每孔1 μg質粒、2 μL Turbofect 轉染試劑在離心管中配制轉染體系,混勻后室溫靜置15 min,滴加至12 孔板中,輕搖混勻。在實驗組中加入4-HT 的DMSO溶液1 μL,終濃度為1 μmol/L,在對照組中加入1 μL DMSO。
1.5 流式細胞術檢測細胞熒光強度
轉染36 h 后,移除1.4 中的細胞上清,每孔用1 mL PBS 漂洗1 次后加入300 μL 1%胰酶,37℃孵育2 min。用1 mL PBS 將細胞吹打重懸并轉移至流式管中,4℃、600 r/min 離心5 min。棄上清,重新用3 mL PBS 重懸細胞,再次4℃、600 r/min 離心5 min,棄上清,用150 μL PBS 重懸細胞,流式上機檢測FITC 值。
1.6 統(tǒng)計學處理
采用SPSS 19.0 軟件進行數據分析。采用t檢驗分析比較添加小分子的實驗組與對照組的熒光強度,P<0.05 為差異有統(tǒng)計學差異,P<0.01為差異具有統(tǒng)計學顯著差異。
1.7 4-HT 最適濃度的探究
質粒轉染細胞步驟同1.4。轉染細胞后,分別在實驗組中添加梯度稀釋的4-HT,使4-HT 在培養(yǎng)基中的終濃度分別為16、8、4、2、1、2-1、2-2、2-3、2-4、2-5、2-7、2-9、2-11、2-13、2-15μmol/L。后續(xù)檢測及統(tǒng)計學處理同1.5 和1.6。
2 結果
2.1 pMD-eGFP-Intein 的構建
eGFP 能夠在藍色光的激發(fā)下發(fā)出綠色熒光。eGFP 的發(fā)光依賴于其緊密的β桶結構,在內含肽替換位點的選取上主要考慮4-HT 容易接觸到的β桶的外側以及β桶兩端的無規(guī)卷曲區(qū)域,期望能夠實現內含肽在剪接前因破壞了eGFP 發(fā)色團共軛結構而使eGFP 不能發(fā)出熒光,而內含肽發(fā)生剪接后eGFP 發(fā)色團共軛結構得到恢復,可以正常發(fā)光,從而實現小分子對eGFP 蛋白功能的調控。由于內含肽的剪接會殘留一個半胱氨酸殘基,因此選取半胱氨酸以及與半胱氨酸結構相似的丙氨酸、絲氨酸和蘇氨酸。在eGFP 蛋白上選取C49、C71、S87、R109、A111、S148、T231 共7個位點插入內含肽。所選取的位點在eGFP 基因組中的位置以及在eGFP 結構上的位置參見圖1。在選取的位點處通過反向PCR 對質粒進行線性化,與PCR 得到的內含肽片段進行同源重組,測序分析后,獲得序列正確的7 個pMD-eGFP-Intein 質粒。
2.2 內含肽對eGFP 蛋白的調控
293 T 細胞分別轉染7 個不同位點嵌入內含肽的重組eGFP 質粒36 h 后,未添加4-HT 的對照組均未檢測到顯著的綠色熒光,說明內含肽的插入影響了eGFP 的正常結構,導致無法激發(fā)綠色熒光。在7 個添加了4-HT 的試驗組中,S87、S148、R109 組可見顯著的熒光恢復(圖2)。在這3 組中,S148 位點插入的eGFP 重組質粒轉染后熒光恢復強度最高,S87 次之,R109 最低。其中,R109是文獻中已報道的內含肽調節(jié)eGFP 發(fā)光的剪接位點[7],能夠觀測到熒光的部分恢復,但恢復效率不高,其他插入位點在熒光顯微鏡下未見熒光。
轉染36 h 后收集細胞,采用流式細胞術定量檢測細胞中eGFP 的熒光強度。結果顯示(圖3),A111、S148、S87、C71、R109 組添加4-HT 后,熒光強度強于未添加4-HT 的對照組,具有統(tǒng)計學差異(P<0.05)。其中S87、S148、R109 組的熒光強度顯著強于對照組(P<0.01),這3 組相較于陽性對照,加入4-HT 后熒光強度恢復率分別為31%、9%和5%(圖3),S148 組熒光恢復率最高,而R109 組的熒光恢復率較低,這與熒光觀測結果相吻合。未加4-HT 的對照組,熒光強度僅為陽性對照組的6%、3%和2%。同時也發(fā)現,剪接效率較高的位點,如S87、S148,其本底值(未添加4-HT 的對照組的熒光強度)也會略高。通過比較有/無4-HT 的熒光比值,可以看出S148、S87 的剪切效率分別為R109 的8 倍和2 倍。

圖1 pMD-eGFP-Intein 的構建
2.3 4-HT 最適濃度的探究
選擇剪接效率最高的S148 位點進行實驗,探究該位點對應的4-HT 最適濃度。將12 孔板中4-HT 的濃度從16 μmol/L 梯度稀釋至2-15μmol/L,在質粒轉染細胞36 h 后流式檢測熒光強度,發(fā)現S148 位點的內含肽剪接效率是高度劑量依賴的,濃度低于4 μmol/L 時,剪接效率與4-HT 的濃度正相關,最適濃度定為4 μmol/L(圖4)。
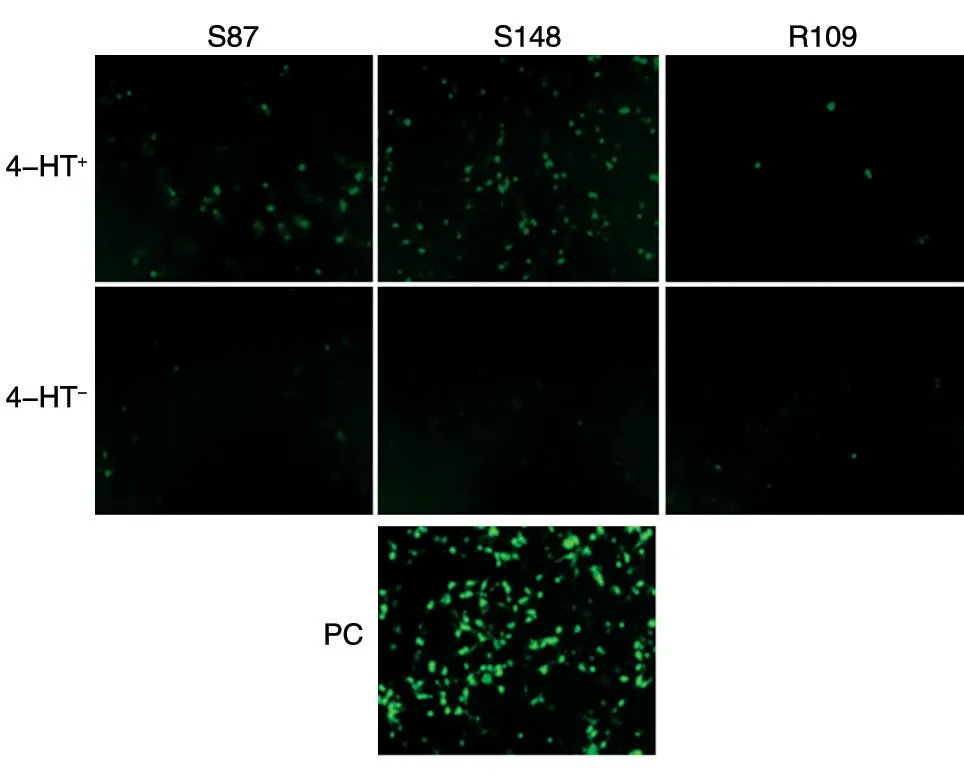
圖2 4-HT 調控eGFP 熒光檢測結果
3 討論
小分子依賴的內含肽調控系統(tǒng)是一類非常有吸引力的分子開關,原則上它們可以插入感興趣的任意蛋白質中,使得蛋白質功能依賴于小分子的激活。與轉錄和翻譯過程的條件控制相比,內含子剪接過程非常快速,因此,在通過激活轉錄或翻譯不能實現的時間尺度上添加小分子效應物后,可以激活積累的預裂蛋白。此外,與更常見的配體誘導轉錄行為相比,這種翻譯后激活取決于添加的小分子的濃度。因此,小分子激活的內含肽技術兼具小分子化學遺傳方法的目標特異性與經典遺傳學方法的通用普遍性特點。將進化的內含肽蛋白插入活細胞中的多種不相關蛋白質中,證實蛋白質功能的配體依賴性激活是普遍的、迅速的、劑量依賴性的和翻譯后修飾的。該過程不需要特定的細胞環(huán)境以及輔助因子的參與,甚至可以在體外進行。內含肽因其獨特的剪接功能,在蛋白質工程、基因診斷和治療、轉基因植物研究等領域有著廣闊的應用前景。

圖3 流式細胞術檢測熒光強度
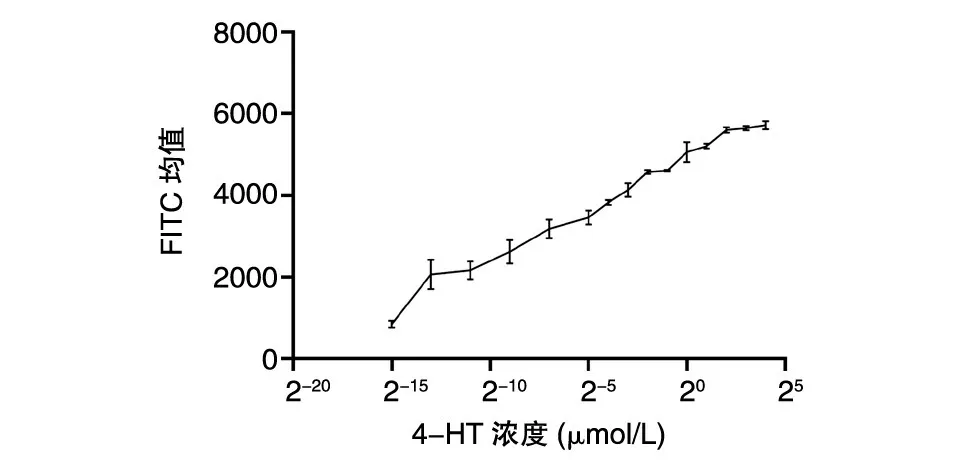
圖4 4-HT 濃度對剪接效率的影響
針對內含肽剪接的特點與eGFP 的結構與功能,利用同源重組的方法,我們將小分子調控的內含肽基因分別嵌入eGFP 蛋白中7 個可能的氨基酸位點,得到7 種pMD-eGFP-Intein 重組質粒。在所選取的7 個氨基酸位點中,包括文獻報道的R109 在內的3 個插入位點能夠實現小分子對eGFP 蛋白功能的調控,新發(fā)現的2 個插入位點S87 與S148 的剪接效率分別為9%和31%,遠高于文獻報道的剪接位點R109(剪接效率5%)。同時我們也發(fā)現,在剪接效率較高的位點,本底值相應也略高,這可能是由于在高效率剪接的位點上除了內含肽上受體結合域與4-HT 的碰撞幾率更高之外,內含肽自身的N 端與C 端結構域也比較靠近,自剪接的發(fā)生概率也會更高一點。
在S148 位點上,探究了4-HT 的最適濃度。發(fā)現4-HT 濃度在4 μmol/L 以下時,剪接效率與4-HT 濃度呈正相關,而當4-HT 濃度達到4 μmol/L 時,剪接效率不再增加,這說明在S148 位點上,4-HT 的最適濃度約為4 μmol/L,與文獻報道的R109 位點的最適工作濃度為1 μmol/L 存在差異,說明在所選取的不同調控位點上,4-HT 的最適濃度存在差異。由于差異不是很大,所以如果探究內含肽對于其他蛋白的調控,可以先選取1 μmol/L 尋找內含肽的替換位點,然后在調控效果較好的位點上摸索4-HT 工作濃度。
本實驗驗證了小分子依賴的內含肽剪接技術在eGFP 蛋白上應用的可行性,相較于文獻報道的位點得到了剪接效率更高的位點,并摸索了在相應位點上的小分子工作濃度,為后續(xù)這項剪接技術的應用打下了良好基礎。
猜你喜歡
瘋狂英語·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數理化(高中版.高考數學)(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經濟(2017年6期)2017-07-12 09:27:16
中國衛(wèi)生(2016年9期)2016-11-12 13:27:54
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國洗滌用品工業(yè)(2015年7期)2015-02-28 19:02:38
電子設計工程(2015年12期)2015-02-27 12:06:10
中國衛(wèi)生(2014年11期)2014-11-12 13:11:32
