布魯菌外膜蛋白Omp19突變體的構建及免疫原性研究
2019-12-25 08:43:40郭鳳羽殷瑛宰曉東李汭樺王美榮李耀輝胡凱李玉杰徐俊杰陳薇
生物技術通訊 2019年5期
郭鳳羽,殷瑛,宰曉東,李汭樺,王美榮,李耀輝,胡凱,李玉杰,徐俊杰,陳薇
軍事科學院軍事醫學研究院生物工程研究所,北京100071
布魯菌(Brucella)是一種胞內寄生的革蘭陰性細菌,是布魯菌病的病原體。布魯菌病是一種人畜共患的慢性傳染病[1],世界衛生組織(WHO)將該病歸為7 種被忽視的人畜共患病之一,是一種導致貧困持續存在的疾病[2]。人類主要通過食用被布魯菌污染的肉類及制品,或者接觸到被布魯菌感染的動物組織等方式罹患布魯菌病[3]。人布魯菌病的癥狀主要包括波浪熱、盜汗和體虛等,此外還常伴有全身乏力、頭痛、關節痛、陽痿等,孕婦感染布魯菌后容易引發自然流產[4]。由布魯菌病所引起的這些慢性感染往往難以根治,給患者的生理及心理健康帶來較大危害。布魯菌主要分布于發展中國家,全世界有170 多個國家和地區存在人、畜布魯菌病的流行,布魯菌病患者約600 萬人,每年新發病例約50 萬人次[5-6]。布魯菌病在我國主要集中在北方農牧業地區,現已擴展至全國31 個省市地區[7]。目前對布魯菌病的防治主要依靠疫苗,減毒活疫苗104M 是目前我國惟一獲批的人用布魯菌病疫苗,其安全性和有效性得到一定驗證,但仍存在接種方式復雜、免疫機制不明確、殘余毒力等問題[8]。因此,新型人用布魯菌疫苗的研制對于布魯菌病的預防和控制具有重要意義。
外膜蛋白19(outer membrane protein 19,Omp19)是存在于布魯菌表面的一種蛋白,在種型復雜的布魯菌中的同源性達90%以上[9]。Omp19蛋白全長有177 個氨基酸殘基,前端含有的信號肽使蛋白在胞內合成后被跨膜轉運,信號肽切割后蛋白被錨定于細菌外膜[10]。Omp19 對布魯菌黏附、侵襲、定殖及胞內存活等多個生理功能起關鍵作用[11]。此外,Omp19 還被報道能夠誘導Th1型免疫應答,是對抗布魯菌感染的重要靶標[12],也被認為有希望成為抗布魯菌病亞單位疫苗的重要候選組分[13]。Fiorentino 等通過敲除布魯菌疫苗株S19 的Omp19 基因發現Omp19 的缺失會使該疫苗失去保護能力[14]。另一項研究表明,給BALB/c小鼠口服重組表達的Omp19 蛋白,可以保護免疫小鼠對抗布魯菌的攻擊[15]。
本研究中,我們以前期構建的含有Omp19 核酸序列的質粒,對序列中編碼Cys 的堿基進行缺失或替換突變,構建一系列蛋白突變體,從中篩選到不形成2 條帶及多聚體的蛋白,比較突變體蛋白與Omp19 在理化性質及免疫保護性上的異同,為以Omp19 為組分的布魯菌重組蛋白疫苗的研究提供依據。
1 材料與方法
1.1 材料
6~8 周雌性BALB/c 小鼠購自北京維通利華實驗動物技術有限公司;大腸桿菌感受態DH5α、BL21(DE3)購自天根生化有限公司;布魯菌株104M、A19,質粒pET-32a 為實驗室保存;2×EasyTaqPCR supermix 購自北京全式金生物技術有限公司;限制性內切酶XhoⅠ、NdeⅠ,T4DNA 連接酶及T4多聚核苷酸激酶購自NEB 公司;核酸凝膠回收試劑盒及小量質粒提取試劑盒購自天根生化有限公司;HRP 標記的羊抗鼠IgG、IgG1、IgG2a購自Abcam 公司;SPXL 陽離子純化柱購自通用電氣公司;TMB 單組分顯色液及終止液、云克隆細胞因子檢測試劑盒購自北京蒂諾奧科技有限公司;引物由生工生物工程有限公司合成。
1.2 突變體質粒的構建
以前期構建的含有Omp19 核酸序列的質粒pET32a-Omp19 為模板,用引物M1u/M1d 擴增編碼Omp19 第1 位Cys 缺失突變體M1 的DNA 序列(94℃ 5 min;94℃ 30 s、60℃ 1 min、72℃ 1 min,30 個循環;72℃10 min)。用NdeⅠ和XhoⅠ酶切位點和T4DNA 連接酶將Omp19-M1 連接到pET32a 中,得到質粒pET32a-Omp19-M1。以測序正確的pET32a-Omp19-M1 為模板,分別用突變引物M2u/M2d、M3u/M3d、M4u/M4d 擴增Omp19-M1的86 位和106 位Cys 單點突變體和雙點突變體基因序列。以pET32a-Omp19 為模板,用突變引物M5u/M5d 擴增Omp19 的86 位和106 位雙點突變體(94℃5 min;94℃30 s、60℃30 s、72℃6 min,25 個循環;72℃ 7 min)。引物見表1,所有突變體結構示意圖見圖1。用T4多聚核苷酸激酶和T4DNA 連接酶將質粒進行磷酸化和連接,得到質粒pET32a-Omp19-M2~M5。
1.3 突變體蛋白的表達、鑒定和純化
將序列正確的突變體質粒轉化大腸桿菌BL21(DE)感受態,挑取工程菌單克隆接種到5 mL LB 培養基(含100 μg/mL 氨芐青霉素)中,37℃、220 r/min 過夜振蕩培養。次日以1∶100 的比例將培養物轉接至5 mL LB 液體培養基,37℃、220 r/min 振搖至菌液D600nm為0.6~0.8 時,加入終濃度為1 mmol/L 的IPTG 于28℃誘導表達5 h。離心收集菌體,進行SDS-PAGE 分析。
將過夜培養的重組菌以1∶100 的接種量接種至1 L LB 培養基(含100 μg/mL 氨芐青霉素),37℃振蕩培養至菌液D600nm為0.6~0.8,加入終濃度為1 mmol/L 的IPTG 于28℃誘導培養5 h,8000 r/min 離心5 min 收集菌體,菌體沉淀用40 mL 緩沖液(20 mmol/L NaAc,pH4.0)重懸,冰上超聲20 min,4℃、12 000 r/min 離心10 min,上清用0.45μm 濾器過濾作為純化樣品。用SPXL 陽離子柱,以梯度洗脫(洗脫液為20 mmol/L NaAc,1 mol/L NaCl,pH4.0)的方式純化目的蛋白。用15%的SDS-PAGE 鑒定蛋白,Lowrry 法檢測蛋白濃度。

圖1 突變體結構示意圖

表1 Omp19 突變體構建引物
1.4 突變體蛋白的理化性質分析
將純化得到的Omp19、M1、M4 委托北京百泰派克生物科技有限公司用液相色譜串聯質譜開展相對分子質量、N 端序列、二硫鍵的檢測。委托中國科學院過程工程研究所采用圓二色(circular dichroism,CD)光譜開展蛋白二級結構的檢測分析。
1.5 突變體蛋白的免疫評價
40 只6~8 周齡的BALB/c 雌性小鼠隨機分成5組,分別經皮下注射免疫Omp19、M1、M4 蛋白及104M 菌株,免疫佐劑為Al(OH)3。免疫104M 組為陽性對照組,PBS 組為陰性對照組(表2),除104M 免疫組只于0 d 免疫外,其余各組于0、14、28 d 免疫。
1.5.1 血清抗體滴度檢測 免疫35 d 時采血,分離血清,ELISA 法檢測抗Omp19 特異性抗體。Omp19 用包被緩沖液稀釋至2 μg/mL,按100 μL/孔加入96 孔酶標板,4℃過夜包被;次日用PBST(PBS,2%吐溫,pH7.2)洗板4 次,加入封閉液(PBS,2% BSA),37℃封閉1 h;洗板,待測小鼠血清梯度稀釋,37℃孵育1 h;洗板,按1∶10 000 的比例加入HRP 標記的羊抗鼠抗體IgG(檢測總抗體水平)或IgG1、IgG2a(檢測抗體亞類水平),37℃孵育45 min;洗板,加入TMB 單組分顯色液避光顯色6 min,加入終止液終止顯色。用Bio-Rad 酶標儀檢測D450nm和D630nm值,以D=D450nm-D630nm作為最終吸光度值,大于2.1 倍D(陰性)的最大稀釋度為該樣品的抗體滴度。
二是加強“職業化”建設,在技能培訓上強力度。近年來,馬鞍山市人社局充分發揮下崗再就業培訓基地的職能,聯合馬鞍山市婦聯開辦家政服務員職業資格認證培訓和專題講座,共開辦培訓班16期,培訓人員3871人次,2465人獲初、中級職業資格證書。馬鞍山市工人文化宮職教中心圍繞“家政服務工程”舉辦培訓班6期,300多名農民工、失業人員參加培訓。
1.5.2 細胞因子檢測 免疫后42 d,斷頸法隨機處死3 只小鼠,將無菌取出的脾臟置于篩網上,于RPMI1640 培養基中研磨;將研磨液轉移至15 mL離心管中,室溫500 r/min 離心5 min,棄上清;向沉淀中加入3 mL 紅細胞裂解液(ACK),室溫裂解5 min,再加入RPMI1640 培養基至終體積為10 mL,室溫500 r/min 離心5 min,棄上清;加入2 mL RPMI1640 培養基重懸細胞沉淀,得到脾臟單細胞懸液(1×106/mL)。在96 孔細胞培養板的各孔中加入100 μL 各組細胞懸液,再加入Omp19(5 μg/孔)用于刺激培養細胞,分別設置ConA(0.5 μg/孔)和RPMI1640 培養基作為陽性對照和陰性對照,37℃、5% CO2培養箱中孵育72 h 后離心收集細胞上清,用細胞因子檢測試劑盒檢測IFN-γ、TNF-α、IL-2、IL-6、IL-17A 的分泌水平。
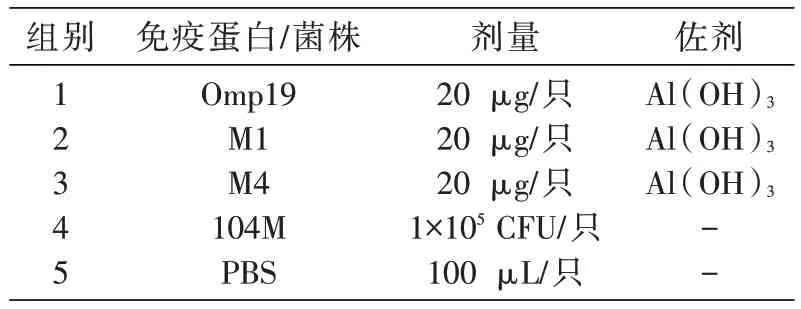
表2 蛋白免疫評價方式
1.6 攻毒評價
免疫后第35 d,各組隨機選取5 只小鼠,腹腔注射布魯菌株A19(1×106CFU/只)進行攻毒評價,104M 疫苗株免疫組(1×105CFU/只)作為陽性對照,PBS 免疫組作為陰性對照。14 d 后將小鼠處死,無菌條件下摘取小鼠脾臟,將臟器在PBS 溶液中研磨制成勻漿,取組織勻漿100 μL 做適度濃度梯度稀釋后涂布無抗性的TSA 平板,37℃孵箱培養3 d 后進行臟器菌載量計數。結果以菌載量對數的平均值的形式(mean LgCFU)表示,同時將免疫組小鼠脾臟中布魯菌菌載量相對于對照組小鼠菌載量下降的數值定義為保護指數,即對照組與實驗組的平均LgCFU 的差值。
1.7 統計學方法
顯著性分析采用單因素方差分析或t檢驗,以P<0.05 為有統計學差異。數據統計分析及制圖均由GraphPad Prism 7.0 完成。
2 結果
2.1 突變體表達載體的構建
以pET32a-Omp19 質粒為模板擴增M1 的基因片段,通過2 條攜帶缺失突變的引物,分別將編碼Omp19 第1 位Cys 的堿基缺失,得到長度為468 bp 的M1 的序列片段,與理論值相符(圖2A)。同理,分別以pET32a-Omp19 和pET32a-M1質粒為模板,利用反向PCR 分別擴增得到大小約6000 bp,含有86、106 位單位點及雙位點突變的pET32a-M2~M5 質粒的線性片段(圖2B)。經測序鑒定,得到的核酸序列與預期一致。
2.2 突變體蛋白的表達與純化
對序列正確的突變體質粒進行誘導表達,陽離子柱SPXL 純化蛋白,用SDS-PAGE 對Omp19、突變體蛋白M1~M5 進行還原及非還原電泳(圖3)。突變體M1 相對理論分子質量為15 810,M2和M3 為15 780,M4 為15 820,M5 為15 870,電泳結果顯示條帶大小約為23 000,與預期稍有差異。此外,電泳結果顯示,Omp19 在非還原電泳條件下存在多聚體和多余條帶,第2 位Cys 的缺失(M1)消除了多余條帶的影響,第86 和106 位Cys 的替換(M4)不僅去除了多余條帶還能去除多聚體。

圖2 突變體片段擴增及質粒構建
2.3 突變體蛋白理化性質分析
為了研究Cys 的缺失或替換對Omp19 原有理化性質和免疫原性的影響,利用液相色譜串聯質譜分別比較了Omp19、M1 及M4 在相對分子質量、N 端序列組成及二硫鍵組成上的異同。利用圓二色光譜分別比較了3 種蛋白二級結構的異同。
2.3.1 相對分子質量測定 采用液相色譜串聯質譜(LC-MS/MS)檢測Omp19、M1、M4 蛋白的相對分子質量(圖4)。測得Omp19 的相對分子質量為31 550(理論值15 909),約為理論值的2 倍;M1的相對分子質量為15 804(理論值15 805),M4的相對分子質量為15 757(理論值15 757),均與其理論值相差不大。
2.3.2 N 端序列測定 蛋白質的N 端與蛋白質的功能和穩定性息息相關。本研究檢測了M1 及M4蛋白的N 端6 個氨基酸殘基,M1 N 端序列為NH2-Met-Gln-Ser-Ser-Arg-Leu,M4 N 端序列為NH2-Met-Gln-Ser-Ser-Arg-Leu,與預期序列一致。

圖3 突變體蛋白表達及純化

圖4 質譜分析測定Omp19、M1 及M4 的相對分子質量
2.3.3 二硫鍵分析 采用蛋白酶解及LC-MS/MS方法檢測Omp19、M1、M4 蛋白分子中二硫鍵的數量及位置,在Omp19 第1、86、106 位均檢測到二硫鍵存在;M1 第86 及106 位檢測到二硫鍵,M4 未檢測到二硫鍵的存在,與預期結果一致。
2.3.4 二級結構分析 遠紫外圓二色光譜可以提供蛋白質的二級結構信息。Omp19、M1 及M4 的遠紫外圓二色光譜如圖5,3 種蛋白的光譜圖幾乎完全重合。通過儀器自帶軟件,擬合了3 種蛋白中4 種二級結構的含量,如表3 所示。可以看到,3 種蛋白的α螺旋、β折疊和自由卷曲3 種二級結構的含量基本一致,只在β轉角有稍許差異,這個結果提示2 種突變體蛋白各二級結構的含量與Omp19 稍有不同但差別不大。
2.4 突變體蛋白的免疫評價
2.4.1 抗體水平 小鼠免疫后第35 d 取血,ELISA檢測針對Omp19 的特異性總抗體水平及抗體亞類水平(圖6)。結果顯示Omp19、M1、M4 免疫組激發的針對Omp19 的特異性IgG 抗體滴度分別為3.53×104、2.19×104、2.76×104,3 組間無顯著性差異。此外,如圖6B 所示,Omp19、M1、M4 免疫組激發的抗體亞類IgG1/IgG2a 的比值分別為1.48、1.45、1.40,IgG1/IgG2a 比值均大于1,3 組間無顯著性差異。

圖5 Omp19、M1 及M4 的遠紫外圓二色譜

表3 Omp19、M1 及M4 的二級結構分析
2.4.2 細胞因子水平 小鼠初次免疫后42 d,用ELISA 檢測試劑盒測定Omp19 體外刺激后各組小鼠脾細胞培養上清中細胞因子的含量(圖7)。結果表明,Omp19 抗原刺激后,Omp19、M1 及M4 免疫組小鼠脾細胞培養上清中檢測到的IFN-γ、IL-2、IL-6 分泌水平沒有顯著性差異,但M4 免疫組小鼠脾細胞培養上清中檢測到的TNF-α和IL-17A 水平顯著低于Omp19 和M1 組。
2.5 免疫保護效果
為了評價3 種蛋白的保護效果,小鼠初次免疫后第35 d,用布魯菌A19(1×106CFU/只)對免疫小鼠進行腹腔攻擊,利用保護指數評價Omp19、M1 及M4 的保護效力。結果顯示,Omp19及突變體免疫組小鼠攻毒后小鼠脾臟中的菌載量相對于PBS 組均顯著降低(P<0.05),3 組之間沒有顯著差異,保護指數分別為1.42、1.39、1.38,均高于PBS 組。同時也觀察到3 組的保護指數與104M 疫苗株存在一定差距,結果見表4。
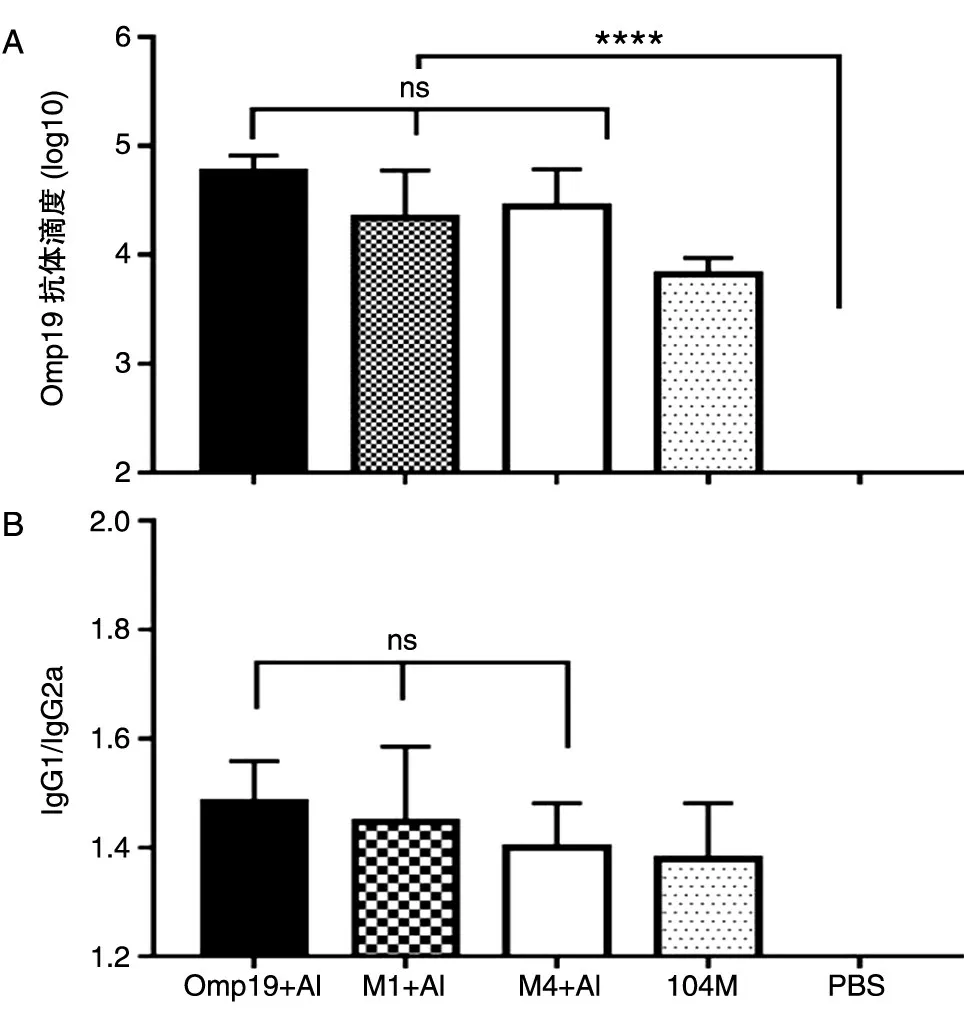
圖6 Omp19 抗原特異的抗體水平
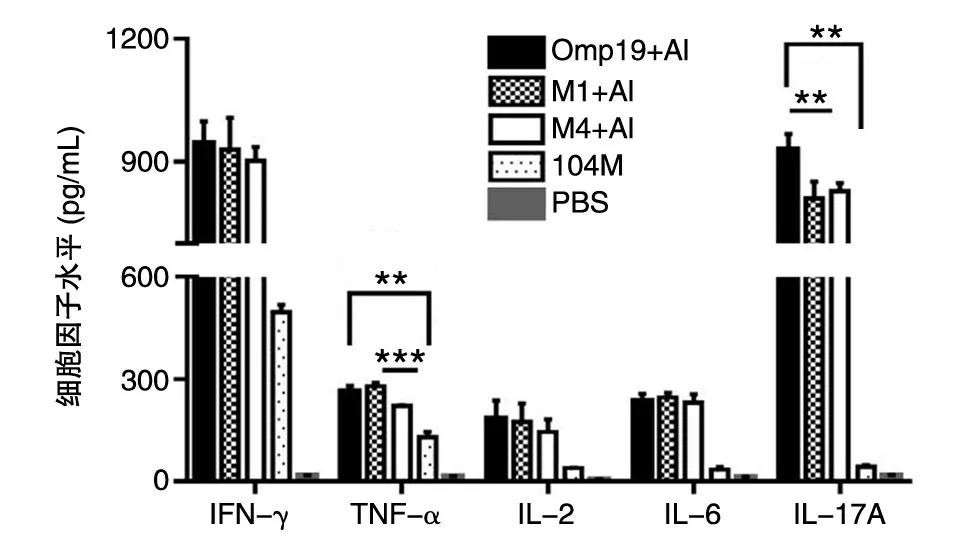
圖7 Omp19 特異的細胞因子水平

表4 小鼠攻毒實驗保護性效果統計表
3 討論
我國的布魯菌病疫情自1993 年出現反彈,隨后發病率迅速增長[16]。最新的全國法定傳染病疫情概況數據顯示,2018 年我國上報布魯菌病新發病例達到3.8 萬人次,發病率為2.73/10 萬,仍然處于較高位置[17]。疫苗是預防和控制布魯菌病的主要手段。目前,獸用布魯菌病疫苗已獲得廣泛使用,在控制布魯菌病疫情方面發揮著重要作用。人用布魯菌病疫苗研制較為困難,目前國外尚無疫苗上市,我國惟一獲批的人用布魯菌病疫苗為104M 株。104M 是一種皮上劃痕接種的減毒活疫苗,存在免疫機制不明確、有殘余毒力等問題[8]。隨著生物技術的不斷發展,以重組蛋白疫苗與核酸疫苗為主的新型疫苗近年來成為布魯菌病疫苗研究的熱點[18],國內外也對包括Omp19 在內的多個布魯菌抗原進行了初步研究,其中Omp19 被認為對布魯菌具有一定的保護作用[9],但是相關的研究都不系統。
實驗室前期發現在制備Omp19 蛋白時,目的蛋白附近存在另一條與之大小類似,且能與多抗結合的條帶,同時該蛋白還易形成多聚體。經過對Omp19 蛋白氨基酸序列的分析,發現Omp19 蛋白序列分別在第1、86、106 位含有Cys,推測這可能是造成上述現象出現的原因。為了探究Cys 在序列中的作用,我們構建了一系列蛋白突變體并比較了相應的理化性質和免疫原性。SDS-PAGE鑒定結果顯示Omp19 及突變體蛋白的相對分子質量為25 000,與理論值稍有差異;得到的突變體蛋白M1 去除了目的蛋白附近多余條帶并減少了多聚體或二聚體的形成,而突變體M4 不僅可以去除多余條帶,還能同時去除多聚體或二聚體。質譜檢測結果顯示Omp19 的相對分子質量為31 550,約為理論值的2 倍;突變體M1、M4 的相對分子質量分別為15 804、15 757,與理論值基本相符。提示重組表達的Omp19 蛋白主要以二聚體形式存在,缺失第1 位Cys 或同時對86 位Cys 進行Ala 替換,對106 位Cys 進行Ser 替換可以減少或避免二聚體的形成。二硫鍵檢測結果表明,Omp19 蛋白中可以檢測到更多數量的二硫鍵位點,而M4 蛋白中檢測不到二硫鍵位點的存在,結合SDS-PAGE 和的相對分子質量檢測結果,推測Omp19 蛋白中多聚體或二聚體的形成可能是由Cys 介導的二硫鍵引起。此外,圓二色譜結果顯示,Cys 的缺失或替換不會對Omp19 的二級結構造成太大影響。免疫學檢測結果表明,相較于Omp19,M1 和M4 中Cys 的缺失或替換不會顯著影響Omp19 特異的總抗體水平和所激發的免疫反應類型,主要以Th2 型反應為主。Omp19 及突變體蛋白M1、M4 免疫組小鼠脾細胞經抗原體外刺激后分泌的Omp19 特異的IFN-γ、IL-2、IL-6 水平沒有顯著性差異,但M4 免疫組小鼠脾細胞培養上清中分泌的TNF-α 和IL-17A 水平顯著低于Omp19 和M1 組。這個結果提示86 和106 位Cys的替換會影響Omp19 特異的部分細胞因子的產生。攻毒實驗結果表明,Omp19 及其突變體對布魯菌A19 株的攻擊均具有一定的保護效果,突變體蛋白激發的免疫保護效力與Omp19 沒有顯著性差異。
綜上所述,Cys 會影響Omp19 重組蛋白的制備和質控,其缺失或替換對Omp19 原有的理化性質和免疫原性影響不大。本工作為Omp19 作為布魯菌亞單位疫苗有效組分的研究提供了依據。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
