豬FTO基因多態性與肉質性狀的關聯分析
2019-12-24 07:06:14楊又兵卞軍平李世豪
浙江農業學報 2019年12期
楊又兵,卞軍平,吳 燕,婁 然,李世豪
(1.河南科技大學 動物科技學院,河南 洛陽 471023;2.洛陽市動物遺傳育種重點實驗室,河南 洛陽 471023)
脂肪量和肥胖相關基因(fat mass and obesity associated gene,FTO)廣泛地分布在動物的各個組織之間,在畜禽的生長發育、能量代謝平衡和脂肪代謝等方面發揮重要的作用[1-2]。首先,FTO基因是在小鼠上克隆出來的[3],隨后,Stratakis等[4]在研究人類染色體時發現FTO基因與肥胖相關,Frayling等[5]發現FTO基因的過度表達會提高動物采食量,其他研究者[6-8]發現FTO基因在控制肥胖方面發揮關鍵作用。在一些生物醫學研究領域,豬是研究肥胖相關基因的最好模型,大量研究[9-11]發現,豬的脂肪性狀影響著豬的肉質性狀。后來,隨著FTO基因的深入研究,該基因也被越來越多的學者應用在豬的脂肪沉積和肉品質研究中。例如:Tao等[12]在研究不同品種的豬FTO基因mRNA表達量水平發現品種對豬FTO基因mRNA表達量具有顯著影響;Chung[13]研究了FTO基因的SNP位點與韓國牛的肉質性狀的關聯性,研究表明位于外顯子3上的g.125550A>T位點與肉質性狀具有顯著相關;Fan等[14]在研究巴克夏豬×大白豬資源群體時表明FTO基因第3外顯子c.594C>G位點各基因型與肌內脂肪總量存在極顯著相關性。

1 材料與方法
1.1 試驗動物
從河南省洛陽市豫西黑豬保種場抽取100~120 kg的27頭豫西黑豬個體(編號HB),從河南洛陽正大食品有限公司抽取100~120 kg的27頭長白豬個體(編號HL),收集血樣,-20 ℃保存備用。屠宰測量27頭豫西黑豬個體的背膘厚度(BFT)、45~60 min內的pH1、24 h熟化后的pH24、滴水損失,評定其肉色和大理石紋。
1.2 主要試劑和儀器
全血基因組DNA提取試劑盒、2×TaqPCR Master Mix、BstuⅠ限制性內切酶、TaiⅠ限制性內切酶和DL 2 000 bp Maker均來自于鼎國昌盛生物技術有限責任公司(中國北京)。
pHS-3C酸度計:鄭州寶晶電子科技有限公司(中國河南)。背膘檢測儀:北京天翔飛域科技有限公司(中國北京)。高速臺式離心機:上海安亭科學儀器(中國上海)。電泳儀:鼎國昌盛生物技術有限責任公司(中國北京)。梯度PCR擴增儀:BIO-RAD公司(中國上海)。凝膠成像分析系統:BIO-RAD公司(中國上海)。
1.3 引物設計
根據NCBI(https://www.ncbi.nlm.nih.gov/)上已經公布的FTO基因第3外顯子(GenBank:FJ668708)和第4內含子的序列(GenBank:AM931150),利用Primer Premier 5.0軟件(http://www.premierbiosoft.com/),設計FTO基因第3外顯子和第4內含子的引物,由上海生工生物工程股份有限公司合成。第3外顯子(c.594C>G)目的片段為240 bp,退火溫度為56.8 ℃,引物序列為:Bstu I-F:5′-TGCAGATTGAGACCATCCAG-3′;Bstu I-R:5′-TCTTCCCCATGCCAAAGTAG-3′。第4內含子(g.276G>T)目的片段為397 bp,退火溫度為58.4 ℃,引物序列為:TaiI-F:5′-ACAGGCCCTGAAGAGGAAAG-3′;TaiI-R:5′-AGTAACCTGGAGTTCCTGTGG-3′。
1.4 PCR擴增
利用全血基因組DNA提取試劑盒提取DNA,提取步驟參照其說明書,以BstuⅠ-F和BstuⅠ-R為上下游引物進行第3外顯子(c.594C>G)序列的PCR擴增,以TaiⅠ-F和TaiⅠ-R為上下游引物進行第4內含子(g.276G>T)序列的PCR擴增。PCR反應體系(20 μL)為: 2×TaqPCR Master Mix 10 μL,上下游引物各0.7 μL(10 μmol·L-1,DNA模板1 μL,加ddH2O至終體積20 μL。PCR擴增條件為:94 ℃預變性4 min;94 ℃變性30 s,退火溫度退火30 s,72 ℃延伸30 s,35個循環;72 ℃延伸10 min,4 ℃保存。擴增產物經2%的瓊脂糖凝膠電泳,用凝膠成像儀觀察擴增條帶。
1.5 PCR-RFLP酶切反應
用限制性內切酶對FTO的兩個SNP位點進行酶切分型,酶切體系(15 μL):PCR產物10 μL,10×Buffer 2 μL,限制性內切酶(10 U·μL-1)0.5 μL,ddH2O補足至15 μL。反應條件為:BstuⅠ的酶切體系于37 ℃水浴消化3.5 h,用50%的聚丙烯凝膠電泳檢測基因型,TaiⅠ的酶切體系于65 ℃水浴消化3.5 h,用2%的瓊脂糖凝膠電泳,凝膠成像儀觀察記錄,判斷基因型。
1.6 肉質性狀測定
參照1987年制訂的肉質評定標準修改方案《豬肉質評定方法》對27個豫西黑豬樣本的肉質性狀進行評定。背膘厚:用背膘檢測儀測定肩部膘厚、胸腰部膘厚、臀部膘厚,取三處平均值;pH值:用pHS-3C酸度計測定45~60 min的pH1,24 h熟化后的pH24;滴水損失:宰后2 h內,取腰椎處背最長肌修整樣稱取質量m1,在4 ℃下懸掛24 h,用潔凈濾紙吸取表層液體后稱取質量m2,滴水損失=[(m1-m2)/m1]×100%;用美式比色卡進行肉色和大理石紋評分。
1.7 數據統計分析
用POPGENE 32計算等位基因頻率、有效等位基因(Ne)、基因多樣性(Shannon’s information index,I)、期望雜合度(expected heterozygosity,He)和Nei氏多樣性指數,用PIC-CALC計算多態信息含量(polymorphic information content,PIC),用SPSS 20.0的χ2檢驗驗證c.594C>G位點和g.276G>T位點的Hardy-Weinberg平衡,對各基因型之間的肉質性狀數據進行顯著性分析,數據結果以X±SD表示。
2 結果與分析
2.1 FTO基因的PCR擴增結果
FTO基因的c.594C>G位點和g.276G>T位點經PCR擴增后,擴增產物用2%的瓊脂糖凝膠電泳檢測,檢測結果見圖1和圖2,圖1和圖2中的擴增條帶清晰明亮,擴增條帶單一,無非特異性條帶,可用作下一步的酶切反應。
2.2 FTO基因的PCR-RFLP酶切結果
用BstuⅠ限制性內切酶對c.594C>G位點進行酶切,酶切產物用50%的聚丙烯凝膠進行檢測,結果見圖3。用TaiⅠ限制性內切酶對g.276G>T位點進行酶切,酶切產物用2%的瓊脂糖凝膠電泳檢測,結果見圖4。圖3的BstuⅠ限制性內切酶將c.594C>G位點的擴增產物切割成3個片段(240 bp/156 bp/84 bp),出現兩個等位基因C(240 bp)和G(156 bp/84 bp),3種基因型即CC基因型(只240 bp一種條帶)、CG基因型(同時出現240 bp、156 bp和84 bp三種條帶)和GG基因型(出現156 bp和84 bp兩種條帶)。圖4的TaiⅠ限制性內切酶將g.276G>T位點的擴增產物切割成3個片段(397 bp/275 bp/122 bp),出現兩個等位基因G(275 bp/122 bp)和等位基因T(397 bp),3種基因型即GG基因型(出現275 bp和122 bp兩種條帶)、GT基因型(同時出現397 bp、275 bp和122 bp三種條帶)和TT基因型(只397 bp一種條帶)。

M,DL2000 bp DNA marker;1~4,豫西黑豬;5~6,長白豬。M, DL2000 bp DNA marker; 1-4, Henan western black pigs; 5-6, Landrace pigs.圖1 c.594C>G位點PCR產物擴增結果Fig.1 PCR amplification results of SNPs (c.594C>G)
M,100 bp DNA marker;1~2,豫西黑豬;3~4,長白豬。M, 100 bp DNA marker; 1-2, Henan western black pigs; 3-4, Landrace pigs.圖2 g.276G>T位點PCR產物擴增結果Fig.2 PCR amplification results of SNPs (g.276G>T)
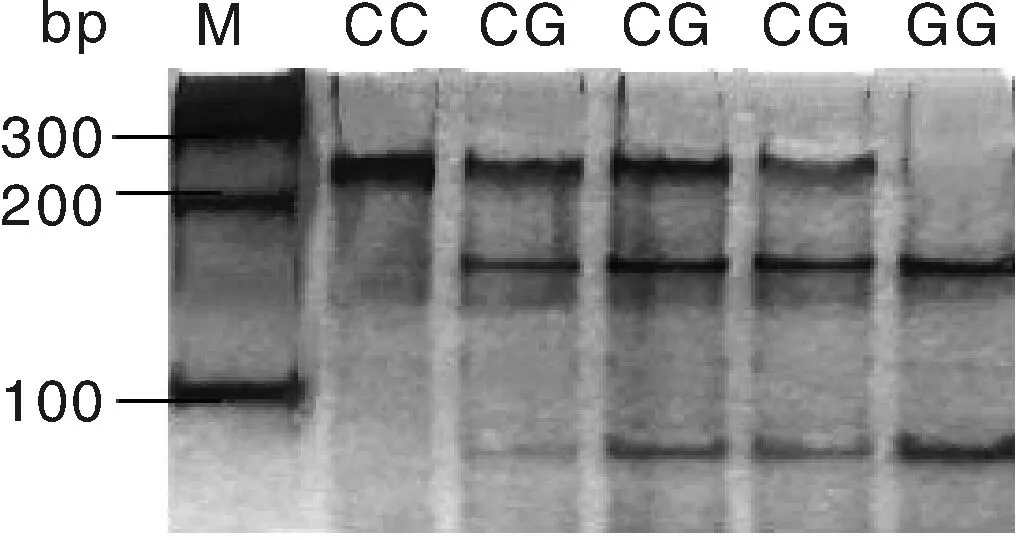
M, 100 bp DNA marker.圖3 c.594C>G位點酶切結果Fig.3 Enzyme digestion results of c.594C>G

M, 100 bp DNA marker.圖4 g.276G>T位點酶切結果Fig.4 Enzyme digestion results of g.276G>T
2.3 FTO基因的基因型頻率與基因頻率
表1是FTO基因c.594C>G位點和g.276G>T位點的基因型頻率與基因頻率。在c.594C>G位點,豫西黑豬和長白豬都有CC基因型、CG基因型和GG基因型3種基因型,優勢等位基因為C等位基因,豫西黑豬的CG基因型頻率(0.681 8)高于長白豬(0.533 3),CC基因型(0.181 8)和GG基因型(0.136 4)的頻率低于長白豬(0.266 7、0.200 0)。三種基因型χ2檢驗的Hardy-Weinberg平衡檢驗P=0.320,頻率符合Hardy-Weinberg平衡(P>0.05)。在g.276G>T位點,豫西黑豬和長白豬都檢測到TT基因型、GT基因型和GG基因型3種基因型,優勢等位基因為T等位基因,3種基因型χ2檢驗的Hardy-Weinberg平衡檢驗P=0.125,說明g.276G>T位點基因型頻率符合Hardy-Weinberg平衡(P>0.05)。
表1FTO基因的基因型頻率和基因頻率
Table1The genotype frequency and allele frequency ofFTOgene

位點LocusHBHLχ2Pc.594C>G基因型頻率Genotype frequencyCC0.181 80.26672.2800.320CG0.68180.5333GG0.13640.2000基因頻率Allele frequencyC0.52270.5333G0.47730.4667g.276G>T基因型頻率Genotype frequencyGG0.11110.14824.1560.125GT0.62960.4074TT0.25930.4444基因頻率Allele frequencyG0.42590.3519T0.57410.6481
2.4 FTO基因的遺傳多態性分析
本研究對豫西黑豬和長白豬群體FTO基因的c.594C>G位點和g.276G>T位點進行了遺傳多態性分析,分析結果見表2。一般通過多態信息含量(PIC)、雜合度(He)和有效等位基因數(Ne)來衡量一個群體的遺傳多態性[19],參照Nagy等[20]的研究,PIC>0.5為高度多態;PIC在0.25~0.50時為中度多態;PIC<0.25時為低度多態。在c.594C>G位點上,豫西黑豬群體和長白豬群體的PIC值分別為0.374 5、0.373 9,兩個群體都屬于中度多態。在g.276G>T位點上,豫西黑豬群體和長白豬群體的PIC值分別為0.369 4、0.352 1,兩個群體都具有多態性,都屬于中度多態。在c.594C>G位點上,豫西黑豬的期望雜合度(He)和基因多樣性指數(I)分別為0.510 6,0.499 0,較長白豬的期望雜合度(0.514 9)和基因多樣性指數(0.497 8)無太大差別。在g.276G>T位點上,豫西黑豬的期望雜合度He(0.498 3)和基因多樣性指數(0.682 1)都略高于長白豬在該位點上的期望雜合度(0.464 7)和基因多樣性(0.648 6)。
表2FTO基因的遺傳多態性
Table2The genetic polymorphic ofFTOgene

位點LocusNeIHeNeiPICc.594C>GHB1.99590.49900.51060.49900.3745HL1.99120.49780.51490.49780.3739g.276G>THB1.95700.68210.49830.48900.3694HL1.83860.64860.46470.45610.3521
2.5 FTO的遺傳多態性與豫西黑豬肉質性狀的關聯分析
本研究將FTO基因c.594C>G位點和g.276G>T位點的多態性與豫西黑豬的肉質性狀進行了關聯分析,分析結果見表3。在c.594C>G位點,等位基因G能顯著提高肉色,CG基因型和GG基因型個體在肉色上顯著高于CC基因型個體(P<0.05),在其他性狀上3種基因型并沒有表現出差異顯著性,但從表3可知,等位基因G能增加背膘厚度和提高大理石紋的評分,在背膘厚度和大理石紋上,GG基因型的個體>CG基因型>CC基因型。在g.276G>T位點,TT基因型、GT基因型和GG基因型這3種基因型在背膘厚度、pH1、pH24、滴水損失、肉色和大理石紋上沒有顯著性差異。但等位基因G能增加背膘厚度,提高pH1和pH24的值,降低滴水損失。在背膘厚度、pH1和pH24上,GG基因型的個體>GT基因型>TT基因型,滴水損失上,TT基因型的個體>GT基因型>GG基因型。
3 討論
影響肉品質的因素主要有飼養管理、營養水平、屠宰條件和遺傳因素,遺傳因素中的肉質性狀相關基因在影響肉品質方面起著至關重要的作用[21]。FTO基因是影響豬肉品質的一個遺傳因素,周波[22]通過原核表達探討豬FTO基因的組織表達規律表明了FTO基因在一定程度上能顯著促進豬脂肪細胞的增殖和分化,影響豬肌內脂肪的沉積。
表3FTO基因不同基因型對豫西黑豬肉質性狀的影響
Table3Effect of different genotypes in theFTOgene on meat quality traits

位點Locus基因型Genotype背膘厚度Backfatthickness/mmpH1pH24滴水損失Driploss%肉色Colorscore大理石紋Musclemarblingc.594C>GCC37.67±15.056.26±0.565.84±0.054.00±1.812.75±0.52 b3.00±0.71CG38.42±6.966.69±0.405.93±0.212.36±1.853.54±0.63 a3.09±0.67GG43.52±12.946.46±0.345.90±0.163.93±2.403.63±0.48 a3.25±0.65g.276G>TTT38.34±12.746.55±0.425.85±0.143.43±2.013.45±0.603.20±0.63GT39.00±7.916.59±0.475.94±0.212.68±2.003.36±0.683.00±0.63GG39.50±5.426.92±0.085.97±0.111.12±0.114.00±0.713.75±1.06
同列數據后沒有相同字母的表示差異顯著(P<0.05)。
The data in the same columns with different letters showed significant difference (P<0.05).
現階段,改善畜禽的肉質性狀是動物育種的一個重要目標[23-24],而脂肪量與肥胖相關基因是影響豬肉肉質性狀的一個重要基因,它在動物的各個組織中都能夠廣泛表達。付言峰等[25]研究了FTO基因對豬脂肪沉積的影響,發現在背肌中FTO基因mRNA的表達量極顯著高于其他任何組織,而且FTO基因的表達量越高,脂肪含量就越高。全建平等[26]利用高分辨率熔解曲線(HRM)技術檢測了374頭杜洛克豬FTO基因的基因型,表明FTO基因的多態性與杜洛克豬的背膘厚、瘦肉率、飼料轉化率、體高、體長等性狀顯著相關。本研究中,在c.594C>G位點的CG基因型和GG基因型個體在肉色上顯著高于CC基因型個體(P<0.05),等位基因G能增加背膘厚度和提高大理石紋的評分,促進豬的脂肪沉積。在g.276G>T位點,等位基因G能增加背膘厚度,提高pH1和pH24的值,降低滴水損失。
豫西黑豬作為河南省一個正在大力推廣的地方類群,有著它自身獨特的優勢,但是,由于其生長速度緩慢,養殖戶們為了提高短期內的經濟效益,對當地的豫西黑豬進行了長期的雜交選育,給豫西黑豬導入了過多的外來血源。昔奮攻等[27]對6個品種的豬的肥胖基因位點進行了RFLP分析,他表示“商品豬大多為瘦肉型豬,中國大部分的地方豬種都為脂肪型豬,具有較強的脂肪蓄積能力”。其他一些研究[28-29]表明,FTO基因對杜洛克群體的背膘性狀具有顯著影響,但是FTO多態性對豬的脂肪沉積性狀的影響在不同地區不同品種的豬之間存在著很大的差異性,這也與Fu等[30]的試驗結果相同,即FTO基因多態性與蘇鐘豬的脂肪沉積性狀相關,但是在某些品種的豬上,這種相關性存在很大的差異。例如,FTO基因對杜洛克的脂肪沉積有極大的顯著性[31],但是,FTO基因在槐豬背膘厚度這一性狀上并不存在顯著相關性[32]。在本研究中,c.594C>G位點的G等位基因能夠增加豬肉的背膘厚度,改善肉色,提高大理石紋評分,有利于脂肪的沉積,g.276G>T位點的G等位基因能增加豬肉的背膘厚度,有效降低豬肉的滴水損失,大大增加了豬肉的系水能力。但是,豫西黑豬和長白豬在c.594C>G位點的優勢等位基因都為C等位基因,在g.276G>T位點的優勢等位基因都為T等位基因,這說明了豫西黑豬混入了長白豬的部分血源,它自身的脂肪蓄積能力有所降低。本研究通過對豫西黑豬FTO基因多態性的分析,說明FTO多態性確實與豫西黑豬的肉質性狀具有相關性,本研究能夠為豫西黑豬的保種選育提供理論依據。
4 結論
在本研究中,FTO基因的c.594C>G位點和g.276G>T位點都具有中度多態性,豫西黑豬在FTO基因c.594C>G位點和g.276G>T位點存在遺傳多態性。在c.594C>G位點,CG基因型和GG基因型個體在肉色上顯著高于CC基因型個體(P<0.05),G等位基因能夠增加豬肉的背膘厚度,改善肉色,提高大理石紋評分,有利于脂肪的沉積。在g.276G>T位點,等位基因G能增加背膘厚度,提高pH1和pH24的值,降低滴水損失,增加了豬肉的系水能力。研究說明,FTO基因的c.594C>G位點和g.276G>T位點可以作為豫西黑豬肉質性狀選育的潛在分子遺傳標記,可為豫西黑豬群體后代肉質性狀的選擇提供參考依據。
