Orai2對成骨細胞增殖、凋亡及分化的作用研究
2019-12-23 05:23:38郭云山郝定均王曉東胡慧敏姜擴黃研生楊小衛
實用骨科雜志 2019年12期
關鍵詞:檢測
郭云山,郝定均,王曉東,胡慧敏,姜擴,黃研生,楊小衛
(西安交通大學附屬紅會醫院脊柱外科,陜西 西安 710054)
骨質疏松癥是一類常見的骨骼系統退行性疾病,嚴重影響老年人的健康。成骨細胞增殖減少、凋亡增加以及分化異常是骨質疏松癥發生發展的重要原因[1-5]。因此,深入研究成骨細胞增殖、凋亡及分化的調控機制,發現并鑒定在骨形成過程中具有成骨作用的關鍵調控分子,將為臨床骨質疏松癥的生物治療提供新的方法。Orai是一類細胞膜上的鈣通道蛋白,包括Orai1蛋白和Orai2蛋白。研究發現缺失鈣通道蛋白Orai1基因會導致小鼠骨密度降低等骨質疏松癥的表型,提示鈣通道蛋白Orai1參與骨質疏松癥的發生過程[6-9]。然而,目前尚無鈣通道蛋白Orai2參與骨質疏松癥發生發展的相關報道。本研究從Orai2調控成骨細胞的增殖、凋亡以及分化入手,研究Orai2在骨質疏松癥發生中的作用,以期更為全面地認識骨質疏松癥發生發展的病理過程。
1 資料與方法
1.1 材料 Orai2干涉片段購自上海生工公司,Orai2、Runx2和Osterix的引物全部從上海生工公司購買,RPMI1640細胞培養液從美國GIBCO BRL公司購買,四氮唑藍(MTT)從中國碧云天試劑公司購買,Real-time PCR試劑盒從美國invitrogen公司購買,Annexin V/PI試劑盒從美國Abcam公司購買,流式細胞儀和Real-time PCR儀從美國BIO-RAD公司購買。
1.2 方法
1.2.1 Real-Time PCR 小鼠成骨細胞系MC3T3-E1分別轉染靶向Orai2的shRNA和空白對照shRNA后,將MC3T3-E1成骨細胞分為對照組和Orai2 shRNA組。對應時間節點將對照組和Orai2 shRNA組收集到2 mL離心管。然后用Trizol法提取細胞總RNA,再將細胞總RNA反轉錄成cDNA,具體步驟如下:在離心管中加入2 μL的模板RNA溶液、1 μL的Oligo dT primer溶液、2 μL dNTP mixture溶液、1 μL Ace反轉錄酶溶液、1 μL RNase inhibitor溶液、4 μL RT buffer溶液,10 μL RNase-free水。反應條件:第一步37 ℃ 30 min,第二步84 ℃ 30 s。反應產物4 ℃保存。然后進行實時定量PCR反應。反應條件:12.5 μL RealMasterMix溶液、2 μL模板cDNA溶液、1 μL正向引物溶液、1 μL反向引物溶液、8.5 μL RNase-free水。反應條件為:第一步94 ℃ 5 min;第二步94 ℃ 60 s、57 ℃ 30 s、72 ℃ 30 s,共30個循環;第三步72 ℃ 5 min;最后4 ℃結束反應。
1.2.2 免疫印記實驗 對照組和Orai2 shRNA組待檢測細胞裂解后進行總蛋白定量,然后取等量的細胞裂解樣品進行SDS電泳90 min,取下SDS凝膠轉膜45 min,然后脫脂奶粉封閉約30 min。洗膜4次后在膜上滴加Orai2的抗體孵育90 min,再次洗膜4次。然后加入相應辣根過氧化物酶(和horseradish peroxidase,HPR)標記二抗。在聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF)上滴加適量化學發光液,然后將PVDF膜置于膠片盒中,曝光適當時間后,進行拍照與定量。
1.2.3 細胞周期檢測 對照組和Orai2 shRNA組細胞培養于無血清培養基24 h后,在細胞培養中10%胎牛血清(fetal bovine serum,FBS),然后繼續培養細胞。24 h后將細胞收集至離心管,用磷酸鹽平衡生理鹽水(public broadcasting service,PBS)洗滌3次后棄上清離心液。然后用1 mL無水乙醇重懸細胞。將細胞置于4 ℃冰箱中過夜。PBS洗滌細胞3次。然后在細胞中加入500 μL的PI染料重懸細胞。將細胞置于4 ℃冰箱中染色40~60min。最后,用流式細胞儀檢測細胞周期,并統計學分析實驗數據。
1.2.4 Annexin V/PI雙染檢測細胞凋亡 對照組和Orai2 shRNA組待檢測細胞用胰蛋白酶處理,然后收集至離心管并離心,用PBS重懸細胞。行細胞計數后取出1×105個細胞,再次離心棄上清液。細胞中先后加入400 μL的Annexin V-FITC檢測液,充分混勻并重懸細胞。將細胞置于37 ℃孵箱避光靜置約30 min。離心棄上清后,加入400 μL PBS輕輕重懸待檢測細胞,并加入10μL的PI,充分混合均勻,靜置在冰盒中20 min。然后用流式細胞儀檢測并統計分析細胞凋亡情況。
1.2.5 MTT法檢測細胞增殖 收集對照組和Orai2 shRNA組細胞至離心管;棄上清液后重懸細胞,然后進行細胞計數,取1×104/mL細胞懸液100 μL滴加到96孔細胞培養板中。分別培養24 h、48 h、64 h后,棄上清液在細胞培養板的每個孔中滴加50 μL MTT和150 μL細胞培養基。然后將細胞培養板在37 ℃靜置。4 h后棄上清,再在細胞培養板的每個孔中滴加200 μL的DMSO溶液溶解MTT 30 min。最后,用酶標儀在570 nm測各個孔的吸光值并統計學分析實驗數據。
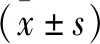
2 結 果
2.1 應用靶向Orai2的shRNA在MC3T3-E1成骨細胞中沉默Orai2的表達 為了明確Orai2在骨形成過程中是否發揮作用,我們在成骨誘導條件下用Western blot和Real-Time PCR的方法檢測Orai2的mRNA轉錄和蛋白表達。Western blot檢測結果發現,成骨誘導后第7天、第14天和第21天,Orai2的蛋白表達水平是逐漸增加的,結果差異有統計學意義(見圖1)。同樣,Real-Time PCR的結果也發現,成骨誘導后第7天、第14天和第21天,Orai2的mRNA轉錄水平也是逐漸增加的,結果差異有統計學意義(見圖2)。以上結果發現成骨細胞分化過程中Orai2的蛋白表達水平逐漸增加,提示:Orai2可能在骨形成過程中發揮了作用。然后我們用Western blot和Real-Time PCR的方法檢測轉染了靶向Orai2的shRNA后Orai2的表達。Western blot檢測結果發現,在轉染了靶向Orai2的shRNA后,Orai2的蛋白表達出現了明顯降低,結果差異有統計學意義(見圖3)。同樣,Real-Time PCR的結果也發現,在轉染了靶向Orai2的shRNA后,Orai2的mRNA轉錄水平出現了顯著降低,結果差異有統計學意義(見圖4)。以上結果提示:我們應用靶向Orai2的shRNA在MC3T3-E1成骨細胞中可成功沉默Orai2的表達。

注:*表示與成骨誘導后0 d比較,差異有統計學意義 注:*表示與成骨誘導后0 d比較,差異有統計學意義圖1 成骨誘導后0、7、14、21 d,檢測成骨細胞Orai2蛋白表達水平 圖2 成骨誘導后0、7、14、21 d,檢測成骨細胞Orai2 mRNA轉錄水平

注:*表示與對照組比較,差異有統計學意義 注:*表示與對照組比較,差異有統計學意義圖3 細胞轉染Orai2 shRNA后,檢測成骨細胞Orai2蛋白表達水平 圖4 細胞轉染Orai2 shRNA后,檢測成骨細胞Orai2 mRNA轉錄水平
2.2 沉默Orai2表達抑制成骨細胞增殖 為了研究Orai2能否參與調控成骨細胞的增殖能力,我們應用靶向Orai2的shRNA在MC3T3-E1成骨細胞中沉默Orai2的表達后,應用MTT法檢測成骨細胞增殖能力的變化。研究發現,與轉染無關干涉片段的對照組相比較,沉默Orai2表達后,成骨細胞的增殖能力明顯降低,結果差異有統計學意義(見圖5)。為了進一步明確Orai2能否參與調控成骨細胞的增殖,我們采用PI染色,流式細胞儀檢測成骨細胞在沉默Orai2表達后細胞周期的變化。結果發現,與轉染無關干涉片段的對照組相比較,沉默Orai2表達后,細胞周期的G0/G1期明顯增加,而細胞周期的S期明顯減少,結果差異有統計學意義(見圖6)。以上結果證實,沉默Orai2的表達可在成骨細胞抑制DNA合成,減少細胞進入S期,抑制成骨細胞的增殖,從而導致成骨細胞的數量減少。

注:*表示與對照組比較,差異有統計學意義圖5 細胞轉染Orai2 shRNA后0、1、2、3、4 d,MTT法檢測Orai2對細胞增殖能力的影響
2.3 沉默Orai2表達促進成骨細胞凋亡 為了研究Orai2的表達異常可否參與成骨細胞凋亡的調控,我們應用靶向Orai2的shRNA在MC3T3-E1成骨細胞中成功沉默Orai2的表達后,采用PI/AnnexinⅤ法檢測成骨細胞凋亡的變化。結果發現,與轉染無關干涉片段的對照組相比較,沉默Orai2表達后,細胞的凋亡比例明顯增加,并且差異有統計學意義(見圖7,P<0.05)。以上結果提示:沉默Orai2的表達可在成骨細胞促進其細胞凋亡,從而導致成骨細胞的數量減少。
2.4 沉默Orai2表達抑制成骨細胞的分化 成骨細胞分化抑制導致成骨細胞數量減少也是骨質疏松癥發生發展的重要原因。Runt相關轉錄因子2(runt-related transcription factor2,Runx2)和鋅指結構轉錄因子(osterix)是反映成骨

注:*表示與對照組比較,差異有統計學意義圖6 細胞轉染Orai2 shRNA后,檢測Orai2對成骨細胞細胞周期的影響

注:*表示與對照組比較,差異有統計學意義圖7 細胞轉染Orai2 shRNA后,檢測Orai2對成骨細胞凋亡影響
細胞成骨分化能力的關鍵轉錄因子[10-12]。為了研究Orai2可否參與成骨細胞分化的調控,我們應用靶向Orai2的shRNA在MC3T3-E1成骨細胞中成功沉默Orai2的表達后,分別在成骨誘導后第7天、14天和21天采用Real Time-PCR方法檢測細胞中Runx2和Osterix這兩個關鍵轉錄因子的mRNA轉錄水平。結果顯示,與轉染無關干涉片段的對照組相比較,沉默Orai2表達后,成骨細胞Runx2和Osterix的mRNA轉錄水平都明顯降低,結果差異有統計學意義(見圖8)。堿性磷酸酶(alkaline phosphatase,ALP)是反映成骨分化的另一重要指標,隨著成骨細胞分化程度的增加,堿性磷酸酶的表達也隨之增加。為了進一步明確Orai2在成骨細胞分化過程中的作用,我們用Real-Time PCR的方法檢測了在成骨過程中Orai2對堿性磷酸酶轉錄的影響。我們發現在成骨分化的早期和晚期,沉默Orai2表達都會顯著抑制堿性磷酸酶的轉錄,結果差異有統計學意義。提示:沉默Orai2的表達可在成骨分化的早期和晚期都明顯抑制成骨細胞的成骨分化能力(見圖9,P<0.05)。這些結果證實沉默Orai2的表達可明顯抑制成骨細胞的成骨分化能力,從而導致成骨細胞的數量減少。

注:*表示與對照組比較,差異有統計學意義 注:*表示與對照組比較,差異有統計學意義圖8 Real-Time PCR檢測轉染Orai2對Runx2和Osterix mRNA轉錄水平的影響 圖9 Real-Time PCR檢測轉染Orai2對堿性磷酸酶(ALP)的mRNA轉錄水平的影響
2.5 沉默Orai2表達抑制成骨細胞Ras-ERK1/2信號通路活性 已有研究發現抑制Ras-ERK1/2信號通路后,成骨細胞會出現增殖減少、凋亡增加以及分化異常等現象。Orai2是定位于細胞膜上的一類介導受體依賴型鈣離子通道,它的主要功能是介導受體依賴型的鈣離子內流。鈣離子進入細胞后會發揮第二信使的作用調控細胞增殖、分化、凋亡等細胞功能。研究發現,鈣離子作為第二信使能直接與Ras-GTP結合而顯著增加Ras-ERK1/2信號通路的活性[13-15]。為了研究Orai2在成骨細胞可否激活Ras-ERK1/2信號通路參與調控成骨細胞的增殖、凋亡以及分化等生物學功能,我們應用靶向Orai2的shRNA在MC3T3-E1成骨細胞中成功沉默Orai2的表達后,然后用Western blot檢測成骨細胞Ras與ERK1/2的表達與活性。我們發現,與轉染無關干涉片段的對照組相比較,沉默Orai2表達后,細胞內Ras與ERK1/2的活性形式Ras-GRF和p-ERK1/2都明顯降低,結果差異有統計學意義(見圖10)。這些結果證實在成骨細胞沉默Orai2表達可明顯抑制Ras-ERK1/2信號通路的活性。

注:*表示與對照組比較,差異有統計學意義圖10 細胞轉染Orai2 shRNA后,檢測Orai2對成骨細胞Ras與ERK1/2的活性的影響
3 討 論
目前,大量研究證實骨質疏松癥發生發展的主要原因包括成骨細胞增殖減少、成骨細胞凋亡增加以及成骨細胞分化異常[1-5]。高度礦化的骨基質由成骨細胞組成。成骨細胞可介導鈣和磷酸鹽沉積到I型膠原基質。已有研究證實細胞外鈣離子濃度增加可以增強體外培養的人間充質干細胞的成骨作用。研究發現,維生素D是鈣穩態和骨礦化的主要調節因子,可在體外促進成骨分化[16-18]。當細胞外液的鈣離子濃度增加時,成骨細胞的增殖、分化以及細胞外基質鈣化等成骨能力都顯著增加,并且凋亡減少[19-21]。還有研究發現,骨質疏松癥患者,由于破骨細胞過度激活,成骨細胞的細胞外液鈣離子水平增加了將近20倍[22]。然而,雖然細胞外液的鈣離子水平顯著增加,但骨質疏松癥患者成骨細胞的成骨能力卻仍處于一個較低水平[23-24],其機制未見報道。細胞外液的鈣離子從胞外進入胞內是鈣離子發揮第二信使作用的主要方式。細胞膜是半通透膜,鈣離子不能自由順濃度梯度擴散,而是需要依賴于細胞膜上的鈣離子載體,也被稱為鈣離子通道。因此,細胞膜上鈣離子通道的表達或者功能異常導致鈣離子內流減少,可能會是成骨細胞在高鈣的細胞外液中卻依然處于較低成骨分化能力的原因。
最近發現了一種細胞膜上鈣通道Orai蛋白,包括Orai1蛋白和Orai2蛋白,其在鈣離子內流過程中發揮了重要的作用。當內質網中的鈣儲存耗盡時,Orai蛋白被激活,允許快速而短暫的鈣離子從胞外流入細胞質,稱為鈣庫操作鈣內流。基因敲除Orai1可導致骨骼發育受損,并出現骨量減少,骨礦物質密度降低[6-9]。然而,Orai2對成骨細胞增殖、凋亡及分化的影響尚未見報道,這限制了我們對Orai蛋白在骨生物學中的作用理解。成骨細胞增殖的減少、凋亡的增加以及分化的異常導致成骨細胞數量減少是骨質疏松癥的發生發展的重要原因。為了研究Orai2是否通過調控成骨細胞的增殖、凋亡及分化等細胞功能參與骨質疏松癥的發生發展,我們在小鼠成骨細胞系MC3T3-E1轉染了靶向Orai2的shRNA來沉默Orai2的表達。我們研究發現,在小鼠成骨細胞系MC3T3-E1沉默Orai2的表達后,成骨細胞的增殖明顯減少。細胞增殖是通過細胞周期來實現的。細胞周期的S期為DNA合成期,是反映細胞增殖能力的重要指標。為了進一步明確Orai2能否參與調控成骨細胞的增殖,我們在成骨細胞沉默Orai2表達后,檢測了細胞周期的變化。結果發現,沉默Orai2表達后,成骨細胞的G0/G1期的細胞比例明顯增加,而S期的細胞比例明顯減少。這些結果提示抑制Orai2的表達可抑制成骨細胞的增殖,從而減少成骨細胞的數量。細胞凋亡的水平是決定成骨細胞數量的另一重要指標。我們在MC3T3-E1成骨細胞中沉默Orai2的表達后,成骨細胞的凋亡比例明顯增加。這提示Orai2的表達可在成骨細胞發揮抗細胞凋亡的作用,從而增加成骨細胞的數量。骨髓間充質干細胞是具有成脂分化、成軟骨分化和成骨分化多向分化潛能的多能干細胞。成骨分化能力的降低也會顯著減少成骨細胞的數量。Runx2和Osterix是成骨細胞成骨分化的兩個關鍵的特異性轉錄因子。研究發現,在骨髓間充質干細胞同時或者分別敲除Runx2和Osterix后,都可導致成骨分化抑制或者骨形成不全[10-11]。另有研究發現在Osterix基因敲除小鼠仍可檢測到Runx2基因的表達,然而在Runx2基因敲除小鼠中卻檢測不到Osterix基因的表達。所以,Runx2位于Osterix上游而發揮調控成骨分化的作用。但是Runx2和Osterix在成骨細胞分化過程如何相互作用,目前仍未研究清楚。堿性磷酸酶是反映成骨分化的另一重要指標,隨著成骨細胞分化程度的增加,堿性磷酸酶的表達也隨之增加[12]。我們研究發現,在小鼠成骨細胞系MC3T3-E1沉默Orai2的表達后,成骨細胞Runx2和Osterix以及堿性磷酸酶的mRNA轉錄水平都明顯降低,提示沉默Orai2的表達可明顯抑制成骨細胞的成骨分化能力,從而導致成骨細胞的數量減少。以上研究結果證實,Orai2的表達降低可減少成骨細胞增殖、增加成骨細胞凋亡、抑制成骨分化,從而減少成骨細胞的數量。因此,Orai2對于成骨細胞數量的維持至關重要。研究發現抑制Ras-ERK1/2信號通路后,成骨細胞出現增殖減少、凋亡增加以及分化異常等現象。Orai2是定位于細胞膜上的一類介導受體依賴型鈣離子通道,它的主要功能是介導受體依賴型的鈣離子內流。鈣離子進入細胞后會發揮第二信使的作用而激活多條信號通路調控細胞功能。鈣離子作為第二信使能直接與Ras-GTP結合而顯著增加Ras-ERK1/2信號通路的活性[13-14]。有研究發現,ERK1/2在成骨細胞內可直接入核結合Runx2啟動子區并顯著促進Runx2的表達[15]。我們在本研究中發現,在小鼠成骨細胞系MC3T3-E1沉默Orai2的表達后,Ras-ERK1/2信號通路活性明顯下降。這說明Orai2的表達抑制可顯著降低Ras-ERK1/2信號通路活性,并減少成骨細胞增殖、增加成骨細胞凋亡、抑制成骨分化。
綜上所述,我們的研究發現,在成骨細胞中沉默Orai2的表達可導致成骨細胞增殖減少、凋亡增加以及分化抑制,從而減少成骨細胞的數量。Orai2的表達異常或者功能異常可能是導致骨質疏松癥發生發展的重要原因。因此,深入研究Orai2在骨質疏松癥發生中的作用,可為骨質疏松癥的生物治療提供新的思路。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
