祁連山高寒草原主要植物種群格局對冬季放牧的響應
2019-12-18 07:25:20彭澤晨侯扶江
生態學報 2019年21期
武 哲,彭澤晨,侯扶江
蘭州大學草地農業生態系統國家重點實驗室,蘭州大學農業部草牧業創新重點實驗室,蘭州大學草地農業科技學院, 蘭州 730020
放牧不僅是對草原最重要的管理方式,也是影響草原群落結構的重要途徑之一[1-6]。放牧家畜是人類日常生活中肉、奶以及皮革的重要來源[7-10]。家畜活動對于草地群落有3種主要影響方式:踐踏、采食與排泄[11-12]。放牧管理對于草原植物種群格局的研究始終是熱點[13-18]。在藏北高寒草原放牧試驗表明中度放牧可提高物種多樣性,重度放牧造成紫花針茅草地過渡到雜類草草地[19];長期重度放牧還會致使草原物種植株矮化,引起草原群落退化[20]。
種群分布格局是指種群在空間上的配置特點和分布狀況[21],是由種群生物學特性、種群間相互關系和種群與環境關系共同作用形成[22-23]。植物種群空間格局的研究有助于了解植物-環境之間的互作規律[24],揭示種群在群落中的地位與綜合生存能力[25]。種群格局計盒維數 (Box-counting dimension)揭示了種群對生態空間的占據能力和利用程度[26-27];而種群格局信息維數 (Information dimension)則從獨立于尺度的角度,揭示了格局強度的尺度變化以及個體分布的均勻狀況[28-29]。
紫花針茅,作為家畜采食的主要牧草,其種群特征反映牧場優質牧草的含量;而不能被家畜采食的醉馬草種群格局直接說明草地退化程度的高低。目前,國內學者采用分形理論對高寒草原的種群空間格局少有研究且多采用部分要素試驗方案分析。這些研究在野外實施時難以達成理想效果,而本研究采用全要素試驗方法觀測連續放牧率對高寒草地的調控,可以避免此問題。祁連山中段肅南鹿場冬季牧場為研究對象,因其地處高寒、環境惡劣,草原群落格局對放牧率的響應尤其敏感。通過研究不同放牧率下紫花針茅與醉馬草的種群格局,分析變化緣由,提出適合牧場管理的方案。
1 材料與方法
1.1 研究區概況
研究區位于甘肅省肅南裕固族自治縣鹿場,地處祁連山北麓中段。地理坐標為38.8°N,99.6°E,平均海拔2850 m。年均溫3.6 ℃,月均溫最高為15.9℃(7月份)。年均降水量253.0 mm,月均降水最多為60.6 mm(7月份)。無霜期長達127 d,只有冷暖兩季之分,水熱同步,均集中于5—9月。牧草4月下旬返青,7月上旬進入生長旺盛期,9月中上旬開始枯黃。土壤類型為山地栗鈣土。根據草原綜合順序分類法,草原類型為寒溫微干山地草原類。
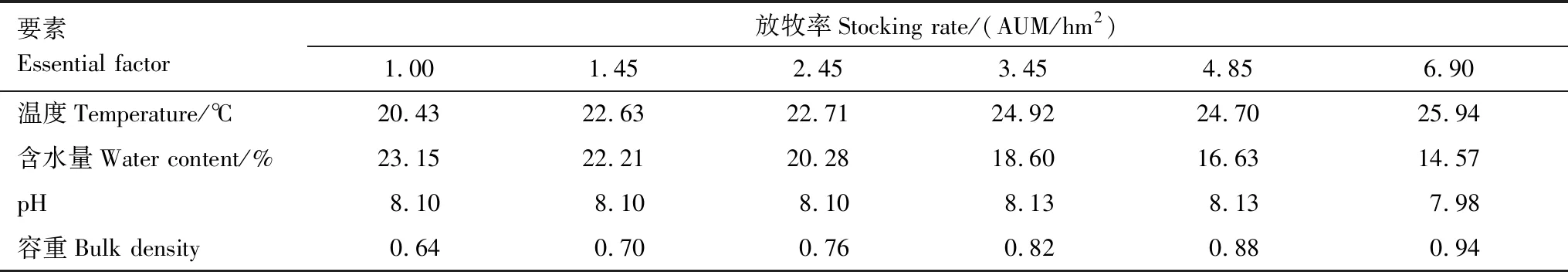
表1 冬季牧場土壤基本要素
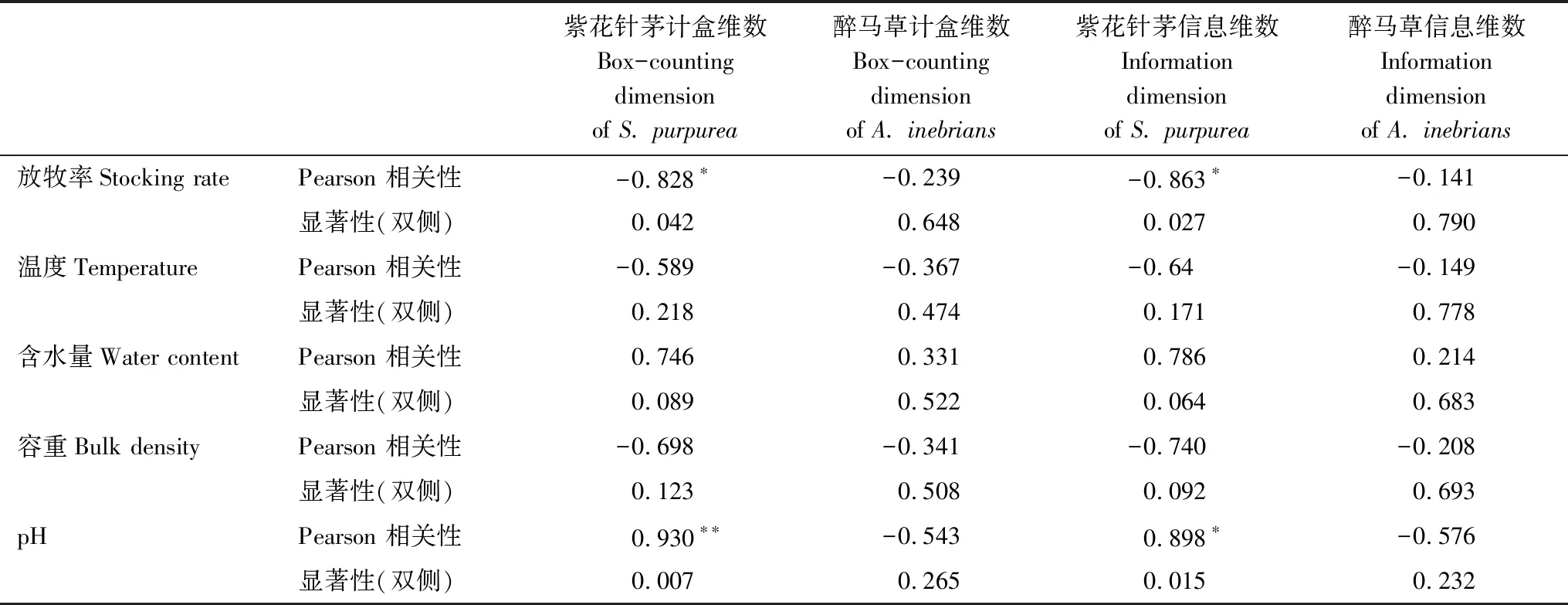
表2 冬季牧場土壤基本要素與植物指標相關性分析
*在0.05 水平雙側上顯著相關;**在0.01水平雙側上極顯著相關
1.2 樣地設置
肅南鹿場冬季牧場地形開闊平坦,整體坡度<3°,面積約160 hm2,每年11月下旬至翌年4月放牧馬鹿。草原主要物種是紫花針茅、扁穗冰草、賴草等,退化地段主要有醉馬草和銀灰旋花等[30]。
通過放牧行為觀測,確認馬鹿放牧的主要采食路徑[31],結合植物生長狀況,以牧場進出口為起點,向外呈放射狀地形成放牧率由高到低的牧壓梯度。選擇3條主要路徑,分別在距牧場出入口0、300、600、900、1200、1500 m處設置樣地,每塊樣地面積約為1 hm2,標明邊界。
試驗選擇遠距離跟蹤觀測馬鹿。根據試驗需求,選擇標記15匹成年馬鹿進行為期15天跟蹤觀測。每天從放牧到歸牧,每2個小時進行15 min的間隔觀測,記錄馬鹿的放牧行為及所處位置。將已觀測的馬鹿放牧行為換算為連續放牧草地的放牧率。公式如下[32-34],
式中,SRi為樣地i的放牧率;SRm為牧場的平均放牧率,為單位時間內牧場中家畜單位與牧場面積的比值;∑Hdi為樣地i家畜出現頻次的總和;μi=1:n∑Hdi為所有樣地家畜出現頻次總和的均值;n為樣地總數。測定各年齡段馬鹿的活體重,換算為家畜單位。本試驗,一個家畜單位為一頭成年甘肅馬鹿母鹿(活體重200 kg左右),帶一只哺乳期的小鹿。從牧場出入口向遠處的6個樣地對應的放牧率分別為: 6.90、4.85、3.45、2.45、1.45 AUM/hm2和1.00 AUM/hm2。
1.3 植物測定與數據分析
于2000年7月底至8月初,進行種群的偏離系數分布格局調查。在各個樣地中隨機選取10個0.25 m2的樣方,測量每種植物的個體數和生物量;并在8月上旬每個樣地設置l個2 m×2 m樣方,進行種群格局的分形分析。
各植物物種根據偏離系數(Deviation coefficient)對試驗結果進行統計檢驗。偏離系數以 Poisson分布的預期假設為基礎,不受數據的測量尺度差異或者數據量綱不同的影響。
式中,X為樣方中每種植物的個體數,n為各樣地中樣方總數。Cx=1時,種群為隨機分布;Cx<1,種群為均勻分布;Cx>1,種群為集群分布。采用t檢驗時,首先判別種群是否偏離Poisson分布。如果差異不顯著,則為隨機分布。如果差異顯著,則根據t值的正負分別作上尾或下尾單側檢驗。t>0,則作上尾單側檢驗;t<0,則作下尾單側檢驗。如果上側檢驗顯著,則Cx> 1,種群為集群分布;如果下尾檢驗顯著,則Cx< 1,種群為均勻分布[35]。
采用Ripley′s K函數以及蒙特卡羅(Monte-Carlo)隨機模擬方法[36],定量分析植物種群點格局變化,
式中,h是空間尺度;K(h)是h為半徑的圓內小格子的數目;λ是種群密度;R為樣方的面積;N是樣方中有冷蒿出現的小格子的總數目;dij表示第i個小格子和第j個小格子之間的距離;Ih(dij)是指示函數。
計盒維數反映種群占據空間的能力[37-38]。測定計盒維數,采用不同的劃分尺度(ε,格子邊長)對樣方逐次刪格化,統計對應于尺度ε的非空格子數N。計盒維數(Db)是ε→0時非空格子數 (N)與劃分尺度(ε)負的對數比,可以簡便地將N與ε在雙對數坐標下進行直線(或分段直線)擬合,以擬合直線斜率的絕對值為計盒維數的估計值,表征種群占據空間的尺度。
信息維數揭示不同尺度上種群個體分布的均勻程度,拐點顯示這種變化規律發生顯著轉換的尺度[39-40]。信息維數在對種群實施網格覆蓋過程中,考慮到每個格子中的個體數目,通過信息量公式給出每一尺度對應的信息量冪律關系。
式中,N是樣方個體總數目,Ni是非空格子i的個體數目,Pi為格子i的概率,ε為研究尺度,Db為信息維數,Di為信息維數。
1.4 數據統計分析
試驗數據用Excel 2007進行初步整理、作圖,用SPSS 20.0進行統計分析。
2 結果分析
2.1 冬季牧場物種分布格局
冬季牧場植物種群格局在放牧梯度上的變化有5種情況(表3):①種群在各放牧率下呈單一集群分布格局,如黑褐穗苔草與堿韭;②種群在各放牧率下呈單一隨機分布格局,如乳白黃耆、星毛委陵菜等;③隨著放牧率增加,分布格局逐漸由隨機分布過渡到集群分布,如紫花針茅、冷蒿等;④種群分布格局隨放牧增強由集群分布過渡到隨機分布,如扁穗冰草、刺芒龍膽等;⑤放牧梯度上種群集群分布與隨機分布交替出現,如阿爾泰狗娃花與賴草。黑褐穗苔草與堿韭的耐踐踏、耐放牧而競爭能力低,形成單一集群分布格局。乳白黃耆與蚓果芥的零星分布,星毛委陵菜的匍匐分枝,波伐早熟禾的分蘗明顯,醉馬草的不可食,使它們的種群格局依舊保持隨機分布。紫花針茅、紫花棘豆與冷蒿的優良適口性以及銀灰旋花的不耐踐踏造成他們種群由隨機分布過渡到集群分布。刺芒龍膽與灰綠藜的不耐牧性、扁穗冰草的過度被采食,使得三者的種群格局由集群分布轉變為隨機分布。
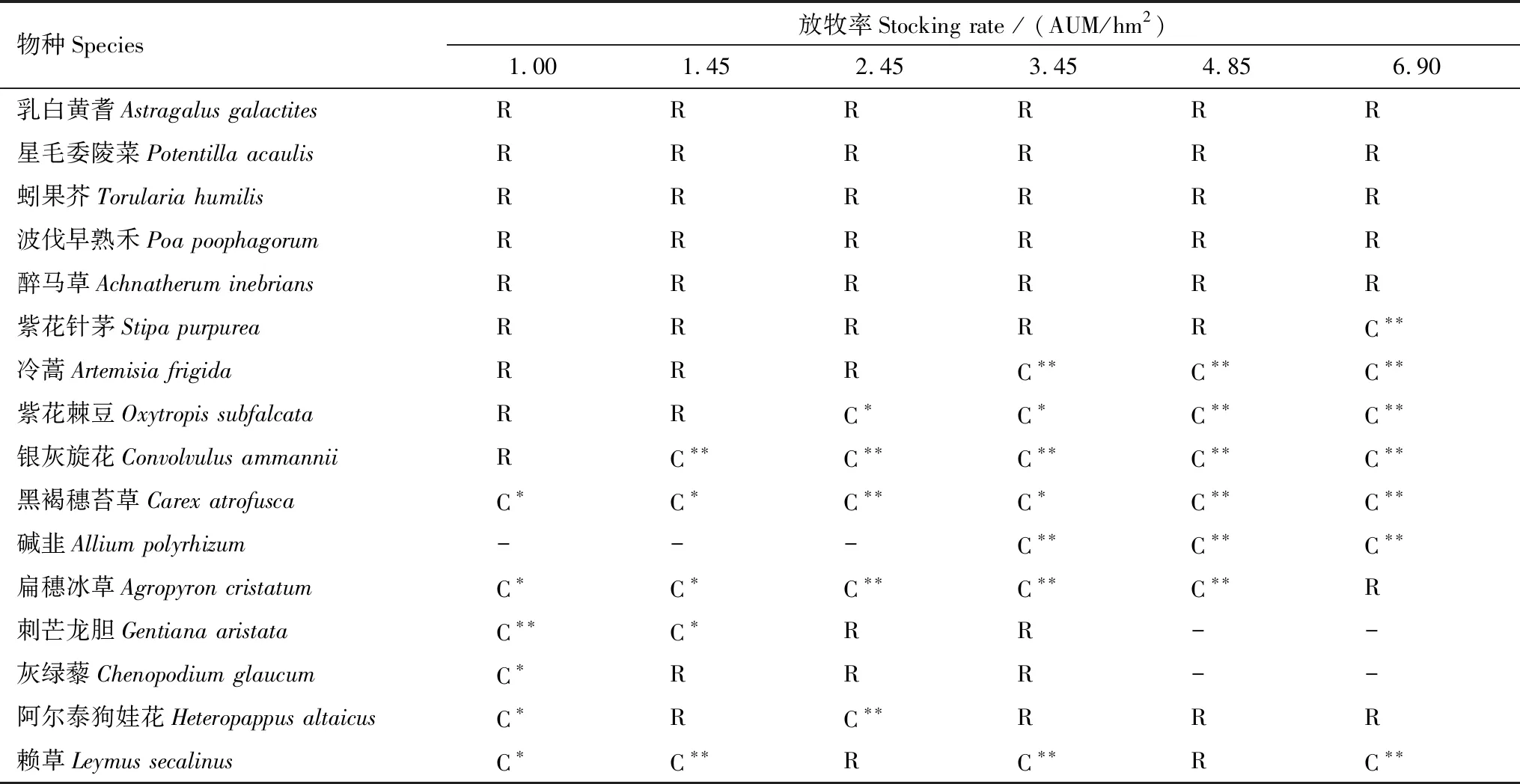
表3 各放牧率下各種群格局的變化
C 集群分布;R隨機分布;*表示各種群偏離系數t檢驗值差異極顯著(P<0.05);**表示各種群偏離系數t檢驗值差異極顯著(P<0.01)
種群的偏離系數作為種群集群程度的統計指標,其大小反映各物種集群程度的高低。在放牧梯度上,其變化主要有4種情況(圖1):①隨放牧增強而上升,如紫花棘豆與冷蒿呈線性升高,而黑褐穗苔草呈指數增大;②隨放牧增大而減少,如刺芒龍膽與灰綠藜呈對數下降;③在放牧梯度上呈凸拋物線變化,存在極大值,如蚓果芥、乳白黃耆、波伐早熟禾、扁穗冰草、銀灰旋花以及醉馬草;④在放牧梯度上呈凹拋物線變化,存在極小值,如紫花針茅。冷蒿作為多年生草本牧草,具備很強的生根萌蘗再生能力。隨著放牧率的增大,扁穗冰草的營養枝數量增加,冷蒿的生殖枝數量下降,匍匐生長,枝條出現不定根并刺激更新芽的生根與萌蘗,生殖方式由有性生殖過渡到無性生殖,斑塊分布越發明顯,種群格局由隨機分布過渡到集群分布[41]。同時,被采食的冷蒿會釋放揮發性有機化合物極顯著地抑制扁穗冰草幼根的伸長與須根的發育,造成扁穗冰草的凱氏帶發育不完全[42],成為扁穗冰草種群格局由集群分布過渡到隨機分布的原因之一。
冬季牧場禾本科植物主要有:紫花針茅、醉馬草、波伐早熟禾、扁穗冰草、賴草。其中,醉馬草、波伐早熟禾均與紫花針茅的種群偏離系數呈顯著負相關關系,紫花針茅與醉馬草、波伐早熟禾存在明顯的種間競爭。紫花針茅、醉馬草、波伐早熟禾、扁穗冰草、賴草的種群偏離系數區間分別為[0.11,3.18]、[0.25,2.50]、[0.39,1.90]、[2.00,12.39]、[0.44,10.54],種群偏離系數極差分別為3.07,2.25,1.51,10.39,10.10。醉馬草、波伐早熟禾的種群偏離系數波動極低,種群格局依舊。紫花針茅的種群偏離系數極差3.07,波動略高,在放牧率6.90 AUM/hm2下,出現最大值3.18,亦發生種群格局由隨機分布改變為集群分布。扁穗冰草在天然草地中采用集群分布,在放牧干擾下,重度放牧使得種群格局轉變為隨機分布。賴草為冬季牧場偶見種,其種群格局無明顯變化規律。黑褐穗苔草作為主要伴生牧草,其種群偏離系數受放牧率影響顯著。
冬季牧場主要有紫花棘豆與乳白黃耆兩種豆科植物。乳白黃耆的種群偏離系數區間為[0.75,2.00],極差1.25;紫花棘豆的種群偏離系數區間為[0.33,15.69],極差15.36。乳白黃耆的種群偏離系數波動小,格局保持隨機分布。紫花棘豆的種群偏離系數波動大,與放牧率正相關,出現隨機分布向集群分布轉變。
冬季牧場也出現3種反映放牧率高低的指示植物。刺芒龍膽和灰綠藜出現在放牧率≤3.45 AUM/hm2的樣地(表3)。隨放牧率增大,種群偏離系數都迅速下降,集群程度急速降低,種群格局由集群分布過渡到隨機分布。而堿韭在≥3.45 AUM/hm2的樣地中出現,堿韭的偏離系數隨放牧率增大平緩上升,集群度變化相對較少(圖1)。
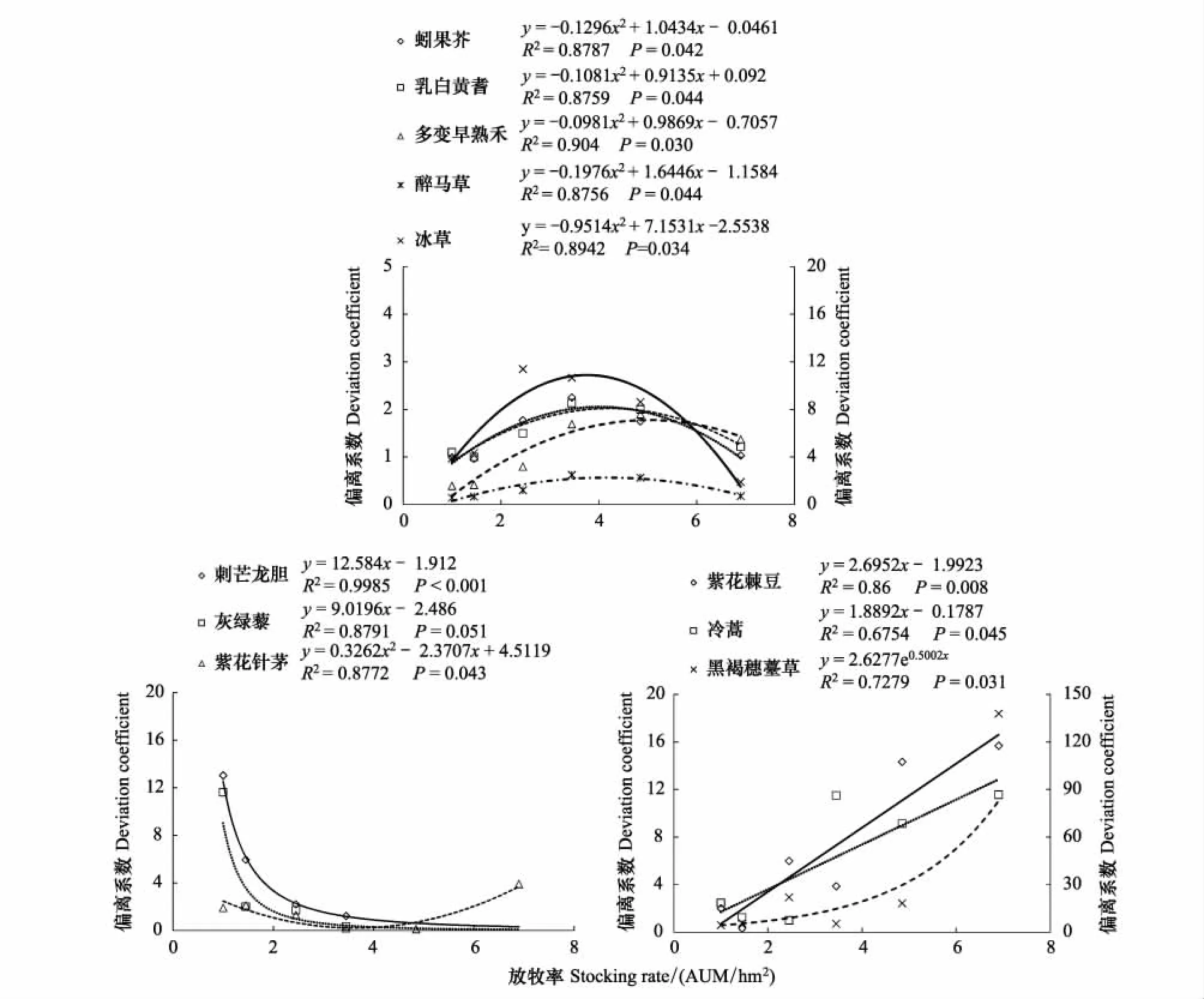
圖1 各放牧率下每種植物的種群偏離系數變化Fig.1 Changes of population deviation coefficient with individual plant under different stocking rates 扁穗冰草、黑褐穗苔草為次縱坐標軸
采用Ripley′s K函數,對紫花針茅種群與醉馬草種群進行點格局分析。在0—1 m的研究尺度內,紫花針茅種群與醉馬草種群的Ripley′s K函數結果均在蒙特卡洛隨機模擬區間內,呈隨機分布格局。紫花針茅種群在放牧率6.90 AUM/hm2時,尺度0.04 m上出現L(d)>0(圖2)。醉馬草種群在放牧率4.85 AUM/hm2時尺度區間[0.02,0.12]和在放牧率6.90 AUM/hm2時尺度區間[0.02,0.08],出現L(d)>0(圖3)。偏離系數及t檢驗方法與Ripley′s K函數及蒙特卡羅隨機模擬試驗,均可定量分析種群分布格局。偏離系數及t檢驗方法直接反映種群的定量格局,并可進行不同放牧率的統計分析。Ripley′s K函數及蒙特卡羅隨機模擬試驗采用點格局分析方法,研究種群在不同尺度內的變化。兩者相比,Ripley′s K函數及蒙特卡羅隨機模擬試驗可具體分析種群變化尺度,偏離系數及t檢驗方法便于放牧率尺度的研究。
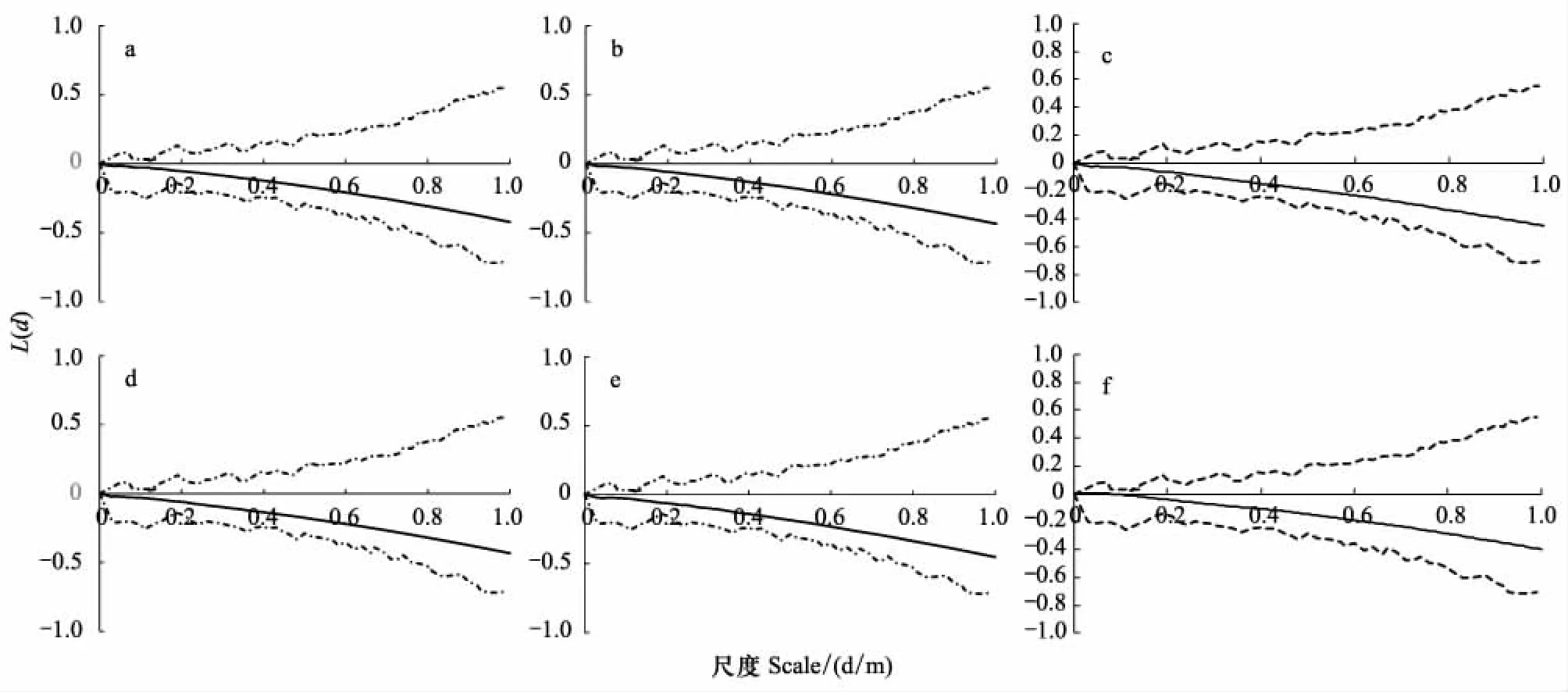
圖2 在0—1 m尺度上,紫花針茅的L(d)隨尺度d的變化Fig.2 Changes of L(d) about S. purpurea with study scales under 0—1m 實水平線是完全空間隨機條件下L(d)-d的0參考值,虛線是99次Monte-Carlo模擬估計得到的L(d)-d的2.5%和97.5%分位數;a、b、c、d、e、f分別為放牧率: 6.90、4.85、3.45、2.45、1.45、1.00 AUM/hm2
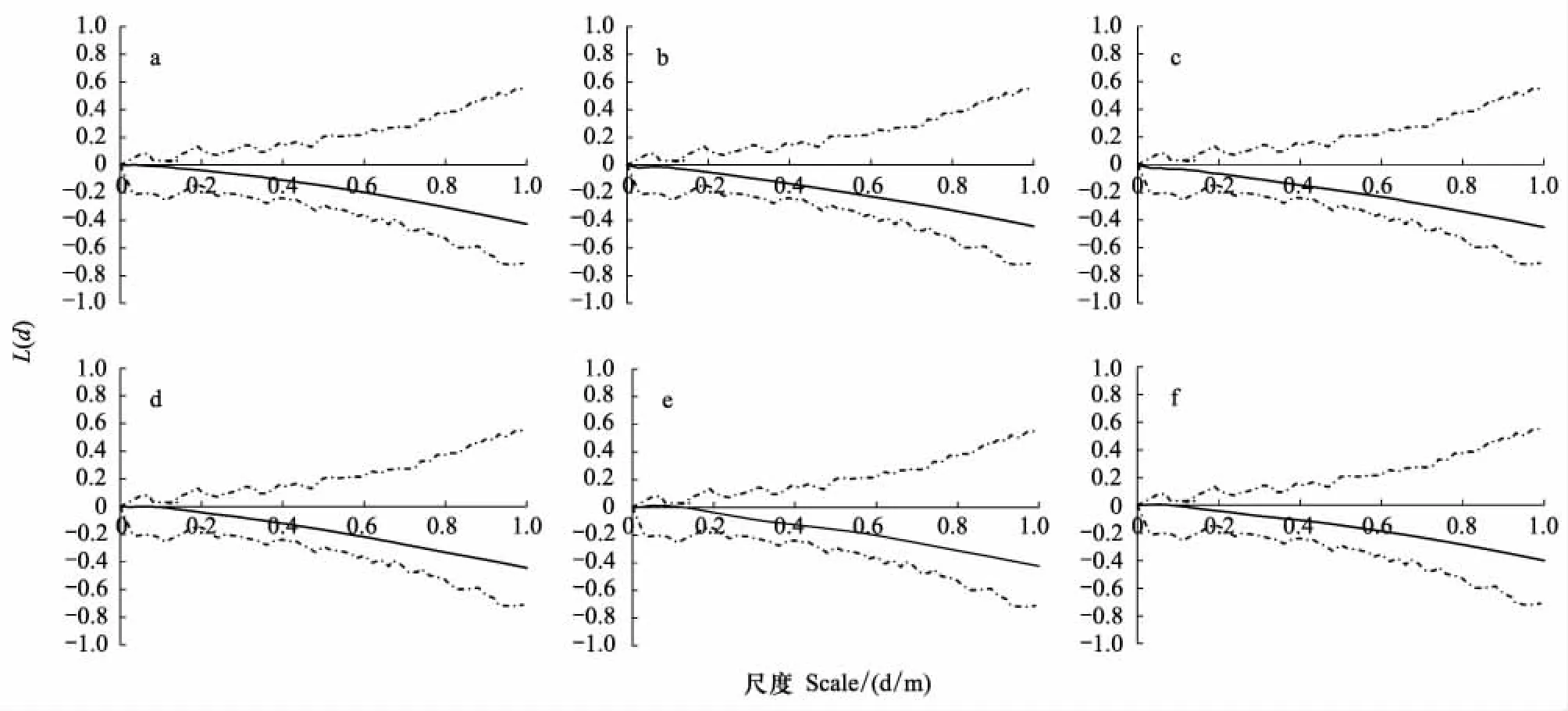
圖3 在0—1m尺度上,醉馬草的L(d)隨尺度d的變化Fig.3 Changes of L(d) about A. inebrians with study scales under 0—1m 實水平線是完全空間隨機條件下L(d)-d的0參考值,虛線是99次Monte-Carlo模擬估計得到的L(d)-d的2.5%和97.5%分位數;a、b、c、d、e、f分別為放牧率: 6.90、4.85、3.45、2.45、1.45、1.00 AUM/hm2
2.2 優勢物種種群分形特征
2.2.1計盒維數
種群格局的變化,歸因于物種生存能力的改變。計盒維數是反映物種生存能力的指數之一,主要說明種群的占據空間能力。在各放牧率下,紫花針茅與醉馬草的種群計盒維數回歸擬合R2≥0.968,P<0.01,在群落尺度內存在自相似性(表4)。
紫花針茅的種群計盒維數區間為[1.596,1.921],極差0.325;醉馬草的種群計盒維數區間為[1.831,1.945],極差0.114。紫花針茅與醉馬草的計盒維數都接近2,表明均占據極大的生態空間,與其在群落中的主導作用相一致。種群的計盒維數極差,反映對外界因素干擾種群抵抗力的強弱。紫花針茅種群的計盒維數極差更大,反映受放牧影響紫花針茅空間占據力發生更明顯改變。
紫花針茅種群的計盒維數與放牧率可曲線擬合(y=-0.0197x2+ 0.1018x+ 1.839,R2=0.9831,P<0.01),而醉馬草種群的計盒維數則隨放牧率變化沒有明顯規律。在放牧率1.967 AUM/hm2時,紫花針茅種群計盒維數達到最大,占據空間最大。紫花針茅與醉馬草的計盒維數存在顯著負相關(R2=0.835,P<0.05),同種群偏離系數的負相關結果相同。表明,紫花針茅與醉馬草的生存競爭激烈。放牧與醉馬草種間競爭成為紫花針茅種群格局改變的主要因素;醉馬草種群格局的直接因素為紫花針茅,間接因素為放牧。結果表明,紫花針茅的種間競爭能力強于醉馬草,而醉馬草對放牧環境的適應能力優于紫花針茅。
紫花針茅與醉馬草兩者的拐點變化隨放牧率的增大發生相反改變(表4),即紫花針茅為“峰”值,則醉馬草為“谷”值,反之亦然,呈負相關關系。拐點尺度反映出種群聚塊尺度,在放牧相對較重的6.90 AUM/hm2和4.85 AUM/hm2樣地,紫花針茅冠幅小于聚塊尺度,表明聚塊由小個體組成,因個體生長不良所致;放牧相對較輕的1.00 AUM/hm2和1.45 AUM/hm2樣地則相反,冠幅大于聚塊尺度,聚塊由生長良好的大個體組成。結果表明,馬鹿重牧可以為醉馬草提供相適應的微氣候環境,壓制紫花針茅種群的生長。
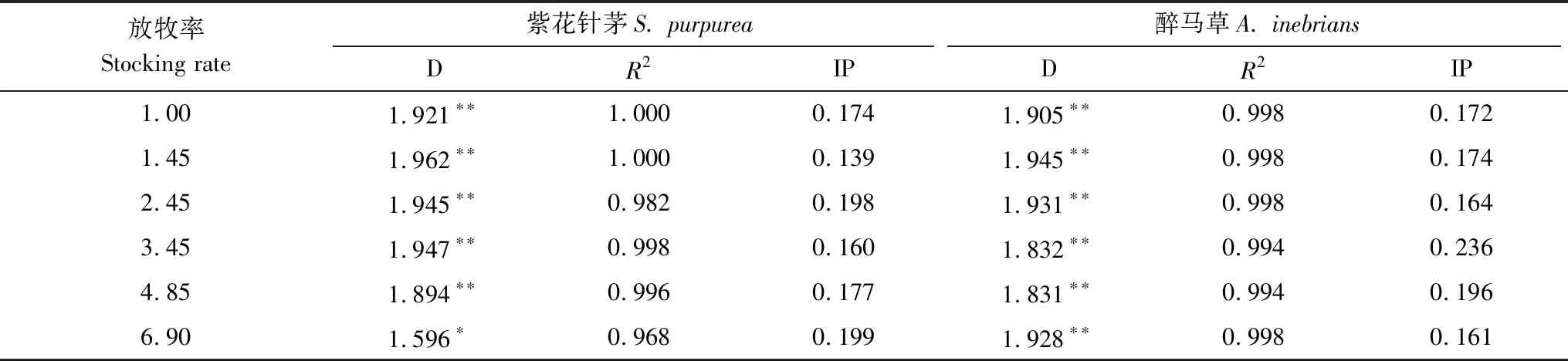
表4 各放牧率下植物種群空間分布格局的計盒維數
D表示計盒維數;R2表示計盒維數的相關系數;IP表示計盒維數的拐點尺度;*表示計盒維數差異極顯著(P<0.05);**表示計盒維數差異極顯著(P<0.01)
2.2.2信息維數分析牧場優勢草種的個體均勻度
種群格局的變化,主要表現在種群分布的均勻度改變。信息維數揭示不同尺度上種群個體分布的均勻程度,拐點顯示這種變化規律發生顯著轉換的尺度。它是計盒維數的推廣,其不但考慮所取格子是“空”還是“非空”,而且考慮了不同的非空格子所提供的信息量大小,從而反映種群空間分布的非均勻性程度。高的信息維數(接近2)表明種群格局強度尺度變化強烈,個體分布不均勻,隨著觀測尺度的改變,種群格局分布類型會出現明顯的變化。信息維數較低,可能是因為個體隨機分布或均勻分布[38]。
紫花針茅與醉馬草的種群直線擬合的相關系數均≥0.968(表5),P<0.01,存在極顯著的冪律關系,說明兩個種群個體分布格局具有高度地自相似性。紫花針茅種群的信息位數區間為[1.590,1.899],極差0.309;醉馬草種群的信息位數區間為[1.633,1.913],極差0.280。紫花針茅種群與醉馬草種群的信息位數均接近2,表明放牧造成兩個種群均勻度變化強烈,種群格局具有較高的空間不均勻性;而紫花針茅的信息位數極差高于醉馬草,說明受放牧影響紫花針茅的均勻度變化更加強烈。
紫花針茅種群的信息維數隨放牧率增加呈下凹趨勢,表明放牧率越大,個體分布均勻性隨尺度的變化越發明顯,變化趨勢可用二次曲線y=-0.015x2+ 0.074x+ 1.814(R2=0.994,P<0.01)擬合。信息維數與計盒維數線性回歸y=1.223x-0.343(R2=0.955,P<0.01),表明伴隨紫花針茅分布越均勻,占據空間越小。
醉馬草種群的信息維數隨放牧率的變化沒有較一致的規律,但與計盒維數極顯著正相關(y=0.421x+ 1.129,R2=0.824,P<0.01)。除放牧率6.90 AUM/hm2外,其他樣地紫花針茅的信息維數均不小于醉馬草,說明放牧對紫花針茅個體分布均勻性影響較大。
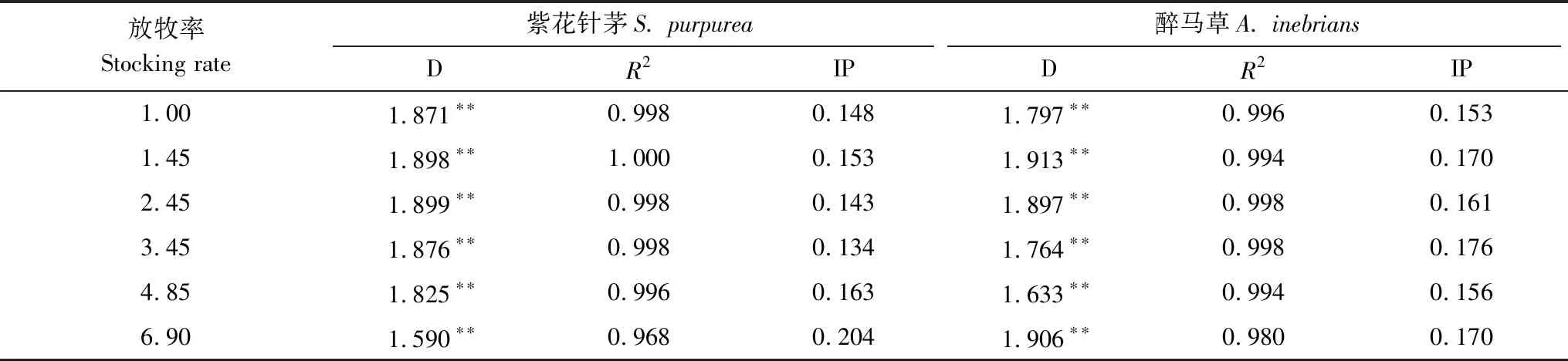
表5 各放牧率下植物種群空間分布格局的信息維數
D表示信息維數;R2表示信息維數的相關系數;IP表示信息維數的拐點尺度;**表示信息維數差異極顯著(P<0.01)
3 討論
本次研究結果表明,隨甘肅馬鹿放牧率的增大,種群的空間占據能力變化,種群均勻度改變,冬季牧場植物種群格局改變[19]。種群格局在大尺度研究時,氣候、土壤、地形等環境因子為主導因子,而局部尺度研究時,放牧成為影響草地群落特征最主要的因子(表1)。
采食與踐踏是牧場種群變化的主要因子[43]。通過偏離系數及t檢驗顯著性分析,了解冬季牧場各種群的分布格局變化。紫花針茅、波伐早熟禾、扁穗冰草通過分蘗與補償生長提高耐牧性;醉馬草因含有生物堿-麥角酰胺和麥角新堿有毒;冷蒿具備生根萌蘗再生能力[41]。伴隨放牧率升高,家畜采食嚴重,冷蒿釋放5類揮發性有機化合物造成扁穗冰草的凱氏帶發育不完全[42]。
紫花針茅種群隨放牧率升高,偏離系數呈凹拋物線趨勢,計盒維數偏低,信息維數均呈凸拋物線趨勢。在重度放牧與輕度放牧下,紫花針茅集群度均高,在6.90 AUM/hm2呈現集群分布;計盒維數卻偏低,占據空間相對較小;信息維數偏小,個體分布越隨機。在輕度放牧時,各植物種群受放牧影響較小,種間競爭相對激烈,紫花針占據空間相對中度放牧較小,個體分布更為均勻,而整體分布集群度偏高。在重度放牧下,紫花針茅生長不良,占據空間很小,整體呈小型“草斑”,偏離系數偏高,局部個體分布偏均勻,信息維數偏低。段敏杰等人在藏北紫花針茅草地放牧發現[19],紫花針茅綜合優勢比在中度放牧最高。中度放牧抑制群落中優勢種而促進其他物種生長[44-45]。本次實驗也顯示中度放牧下紫花針茅計盒維數最大,空間占據能力最強,生長更為優勢。
醉馬草,因生物堿有毒[46],無法被家畜采食。醉馬草生長受放牧調控,有明顯趨勢,但其顯著相關性低于紫花針茅的種間競爭。隨放牧率增大,醉馬草種群偏離系數、計盒維數、信息維數均呈凸拋物線。在輕度放牧下,醉馬草種群在生態系統中占據生長優勢,但相對紫花針茅生長優勢不足;中度重牧下,來自紫花針茅的種間競爭壓力降低,醉馬草種群生長良好,在嚴重重牧下,土壤板結(表1),醉馬草生長困難,計盒維數與偏離系數都降低。劉秀梅實驗表明伴隨放牧強度的增大,土壤含水量逐漸降低,土壤容重逐漸增大,醉馬草顯著減小[47]。
綜合以上分析,在放牧率<3.45 AUM/hm2時,計盒維數方面,紫花針茅與醉馬草均出現極大值,空間占據能力較高;信息維數方面,紫花針茅極高,均勻度極低,偏離系數偏大,集群度偏高,容易形成草斑,導致草場利用不全,牧草資源未合理利用,收益降低。放牧率位于3.45—4.85 AUM/hm2時,醉馬草計盒維數最小,空間占據最低,信息維數相對較低,種群分布相對均勻,格局變化相對緩慢,適合最大限度利用牧場。在放牧率>4.85 AUM/hm2時,紫花針茅種群的計盒維數明顯下滑,占據空間明顯減少,信息維數急降,偏離系數升高,植株矮化,零散分布。因此,預測牧場適宜放牧率4.34 AUM/hm2最佳。
4 結論
伴隨放牧率增大,家畜喜食牧草的種群格局發生急劇變化,可食牧草的集群程度會隨之降低。重度放牧導致毒害草的增生,不可食的草本植物(醉馬草)將會成為退化牧場新的優勢種。采用全要素試驗方法,觀測連續放牧率對高寒草地的調控,可基于當地氣候、土壤等環境因素,提供更為切實提供實試驗成果。本研究結果表明在放牧率4.34 AUM/hm2時,可實現牧草資源的充分利用與生態保護的最佳平衡。目前,本研究對于伴生禾本科牧草的群落演替機理尚不明確及其在生態系統的重要程度尚不確定,有待進一步探究、完善牧場植物群落演替機制。
