茄科植物中HCT基因家族的鑒定及進化和表達分析
2019-12-16 01:42:53李倩蓋江濤白蓓蓓葉秀旭王鵬
江蘇農業科學 2019年19期
關鍵詞:表達
李倩 蓋江濤 白蓓蓓 葉秀旭 王鵬
摘要:莽草酸/奎寧酸羥基肉桂酰轉移酶(shikimic acid/quinic acid hydroxy cinnamyl transferase,簡稱HCT)是辣椒素苯丙烷代謝途徑中關鍵的限速酶,其催化的產物對茄科植物具有重要的生理學意義。為了探清HCT在茄科植物基因組中的狀況,重點考察了其在茄科植物基因組中的分布,并利用生物信息學分析的方法,對HCT基因進行全面的鑒定,共獲得擬南芥、煙草、番茄和辣椒的13條基因序列。其次,構建了HCT在茄科植物中的系統發育樹,并對其進行相關的結構、功能和表達的分析。結果揭示了茄科植物中HCT基因的親緣關系;HCT基因編碼的蛋白結構域高度保守,該保守性體現在植物的結構和功能上。本研究為進一步研究茄科植物中的HCT基因的酶學及生理功能提供了基礎。
關鍵詞:茄科植物;莽草酸/奎寧酸羥基肉桂酰轉移酶;HCT基因;辣椒素;進化;表達
中圖分類號: S641.301文獻標志碼: A
文章編號:1002-1302(2019)19-0065-03
收稿日期:2018-07-03
基金項目:中國熱帶農業科學院基本科研業務專項 (編號:1630032018023)。
作者簡介:李 倩(1994—),女,安徽池州人,碩士研究生,研究方向為植物基因組學。E-mail:a827680831@163.com。
通信作者:王 鵬,博士,副研究員,主要從事植物天然產物生物技術、植物基因組學。E-mail:pwang521@163.com。
辣椒為茄科(Solanaceae)辣椒屬(Capsicum)一年或有限多年生草本植物,是人們常食用的一種風味蔬菜。研究發現,辣椒的果實中含有豐富的維生素、礦物質以及一些微量元素,可以維持人體正常的生理活動,提高身體免疫力,增強人體機能。辣椒素(capsaicin)類物質是辣椒中辣味的物質基礎,早在1876年,辣椒素就由Thresh分離出來并命名,Nelson最先解析了辣椒素的結構,表明它是一種含香草酰胺的生物堿[1],主要的物質是辣椒堿和二氫辣椒堿[2]。辣椒素的生物合成是個復雜的途徑,其間會涉及到多種酶和代謝物的參與,然而本質上,辣椒素主要由苯丙氨酸途徑合成的香草基胺和由支鏈脂肪酸途徑合成的8-甲基-6-癸烯酰經辣椒素合成酶(CS)催化縮合而成[3-4]。研究者在辣椒素的合成途徑中,發掘到了PAL、C4H、HCT、C3H、COMT、KAS、pAMT、ACL和FAT等多個參與辣椒素生物合成的基因[3],查閱相關文獻發現,其中莽草酸/奎寧酸羥基肉桂酰轉移酶(HCT)是在木質素生物合成路徑中發現較晚的一種酶。
目前普遍認同的木質素生物合成途徑大致可以分為3個步驟:(1)莽草酸途徑,由植物光合作用同化產物到芳香氨基酸——苯丙氨酸、絡氨酸和色氨酸的合成過程;(2)苯丙烷途徑,苯丙氨酸到羥基肉桂酸及其輔酶A酯類;(3)木質素合成的特異途徑,羥基肉桂酸輔酶酯類到合成木質素單體及其聚合物的過程 [5-7] 。其中研究的重點大都是放在苯丙烷途徑和木質素合成特異途徑這2個階段。
HCT是控制木質素中的H-單體和G/S-單體相互轉化和聚合的關鍵酶,在苯丙烷3-羥基化步驟上游和下游的苯丙氨酸途徑中發揮著關鍵的雙重調節作用[8-9],其中H-單體主要呈現在禾本科植物中,而S-單體主要在被子植物中存在[6]。Hoffmann等從煙梗提取物中分離純化出一種可以水解羥基肉桂酰基-CoA酯的蛋白質,從它的序列分析結果揭示出該煙草基因屬于一種植物酰基轉移酶的基因家族,其成員也具有各種功能[10]。最佳的酰基供體和酰基轉移酶是香豆酰輔酶A和咖啡酰輔酶A,該酶能夠催化莽草酸和奎酸酯的合成,也催化其逆反應,從而對苯丙烷途徑中的木質素合成有了明確認識。
HCT在茄科植物辣椒的辣椒素生物合成通路中可催化對-香豆酰輔酶A,使其形成C3H底物的同時將C3H的產物咖啡酰莽草酸/奎寧酸進一步催化生成咖啡酰輔酶A。在木質素單體的合成通路中,不同的木質素單體形成的分水嶺是C3H/HCT,其決定了植物體內的碳源流向,是苯丙烷途徑中的限速酶,也是控制木質素H-單體與G/S-單體的一個關鍵點,木質素不同單體間的比例差異對它的含量也會有所影響[8]。木質素自身具有疏水的化學性質,在植物的生長發育過程中,既可以保持植物細胞內的水分,促進植物抵御干旱等不良生態環境,還可以提高植物抗病、抗擊外來干擾,維持正常生長的能力。因此推論,對于植物中HCT基因的研究具有重要的生物學意義和功能。目前,已經從煙草[10]、擬南芥[11]、番茄[12]、桉樹[13]、歐美楊[14]中克隆出來HCT基因,然而,關于HCT基因在其他茄科植物中的克隆和鑒定,國內鮮有相關的系統生物信息學研究報道。
本研究以擬南芥、煙草和番茄中的HCT基因序列作為參考,以茄科植物中的辣椒作為研究對象,通過BLAST序列比對的方法獲得這些植物HCT基因家族的全部序列,利用生物信息學分析方法中的系統發育分析、聚類分析等方法,來預測和分析茄科植物中的HCT基因的相關性質和功能,為后續HCT基因的研究和利用提供理論基礎。
1 材料與方法
1.1 保守結構域鑒定
把HCT基因的蛋白序列作為query訪問,輸入PfamA數據庫開始檢索,獲取相應的結構域信息[15]。
1.2 數據庫搜索
搜索擬南芥、煙草、番茄、辣椒的HCT基因組,各基因組數據從Phytozome中下載得到[16]。通過搜索該蛋白質的模型數據庫,再從結構域進行選擇,獲取HCT家族的序列信息。
1.3 序列比對與進化樹構建
使用probcons比對之前已經搜索到的蛋白質序列,再利用Phy ML構建與之相對應的系統發育樹,之后進行1 000次bootstrap分析[17-18],最后采用Figtree來顯示構建的進化樹結果。
1.4 相對表達量測定
根據番茄HCT基因的CDs序列設計引物,提取番茄樣品的總RNA,反轉成cDNA,篩選引物,之后做熒光定量測定相對表達量。
2 結果與分析
2.1 結構域鑒定
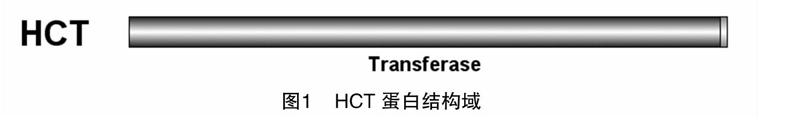
通過與Pfam A數據庫匹配,從已知擬南芥的HCT基因的蛋白質序列中,鑒定得到了1個保守結構域,編號為PF02458.11,顯示該蛋白結構域高度保守,基本占據了整個蛋白質序列的98%,存在很少的非保守序列區域(圖1)。
2.2 含PF02458.11結構域的基因鑒定
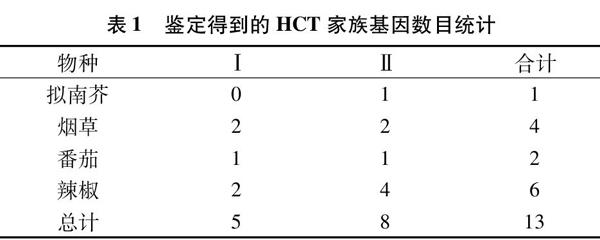
為全面鑒定含有編號為PF02458.11的結構域的基因序列,選取了擬南芥(Arabidopsis thaliana)和茄科植物中煙草(Nicotiana tabacum)、番茄(Solanum lycopersicum)、辣椒(Capsicum annuum)為目標物種,通過BLASTP搜索目標物種基因組的蛋白質模型文件,在BLAST結果中篩選出PF02458.11結構域的序列,最終得到13個HCT家族成員(表1)。
2.3 進化樹構建
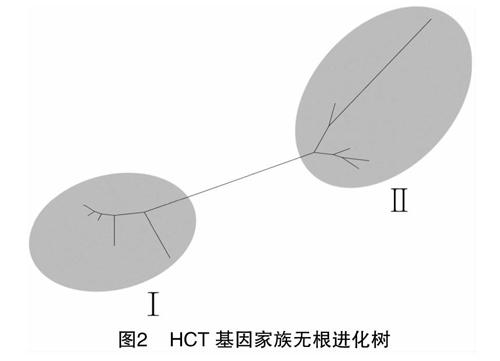
為闡述植物中HCT家族各成員的進化關系,通過最大似然法構建了HCT家族進化樹,結果(圖2)顯示,此家族可分為2條明顯的獨立分支(分別命名為Ⅰ和Ⅱ),共有13條氨基酸序列,其中擬南芥1條、番茄2條、煙草4條、辣椒6條。煙草的2個HCT基因、番茄的1個HCT基因、辣椒的2個HCT基因聚為一支,說明這些基因的親緣關系較近;煙草的2個HCT基因、番茄的1個HCT基因、辣椒的4個HCT基因、擬南芥的1個HCT基因聚為另一支,其中擬南芥單獨分支,說明擬南芥與這些基因的親緣關系較遠(圖3)。
2.4 HCT基因相對表達量分析
通過建立熱圖來進行茄科植物中番茄的HCT基因相對表達量的差異分析。測定結果(圖4)表明,共有野生型和突變型2個基因型,包含3個發育時期:幼果期、轉色期和成熟期,整體來看它們在基因中的表達量較符合生長發育的規律。其中野生型內的Solyc07g043670.1.1、Solyc12g006330.1.1、Solyc09g014280.1.1基因和突變型內的Solyc09g092270.2.1基因在幼果期表達量低,在成熟期表達量高。
3 討論與結論
木質素的生物合成是植物體內碳素次級代謝的一個環節,HCT是木質素合成途徑中研究較晚的一種酶,同時在辣椒素苯丙氨酸合成通路的苯丙烷C3羥基化的代謝途徑中充當關鍵酶和限速酶的重要角色,其代謝產物在植物生長發育的生命活動過程中肩負著不可或缺的作用,這些物質的相對含量也總是與HCT基因的活性保持著關聯性,因而HCT基因對于植物的生命調控具有重大的生物學意義。
本研究對擬南芥、煙草、番茄和辣椒這4種植物的13條HCT基因家族蛋白構建系統發育分析,結果顯示了茄科植物之間HCT基因的親緣進化關系,還鑒定到另一個分支的7條序列與擬南芥的HCT基因分別在2個分支,親緣關系較遠,推測這些基因可能會在茄科植物中發揮重要的生理功能,同時隨著研究的深入,不斷增加的HCT基因完整序列將會構建出更加豐富的系統發育樹。相對表達量的測定結果顯示,本試驗設定的參考對象番茄符合植物生長發育的規律,在幼果期表達量低,成熟期表達量高。
目前,人們對于苯丙烷代謝途徑中的基因研究較多,雖有一些關于植物中HCT基因的研究,但鮮有研究從茄科植物的HCT基因家族層面來分析它的系統進化,包括在辣椒素的相關合成通路中的生長調控影響,還有該基因的表達與辣椒辣味之間的聯系,以及比較茄科植物中HCT基因的性質和功能等等一系列的問題亟待解決。因此本研究以茄科植物為研究對象,對它們的序列結構、性質、進化進行系統分析,為后續研究茄科植物中的HCT基因提供參考依據和理論基礎,促進茄科植物在育種工作中的進一步發展。
參考文獻:
[1]Nelson E K. The constitution of capsaicin—The pungent principle of Capsicum[J]. Journal of the American Chemical Society,1919,188(2):269.
[2]Sarpras M,Gaur R,Sharma V,et al. Comparative analysis of fruit metabolites and pungency candidate genes expression between Bhut Jolokia and other Capsicum species[J]. PLoS One,2016,11(12):e0167791.
[3]成善漢,吳艷閣,賀申魁. 辣椒素生物合成路徑及調控基因研究的最新進展[J]. 分子植物育種,2008,6(2):335-340.
[4]Bennett DJ,Kirby[KG*2]G[KG*2]W. Constitution and biosynthesis of capsaicin[J]. Journal of the Chemical Society C Organic,1968,4:442-446.
[5]王雪霞. HCT基因的克隆及其鑒定[D]. 大連:大連工業大學,2009.
[6]張士花. 不同4CL基因家族成員對植物苯丙烷衍生物代謝調控的研究[D]. 濟南:山東師范大學,2011.
[7]Boerjan W,Ralph J,Baucher M. Lignin biosynthesis[J]. Annual Review of Plant Biology,2003,54(1):519-546.
[8]王雪霞,薛永常,趙文超. 木質素生物合成中C3H/HCT的研究進展[J]. 生命的化學,2008,28(5):650-653.
[9]崔 雪,于月華,申麗婕,等. 海島棉GbHCT基因克隆及生物信息學分析[J]. 生物技術通報,2016,32(4):80-86.
[10]Hoffmann L,Maury S,Martz F,et al. Purification,cloning,and properties of an acyltransferase controlling shikimate and quinate ester intermediates in phenylpropanoid metabolism[J]. The Journal of Biological Chemistry,2003,278(1):95-103.
[11]Niggeweg R,Michael A J,Martin C. Engineering plants with increased levels of the antioxidant chlorogenic acid[J]. Nature Biotechnology,2004,22(6):746-754.
[12]Besseau S,Hoffmann L,Geoffroy P A,et al. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth[J]. Plant Cell,2007,19(1):148-162.
[13]Harakava R. Genes encoding enzymes of the lignin biosynthesis pathway in Eucalyptus[J]. Genetics & Molecular Biology,2005,28(3):601-607.
[14]王雪霞,曹 方,薛永常. 楊樹木質素合成酶hct基因的克隆及核苷酸序列分析[J]. 遼寧林業科技,2009(6):4-7.
[15]Finn R D,Coggill P,Eberhardt R Y,et al. The Pfam protein families database:towards a more sustainable future[J]. Nucleic Acids Research,2016,44(1):D279-D285.
[16]Goodstein D M,Shu S,Howson R,et al. Phytozome:a comparative platform for green plant genomics[J]. Nucleic Acids Research,2012,40:1178-1186.
[17]Do C B,Mahabhashyam M S,Brudno M,et al. ProbCons:probabilistic consistency-based multiple sequence alignment[J]. Genome Research,2005,15(2):330-340.
[18]Guindon S,Delsuc F,Dufayard J F,et al. Estimating maximum likelihood phylogenies with PhyML[J]. Methods in Molecular Biology,2009,537:113-137.
猜你喜歡
甘肅教育(2016年22期)2016-12-20 22:28:05
藝術評鑒(2016年17期)2016-12-19 19:12:28
青年文學家(2016年17期)2016-12-17 20:26:03
中學課程輔導·教師教育(上、下)(2016年17期)2016-12-17 18:11:48
成才之路(2016年35期)2016-12-12 12:08:34
中學課程輔導·教師教育(上、下)(2016年19期)2016-12-07 20:59:07
藝術科技(2016年9期)2016-11-18 18:24:22
藝術科技(2016年9期)2016-11-18 16:48:09
校園英語·上旬(2016年10期)2016-11-16 19:03:00
人間(2016年28期)2016-11-10 00:08:22
