云南省蒙自酸性礦山排水微生物群落結構和功能
2019-12-09 13:02:06梁宗林秦亞玲王沛王保軍劉征華尹華群劉雙江姜成英
生物工程學報 2019年11期
梁宗林,秦亞玲,王沛,王保軍,劉征華,尹華群,劉雙江,3,姜成英,3
研究報告
姜成英 中國科學院微生物研究所研究員。2001年于中國科學院過程工程研究所獲博士學位。2006年6–9月和2008年1月–2009年1月,國家留學基金委選派德國波鴻魯爾大學訪問學者;2018年3–5月,美國東卡羅萊納大學高級訪問學者。長期從事有機污染物的微生物降解和趨化機理及污染環境和特殊環境中微生物種群多樣性和功能多樣性研究;進行極端嗜酸熱古菌和嗜酸細菌鐵、硫代謝調控機制及環境適應機制以及生物冶金技術及礦山污染環境修復治理應用基礎的研究,發現了嗜酸微生物硫代謝新途徑,解析了其環境適應機制。在國內外重要學術期刊上發表相關研究論文50多篇。

云南省蒙自酸性礦山排水微生物群落結構和功能
梁宗林1,2,秦亞玲1,2,王沛1,2,王保軍1,劉征華4,5,尹華群4,5,劉雙江1,2,3,姜成英1,2,3
1中國科學院微生物研究所 微生物資源前期開發國家重點實驗室,北京 100101 2中國科學院大學,北京 100049 3中國科學院環境微生物技術聯合實驗室,北京 100101 4中南大學 資源加工與生物工程學院,湖南 長沙 410083 5中南大學 教育部生物冶金重點實驗室,湖南 長沙 410083
為探究酸性礦山排水生態系統不同環境中的微生物群落和功能,全面了解酸性礦山排水的形成和發展規律,采用高通量測序技術研究云南省蒙自某礦區酸礦水坑和周邊溪水中的原核微生物群落組成,并結合樣本理化特征分析影響群落結構的主要因素,進而解析菌群的環境功能。研究發現酸礦水坑中主要有廣古菌門、變形菌門 (包括α、γ和δ變形菌綱)、硝化螺菌門、厚壁菌門、放線菌門和酸桿菌門等類群,與周邊溪水的群落結構具有明顯差異。群落多樣性與pH呈顯著正相關,而熱原體綱 (Thermoplasmata) 與pH呈負相關,可對群落結構起主導作用。酸礦水坑不同樣本中均具有高豐度的亞鐵原體屬(6.60%–86.34%),酸硫桿菌屬是酸礦水和沉積泥中主要的鐵、硫氧化細菌,而專性鐵氧化的鉤端螺旋菌屬的豐度較低,鐵卵形菌屬幾乎只存在于酸礦水中;此外,嗜酸或耐酸的異養菌廣泛分布于酸礦水和沉積泥中,它們可促進鐵、硫氧化菌的生長及催化礦石溶解。結果表明,pH通過影響微生物多樣性和菌群分布而對酸性礦山排水環境微生物群落結構造成重大影響。
酸性礦山排水,高通量測序,微生物多樣性,群落結構,嗜酸菌
酸性礦山排水 (Acid mine drainage, AMD) 是礦區內常見的一種極端環境,由黃鐵礦 (FeS2) 等硫化礦物在接觸空氣和水的條件下自發氧化及在微生物的參與下加速氧化溶解形成[1],通常呈紅棕色或褐色,也稱為酸性礦山廢水,是礦山廢水中污染范圍最廣且危害最為嚴重的一類[2]。其典型特征是隨著氧化反應的進行,pH逐漸降低,通常為2–4,甚至小于2;具有高濃度的SO42?,以及鐵、鋁、銅、鉛、砷等金屬和類金屬元素。由于廢礦石和尾礦的大量積累,以及舊礦井的廢棄,酸性礦山排水的產生已成為一個全球性的環境問題[3]。
黃鐵礦是地球中含量最為豐富的硫化礦物[4],它的氧化性溶解是酸性礦山排水的主要成因[5],習慣性概括為總反應式:4FeS2+14H2O+15O2→ 4Fe(OH)3+8SO42?+16H+。值得注意的是,黃鐵礦是酸不溶性礦物,其氧化性溶解包含一系列反應過程[1],Fe3+起主要的氧化作用。黃鐵礦與水和氧氣接觸后發生氧化,產生Fe2+,并進一步被氧化為Fe3+;在pH低于4時,Fe2+自發氧化速度慢,它的氧化主要由嗜酸性鐵氧化菌完成[4],且其速度超過自發氧化106倍,使Fe3+快速再生,這是促進硫化礦物氧化的關鍵步驟[6]。另外,硫化礦物氧化還生成還原型無機硫化合物和S0,為硫氧化菌的生長提供能源物質[7]。研究表明,酸性礦山排水自然條件嚴苛且地球化學因素相對簡單,但具有豐富的嗜酸性鐵、硫氧化及異養的細菌和古菌[8-9]。因此酸性礦山排水中的微生物及其鐵、硫元素代謝一直備受關注[10-11]。近些年來,不依賴于培養的微生物群落分析方法如16S rRNA基因克隆文庫、擴增子測序、宏基因組測序以及宏轉錄組測序技術等已廣泛用于酸性礦山排水的研究[12-14],使人們對其中的微生物多樣性、群落結構和功能以及微生物與環境的相互作用有了更全面的認識[15-16]。
酸性礦山排水生態系統包含酸礦水、沉積泥和生物膜等不同環境[13,17],而目前關于其不同環境中微生物群落組成、分布及功能的研究依然較少[17]。本研究采用高通量測序技術分析了云南省蒙自某礦區酸礦水坑不同類型樣本和周邊溪水的原核微生物多樣性及群落結構差異,探究了影響群落結構的主要因素,進而分析了菌群的分布和適應性及重要功能,旨在更全面地了解酸性礦山排水的形成和發展規律及其中的鐵、硫等元素生物地球化學循環[18],發掘極端環境的微生物資源,同時為酸性礦山排水的治理和修復[4]以及微生物浸礦[19]提供科學依據和理論指導。
1 材料與方法
1.1 采樣地點描述和樣本采集
中國云南省蒙自市礦產資源豐富,其中白牛廠銀多金屬礦床已探明銀、鋅儲量達超大型規模,鉛、錫儲量達大型規模[20]。本研究于2017年9月22日采集該礦區內某地點 (東經103°46′47″,北緯23°28′53″,海拔約1 847 m) 自然形成的酸礦水坑中酸礦水 (Drainage)、沉積泥 (Sediment) 和生物膜 (Biofilm) 等不同環境的樣本,并根據pH高低將各酸礦水坑分為B、C、D區。B區采集3個酸礦水樣本 (BD) 和5個沉積泥樣本 (BS),C、D區各采集3個酸礦水樣本 (CD和DD) 和3個沉積泥樣本 (CS和DS),E區采集3個生物膜樣本 (EF)。酸礦水坑周邊一處酸性水流 (Rill) 作為A區,采集3個周邊溪水樣本 (AR)。
1.2 理化參數測定
采用便攜式水質多參數測量儀 (Star A329, Orion, USA) 測定采樣點的溫度、pH和電導率等參數。基于鄰菲啰啉分光光度法 (HJ/T 345-2007) 和比濁法[21],采用便攜式比色計 (DR/890, HACH, USA) 測定酸礦水樣本的Fe2+、Fe3+和SO42?濃度。取適量樣本經微波消解儀 (TOPwave, Analytik Jena, Germany) 消解及電熱板加熱趕酸后稀釋到適宜濃度,采用ICP-OES (Optima 5300 DV, PerkinElmer, USA) 測定元素含量[22]。
1.3 樣本基因組提取
采用DNA提取試劑盒 (PowerMax Soil DNA Isolation Kit, Qiagen) 提取樣本DNA。采用溶劑過濾器和真空泵將酸礦水抽濾通過微孔濾膜 (PES, Φ47–50 mm 0.22 μm, PALL) 以收集樣本中的微生物,用無菌手術剪和鑷子將濾膜剪碎[23]后進行DNA提取。沉積泥和生物膜樣本直接按照試劑盒操作說明進行提取。DNA溶解于Tris鹽酸緩沖液 (pH 8.0)中,?20 ℃凍存,由北京博奧匯玖生物科技有限公司完成16S rRNA基因擴增子測序。
1.4 16S rRNA基因擴增子測序
選擇16S rRNA基因的V4–V5區進行擴增子測序。取DNA樣品與已融合樣本標簽 (Barcodes) 的引物515F (5′-GTGYCAGCMGCCGCGGTAA-3′) 和926R (5′-CCGYCAATTYMTTTRAGTTT-3′) 于預混液 (HiFi HotStart ReadyMix, KAPA) 中配制PCR反應體系。PCR反應條件為:95 ℃預變性 3 min;98 ℃變性20 s,55 ℃退火15 s,72 ℃延伸15 s,共25個循環;72 ℃終延伸1 min。PCR產物經凝膠電泳和回收純化后采用Qubit測定DNA濃度并連接測序接頭完成文庫構建。采用Agilent 2100生物分析儀檢測文庫片段長度分布和濃度,選擇Illumina MiSeq PE250平臺完成 測序。
1.5 高通量測序數據分析
序列數據不含測序接頭,根據樣本標簽進行分樣得到各樣本上、下游單端序列。采用USEARCH v11[24]合并單端序列,并用Cutadapt v2.3[25]去除引物;濾除低質量序列和Singletons,并對高質量序列進行去冗余;按照97%相似性聚類 (Cluster) 形成可操作分類單元 (Operational taxonomic units,OTUs)[26]以及通過去噪 (Denoise) 保留數目大于6的序列獲得擴增子序列變體 (Amplicon sequence variants,ASVs)[27],去除嵌合體,OTUs和ASVs統稱為特征 (Features);最終獲得各特征的代表序列并生成特征表。在QIIME2[28]中基于引物對515F/926R和Silva 132 16S rRNA基因數據庫訓練RDP物種分類器[29],并按照0.7置信度閾值進行物種注釋;根據注釋結果去除線粒體和葉綠體序列并過濾特征表;基于樣本最少序列數對特征表抽平后計算各樣本α多樣性指數和距離矩陣。采用R語言及“vegan”、“ggplot2”、“ComplexHeatmap”等R程序包進行統計分析及數據可視化。
2 結果與分析
2.1 樣本理化特征
周邊溪水與酸礦水坑的物理化學特征差異明顯。各采樣區溫度均約為25 ℃,A區pH為5.07±0.15,在各區中最高;B、C、D區pH逐漸降低且同區酸礦水pH略高于沉積泥 (表1);E區酸礦水坑采集到少量生物膜樣本,pH為1.52±0.08。
酸礦水坑中含量最豐富的元素是Fe,沉積泥中還含有大量的Al、S、K、Mg、Ca、P、As、Cu、Pb等元素。隨著pH的降低,酸礦水中Fe2+、Fe3+和SO42?的濃度,以及各元素 (除Mo和Se) 的含量明顯升高 (表1)。D區酸礦水中總Fe離子濃度高達(65 250±2 198) mg/L,同時含有高濃度的SO42?((134 000±2 000) mg/L) 和有毒的類金屬元素As ((641.7±8.8) mg/L)。如不及時加以控制,持續產生的酸性礦山排水將嚴重污染周圍的水體與土壤。
2.2 酸性礦山排水環境微生物群落多樣性
DD組樣本由于條件極端未提取到DNA,其他7組23個樣本經測序共獲得1 300 684條有效序列,通過聚類和去噪及去除線粒體和葉綠體等特征后共獲得968個OTUs和801個ASVs (表2)。比較發現,B、C、D區各組ASVs數明顯高于OTUs數,表明酸礦水和沉積泥中具有較多序列相似性較高的16S rRNA基因,可能存在較多系統發生關系較近的物種,最近已研究采用去噪形成ASVs的方法分析酸性礦山排水中的微生物群落[30],因此本研究選擇去噪的結果用于后續分析。
各樣本的Alpha多樣性指數見表2。Observed指實際測到的特征數;Evenness為辛普森 (Simpson) 均勻度指數;香農 (Shannon) 指數同時衡量豐富度和均勻度,反映群落多樣性情況;PD表示Faith[31]提出的譜系多樣性 (Phylogenetic diversity)。各樣本覆蓋度 (Coverage) 均高于99%,表明測序深度足夠。AR群落豐富度 (ACE和Chao1) 最高,但均勻度較低;B、C、D區多樣性逐漸降低;EF豐富度較高,但均勻度最低。總體來看,pH更低的采樣區表現為群落多樣性更低,而生物膜具有一定的特殊性。
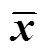
表1 各組樣本的物理化學參數(除AR和EF,±s)
Cond.: conductivity; TDS: total dissolved solids; NA: not available.
2.3 酸性礦山排水環境微生物群落組成
7組樣本共注釋到26個門、45個綱和96個目,通過堆疊柱形圖展示平均相對豐度前20的微生物類群。在門水平 (圖1A) 上,AR中變形菌門 (Proteobacteria) (66.58%) 和擬桿菌門 (Bacteroidetes) (31.41%) 占主要優勢,屬于豐富類群 (平均相對豐度≥1%);其次為芽單胞菌門 (Gemmatimonadetes) (0.92%) 和藍細菌門 (Cyanobacteria) (0.38%),但這兩個門屬于低豐度稀有類群 (平均相對豐度<1%)。BD中變形菌門 (63.98%) 的豐度最高;廣古菌門 (Euryarchaeota) (17.28%)、硝化螺菌門 (Nitrospirae) (11.82%)、酸桿菌門 (Acidobacteria) (4.00%) 和未分類的古菌 (Unclassified Archaea) (1.34%) 成為豐富類群;厚壁菌門 (Firmicutes) (0.61%)、浮霉狀菌門 (Planctomycetes) (0.50%)、放線菌門 (Actinobacteria) (0.35%)、Dependentiae (0.06%) 和Diapherotrites (0.03%) 的豐度增高。BS中廣古菌門 (50.12%) 的豐度明顯更高,在環境中占主導地位 (平均相對豐度≥50%),未分類古菌(14.49%)、硝化螺菌門 (12.52%)、放線菌門 (3.04%)、Diapherotrites (0.74%) 和綠彎菌門 (Chloroflexi) (0.48%) 相比于其他組中豐度最高。與B區相比,CD中廣古菌門 (65.51%) 和Diapherotrites (0.45%) 的豐度更高;隨著酸礦水酸度和離子濃度的升高,CS和DS中廣古菌門的豐度逐漸升高,分別為55.76%和86.40%。生物膜EF中,廣古菌門的豐度高達96.40%,其他門為稀有類群。
在綱和目水平 (圖1B–C) 上,酸礦水坑樣本中熱原體綱 (Thermoplasmata) 熱原體目 (Thermoplasmatales) 豐度最高且分布最廣,其次為γ變形菌綱 (Gammaproteobacteria) 酸硫桿菌目 (Acidithiobacillales) 和未定位的類群;而γ變形菌綱中的β變形菌目 (Betaproteobacteriales) 和未培養的類群,以及α變形菌綱 (Alphaproteobacteria) 和擬桿菌綱 (Bacteroidia) 則主要存在于周邊溪水AR中。硝化螺菌門中只檢測到硝化螺菌綱 (Nitrospira) 的硝化螺菌目 (Nitrospirales),主要存在于BD(11.82%)和BS(12.52%)中。α變形菌綱醋桿菌目 (Acetobacterales) 和酸桿菌綱 (Acidobacteriia) 酸桿菌目 (Acidobacteriales) 主要分布于BD (17.72%和4.00%) 中。梭菌綱 (Clostridia)梭菌目 (Clostridiales) 在酸礦水坑各組中保持較低豐度 (0.29%–1.95%)。酸微菌綱 (Acidimicrobiia) 中未培養的類群和酸微菌目 (Acidimicrobiales) 主要分布于BS (1.82%和1.10%) 中。δ變形菌綱 (Deltaproteobacteria) 的Sva0485[32]和微古菌綱 (Micrarchaeia) 的ARMAN-2主要位于BS (4.30%和0.72%) 和CD (2.26%和0.45%) 中。
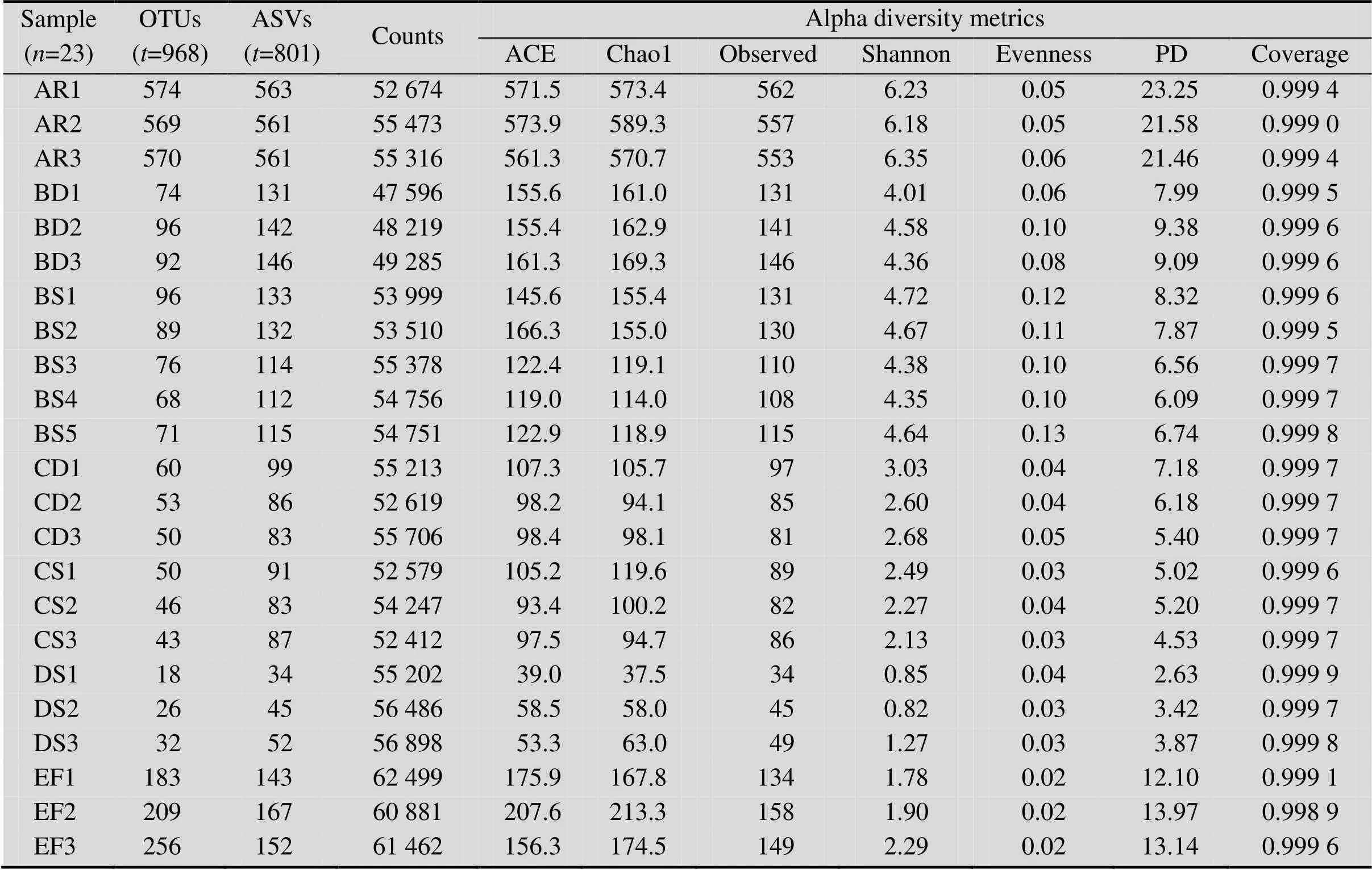
表2 各樣本聚類和去噪的結果及α多樣性指數
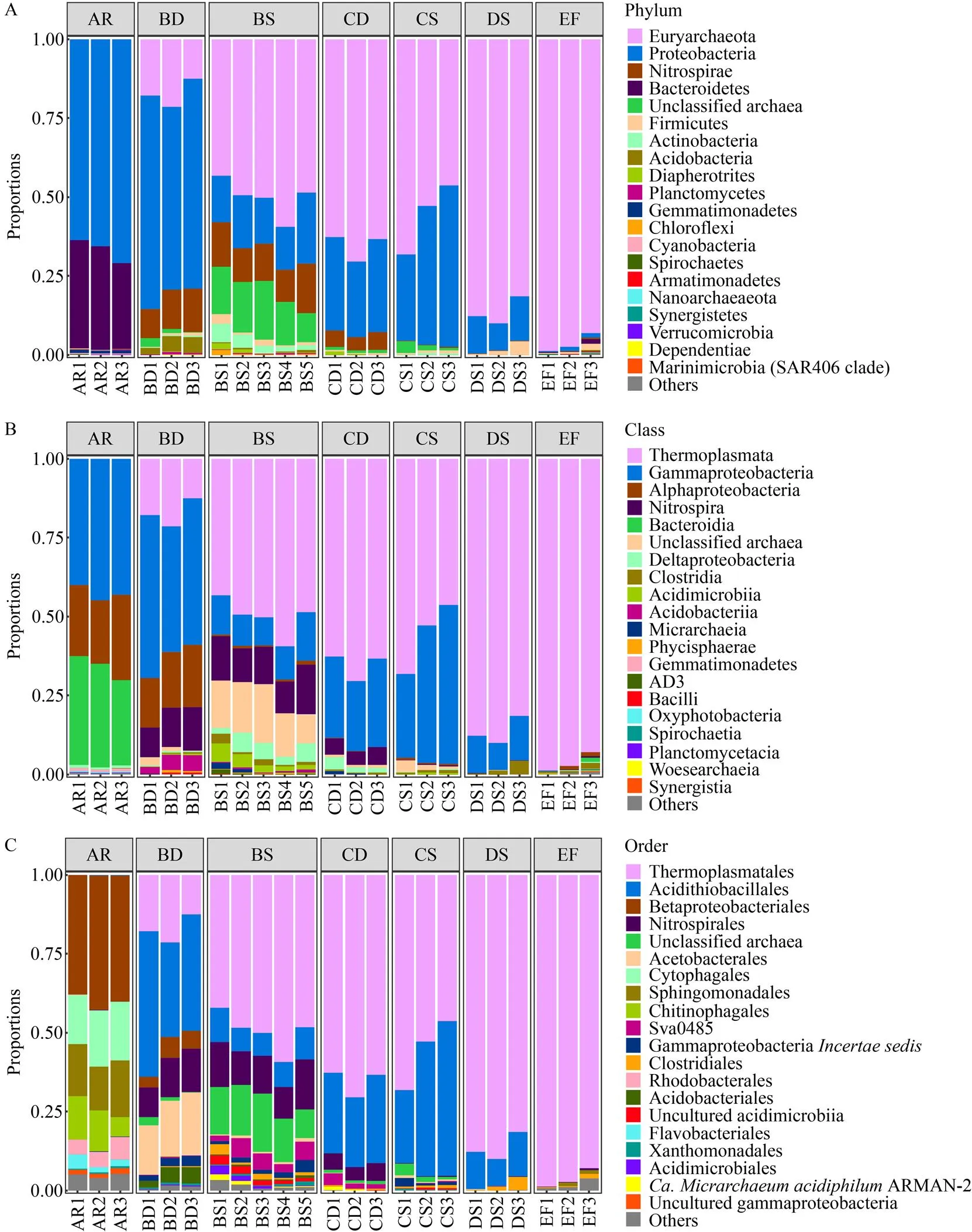
圖1 酸性礦山排水環境微生物類群在不同分類水平上的相對豐度
2.4 酸性礦山排水環境微生物群落結構差異
基于Bray-Curtis相異和Jaccard相似距離矩陣進行主坐標分析 (Principal coordinates analysis, PCoA),結果表明,考慮群落組成豐度 (圖2A) 時,各組群落結構間存在明顯差異;而不考慮豐度(圖2B)時,相同采樣區的酸礦水和沉積泥間差異明顯減小。進一步基于Bray-Curtis距離作PERMANOVA檢驗,結果同樣表明,各組群落結構存在極顯著差異 (2=0.973,=0.001),酸礦水和沉積泥間均具有明顯或顯著差異 (表3)。以上結果說明,各采樣區和樣本類型的群落結構存在明顯差異,同區酸礦水和沉積泥的群落結構差異主要受豐度的影響。層次聚類 (圖3) 直觀地展示各樣本群落結構的相似程度,可見周邊溪水和生物膜明顯區別于其他組;同區酸礦水和沉積泥較為相似;C區與D區樣本更相似。
2.5 酸性礦山排水環境微生物群落對環境變量的響應
對各樣本pH和Shannon指數進行線性回歸分析 (圖4),同時計算皮爾遜 (Pearson) 相關系數。結果顯示,pH和Shannon指數呈極顯著正相關 (=0.804,≤0.001),表明群落多樣性隨pH降低而降低。
進一步以綱水平 (γ變形菌綱以目水平) 群落組成及豐度進行冗余分析 (Redundancy analysis, RDA),紅色劃線表示平均相對豐度 (>0.05%) 前20的微生物類群 (圖5)。RDA1和RDA2軸的解釋率分別為54.55% (≤0.05) 和19.97% (≤0.05),表明群落結構主要趨勢已被pH (≤0.05) 和樣本類型 (≤0.05) 解釋,pH及周邊溪水和生物膜對群落結構具有重大影響。包括α變形菌綱、擬桿菌綱、熱原體綱、梭菌綱、β變形菌目和γ變形菌綱未培養的類群在內的6個微生物類群對第一軸有重要貢獻,熱原體綱只與梭菌綱呈正相關;其余14個類群則對第二軸有主要的貢獻,之間多呈正相關。
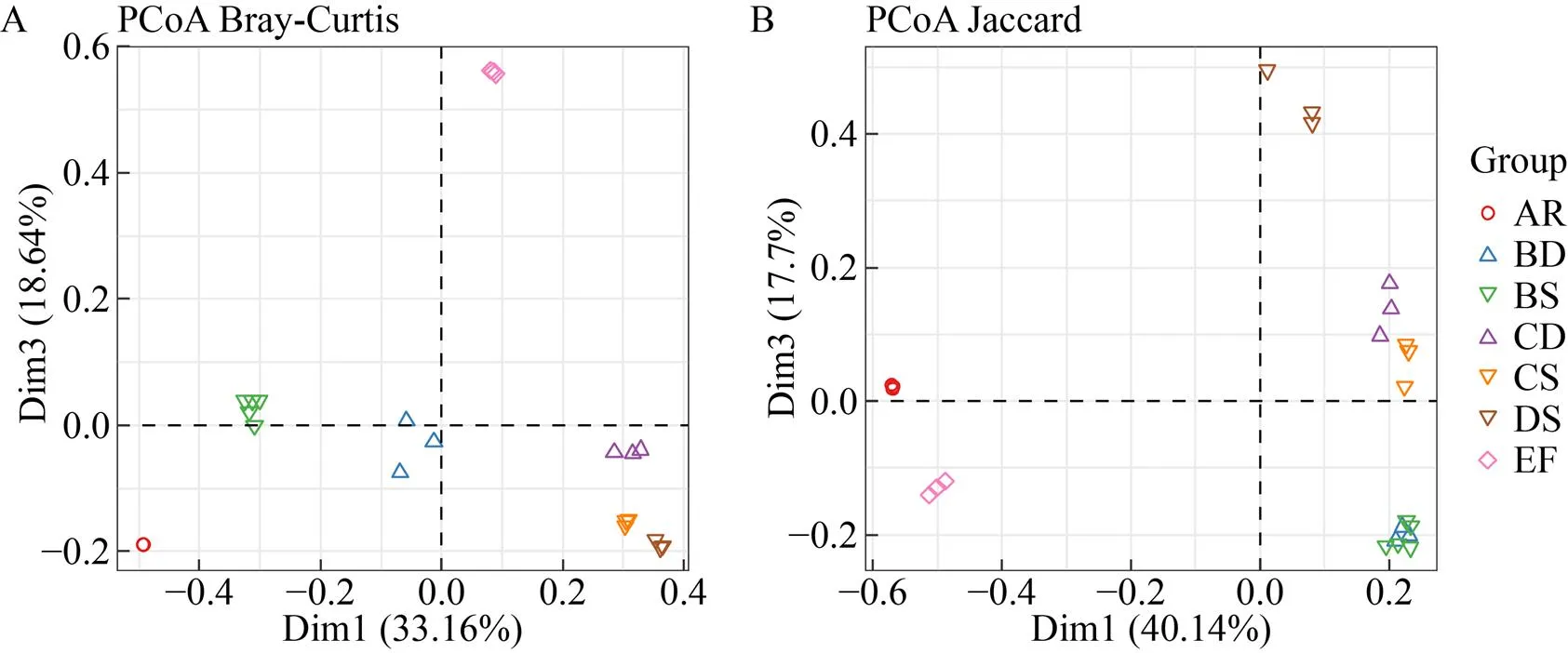
圖2 基于Bray-Curtis相異和Jaccard相似距離矩陣的主坐標分析(PCoA)
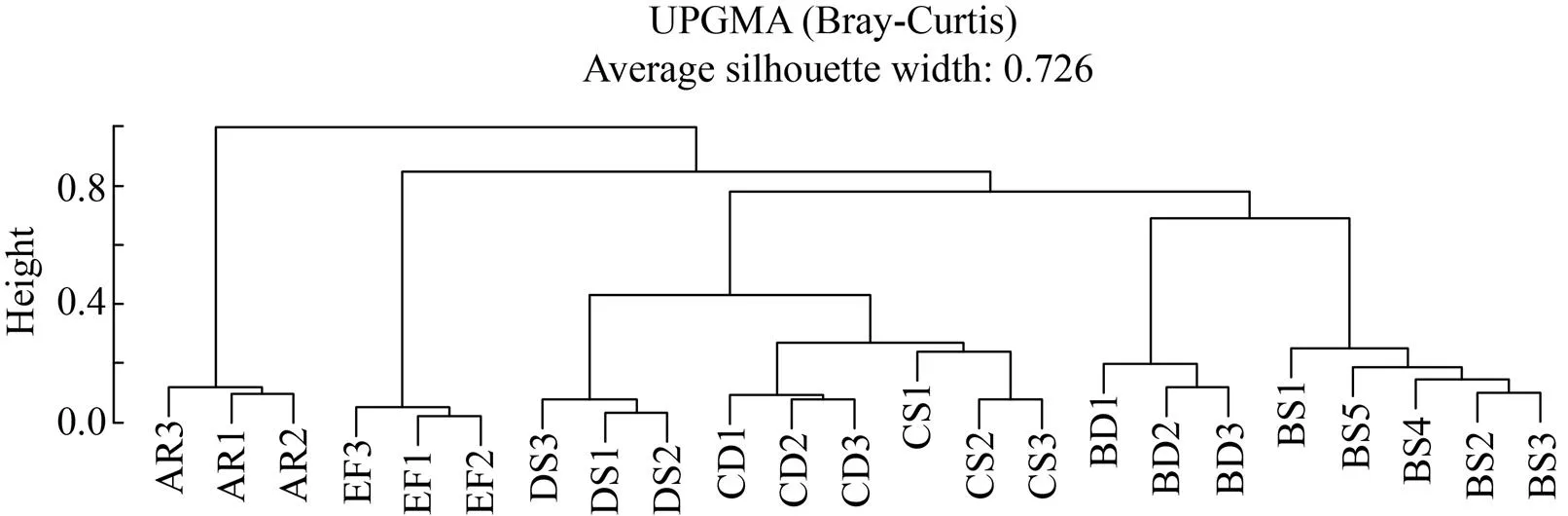
圖3 基于非加權組平均法 (UPGMA) 和Bray-Curtis距離矩陣對各樣本群落結構相似性的層次聚類分析
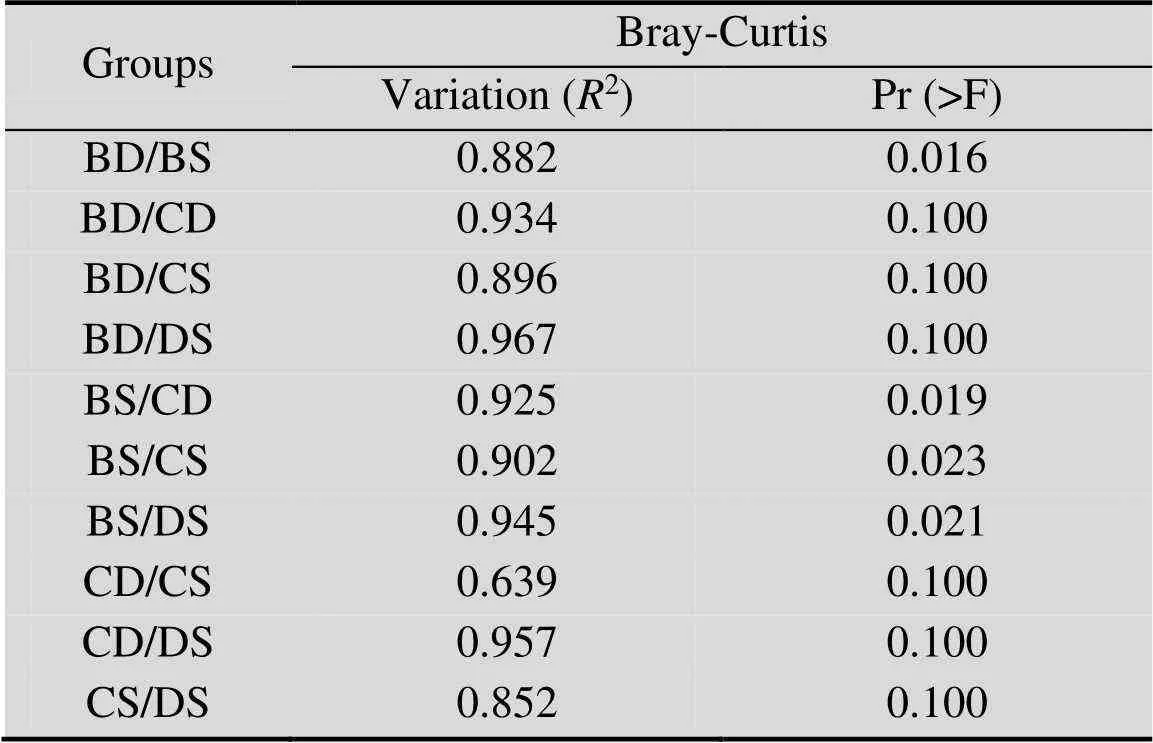
表3 基于Bray-Curtis距離矩陣對各組樣本(除AR和EF)群落結構差異進行置換多元方差分析(PERMANOVA) 檢驗的結果
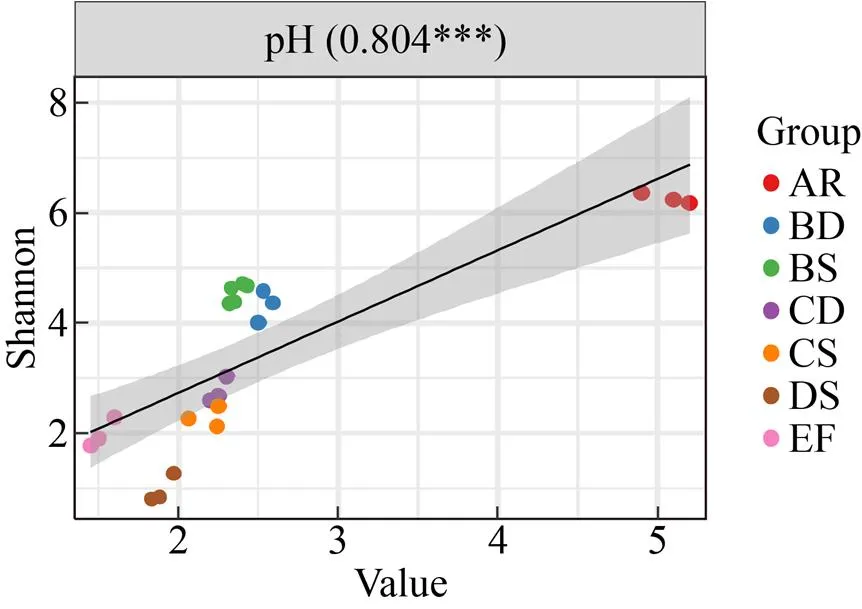
圖4 各樣本pH和Shannon指數的線性回歸分析及Pearson相關系數
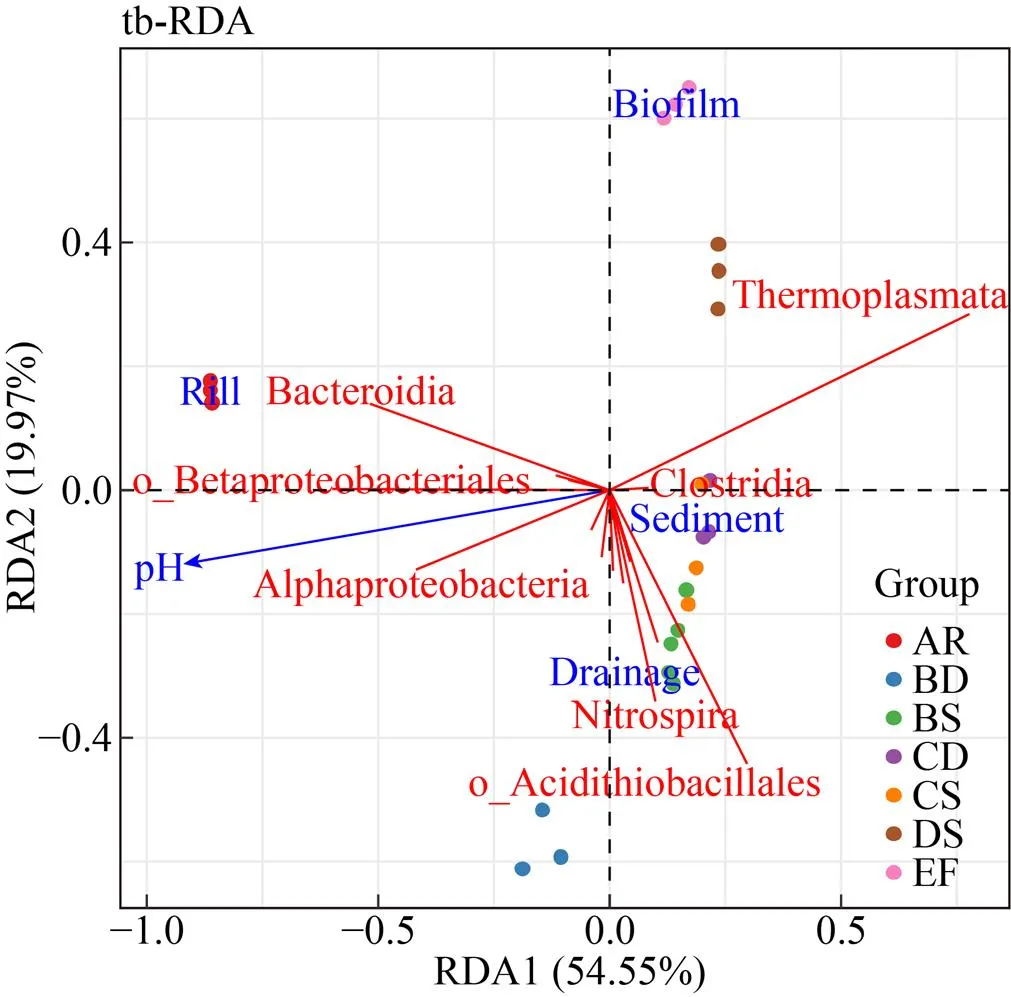
圖5 綱水平微生物類群的冗余分析(RDA)
在屬水平 (圖6) 上,AR中的菌群與其他組具有明顯差異,特有的高豐度類群主要歸為α變形菌綱、β變形菌目和擬桿菌綱,且明顯聚為一簇,其他低豐度的嗜酸性微生物極有可能遷移自酸性礦山排水。
整體來看,酸礦水坑中的菌群組成極為相似,亞鐵原體屬、酸硫桿菌屬、鉤端螺旋菌屬等豐度高且分布廣的類群明顯聚為一簇。亞鐵原體屬 (6.60%–86.34%) 的豐度與酸度呈顯著正相關,在條件極端的DS中達到最高,此時硫化芽胞桿菌屬的豐度也最高 (1.58%)。酸硫桿菌屬在酸礦水和沉積泥 (pH 2.0–2.5) 中的豐度達8.75%–38.29%。鉤端螺旋菌屬主要分布于BD和BS (11.82%和12.52%) 中。此外,BD中分布有最高豐度的酸桿菌屬(2.44%)、醋桿菌科 Acetobacteraceae未培養的類群 (5.90%)、嗜酸菌屬(11.12%)、鐵卵形菌屬(5.26%)和(2.16%) 等,而BS中主要為熱原體科 (Thermoplasmataceae) 的未培養類群 (15.81%)、(21.64%)、(0.63%)、A原體 (A-plasma) (3.17%) 和E原體 (E-plasma) (1.26%)[33]以及鐵線菌屬(1.04%) 等,這些微生物類群多與酸度呈顯著或極顯著負相關。EF中主要為亞鐵原體屬和酸原體屬(79.26%和16.97%),幾乎不存在 (平均相對豐度<0.01%) 或未測到酸硫桿菌屬和。
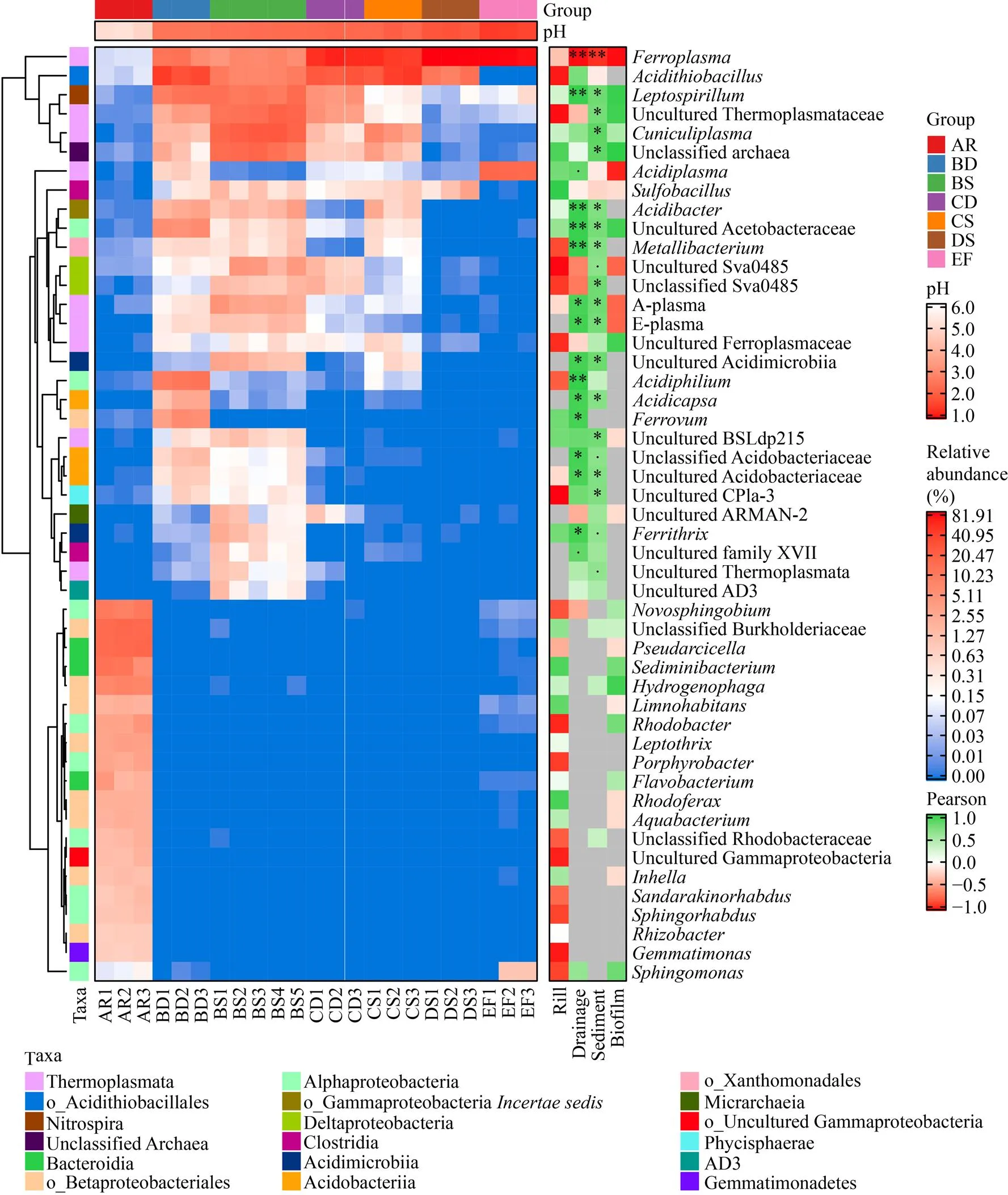
圖6 屬水平上微生物類群的相對豐度及與酸度的Pearson相關性組合熱圖
3 討論
3.1 酸性礦山排水環境菌群的分布及適應性
結合群落結構及菌群與環境因子相關性分析發現,本研究蒙自酸礦水坑不同環境與周邊溪水群落組成差異較大,起主要作用的環境因子為pH。周邊溪水pH最高,群落豐富度也最高,主要有變形菌門β變形菌目和α變形菌綱以及擬桿菌門擬桿菌綱,其特有的高豐度類群在酸礦水坑中難以存活,只有少量存在于生物膜中 (圖6),生物膜中可能形成了特殊的微環境。擬桿菌綱幾乎只分布于周邊溪水中,與pH呈明顯正相關 (圖5)。通常擬桿菌門在酸性礦山排水中不常見,在環境酸度較低或測序深度足夠大的情況下偶有報道[22,34],其僅有的耐酸類群最適pH大于5.0[35]。可認為擬桿菌門不適應酸性礦山排水環境,但與擬桿菌綱呈明顯正相關的β變形菌目和α變形菌綱中具有適應性較好的鐵卵形菌屬和醋桿菌目,這些微生物主要分布于B區酸礦水中。
廣古菌門、變形菌門 (α、γ和δ變形菌綱)、硝化螺菌門、酸桿菌門、放線菌門和厚壁菌門等廣泛分布于蒙自酸礦水坑中,至少在一組中為豐富類群,這些微生物常見于酸性礦山排水環境且表現出很強的適應性[22],此外,Diapherotrites、浮霉狀菌門、綠彎菌門、Dependentiae (TM6)[36]和藍細菌門也存在于酸礦水坑不同環境中,最近已有研究報道這些類群分布于不同的酸性礦山排水中[30,37]。本研究發現熱原體綱熱原體科未培養的類群 (ASV_8, 16, 19, 20) 與A、E原體和在沉積泥中豐度均較高,其分布特征與鉤端螺旋菌屬最為近似。酸桿菌門的主要存在于酸礦水中,而有研究發現該門的類群在沉積泥中豐度也較高[37]。值得關注的是,本研究酸礦水坑中具有高豐度的未分類的古菌 (ASV_7, 18, 25),其與Rio Tinto河酸性礦山排水中未培養的古菌[38]聚為一個OTU,它們在環境中的作用和適應性有待進一步研究。ARMAN-2是該環境中主要的Diapherotrites類群,在沉積泥和酸度更低的酸礦水中均有分布,而在生物膜中幾乎不存在;其他未培養的類群Sva0485和浮霉狀菌門的CPla-3在酸礦水和沉積泥中均有分布,而綠彎菌門的AD3主要分布于沉積泥中,這些類群在酸性礦山排水中較少有報道[13,39]。
研究表明,pH對酸性礦山排水環境微生物群落結構具有重大影響。隨著硫化礦物的氧化性溶解,酸性礦山排水的pH不斷降低,進而溶解更多的金屬和類金屬元素,離子濃度不斷升高,群落結構及菌群分布可能也發生著適應性的變化。本研究發現同區酸礦水和沉積泥的群落結構差異主要表現在群落組成豐度上,隨著pH的降低,酸礦水坑微生物群落多樣性明顯降低,表現為群落組成和結構均趨于簡單,主要原因可能為熱原體綱對該環境的適應性極強,其豐度與pH呈明顯負相關,在極端條件下對群落結構起主導作用,在屬水平上表現為亞鐵原體屬在低pH采樣區中豐度明顯升高。
3.2 酸性礦山排水環境菌群的功能分析
鐵卵形菌屬主要分布于B區酸礦水 (pH 2.54±0.05) 中,在酸度和離子濃度更高的分組中均未測到。該屬成員以Fe2+和黃鐵礦自養生長,對有機物敏感[40],廣泛分布于酸性礦山排水環境,中度嗜酸,在pH 2.0–4.0的條件下[22,41]和一些特殊生 境[42]中可占主要優勢。推測鐵卵形菌屬主要在酸性礦山排水形成初期進行鐵氧化,促進礦石溶解,而隨著環境酸度和離子濃度的升高,其生態地位逐漸被其他鐵氧化菌取代。
酸硫桿菌屬在酸礦水和沉積泥中均有較高的豐度 (8.75%–38.29%),可利用還原型無機硫化合物和S0作為能源自養生長,部分種 (.等) 還可利用Fe2+和黃鐵礦等硫化礦物,以及還原Fe3+[43]。該屬是酸性礦山排水中常見的嗜酸性鐵、硫氧化細菌,最適pH小于3。本研究從酸礦水樣本中分離得到了該屬多株菌,包含潛在新種,可應用于生物浸礦中,促進硫化礦物的氧化。鉤端螺旋菌屬 (0.09%–12.52%) 的豐度僅次于酸硫桿菌屬,spp.是嚴格好氧的嗜酸性鐵氧化細菌,以Fe2+作為唯一能源嚴格自養生長,對有機物極其敏感。相比于酸硫桿菌屬,該屬適應更低的pH,在靠近礦石溶解的區域通過再生Fe3+對硫化礦物的氧化可能發揮著主要的促進作用[44]。
亞鐵原體屬 (6.60%–86.34%) 在酸礦水坑中分布最廣,該屬在極端低pH和高濃度金屬離子的環境中廣泛分布且可占主要優勢[12,45],在生物浸礦體系和酸性礦山排水中相繼得到分離培養,極端嗜酸,好氧且兼性厭氧,能氧化Fe2+及還原Fe3+,同化CO2和有機碳源[46-47]。本課題組的一些研究表明,該屬鐵氧化能力極強。這類不具細胞壁的古菌在酸性礦山排水的形成和發展中發揮著極其重要的作用[48]。
除鐵、硫氧化菌,酸礦水坑中還廣泛分布著酸桿菌屬、嗜酸菌屬、、鐵線菌屬、和硫化芽胞桿菌屬等嗜酸、中度嗜酸或耐酸的異養菌,通過利用環境中的有機物,驅動C元素循環,促進鐵、硫氧化菌的生長。此外,[49]、spp.和[50]可利用還原型無機硫化合物和S0以及Fe2+或硫化礦物兼性化能無機營養或混合營養生長,促進礦石的氧化性溶解;spp.[51]、[52]、[53]、.、酸性礦山排水來源的spp.[54]和spp.中多個成員[55-56]均可在微好氧或厭氧的條件下還原Fe3+,直接參與Fe循環,產生Fe2+供鐵氧化菌利用及催化三價鐵礦的還原性溶解。結合成熟酸性礦山排水中的一些研究[57]及本研究微生物類群相關性分析的結果推測,spp.在極端條件下可能與亞鐵原體屬協同作用以溶解礦石。
4 結論
云南省蒙自某礦區酸性礦山排水環境酸礦水坑和周邊溪水的群落結構存在明顯差異,pH是對其影響最大的環境因素。酸礦水坑群落多樣性較低,但具有豐富的嗜酸性鐵、硫氧化菌,以及代謝能力多樣的嗜酸性異養菌,是鐵、硫元素代謝和生物浸礦等研究和應用的極端菌種資源庫。在嚴苛的環境條件下,酸礦水坑中還蘊含著較多未分類和未培養的微生物類群,它們的生理代謝和環境功能可通過高通量測序數據進行分析,但仍依賴于菌株分離培養來進一步驗證;環境中更為復雜的微生物之間的相互作用及其對群落結構的影響也有待進一步研究。另外,嗜酸菌的遷移及其對環境的影響也需引起足夠的關注。
[1] Johnson DB. Chemical and microbiological characteristics of mineral spoils and drainage waters at abandoned coal and metal mines. Water Air Soil Pollut, 2003, 3(1): 47–66.
[2] Akcil A, Koldas S. Acid mine drainage (AMD): causes, treatment and case studies. J Clean Prod, 2006, 14(12/13): 1139–1145.
[3] Kefeni KK, Msagati TAM, Mamba BB. Acid mine drainage: prevention, treatment options, and resource recovery: a review. J Clean Prod, 2017, 151: 475–493.
[4] Johnson DB, Hallberg KB. Acid mine drainage remediation options: a review. Sci Total Environ, 2005, 338(1/2): 3–14.
[5] Evangelou VP, Zhang YL. A review: pyrite oxidation mechanisms and acid mine drainage prevention. Crit Rev Environ Sci Technol, 1995, 25(2): 141–199.
[6] Singer PC, Stumm W. Acidic mine drainage: the rate-determining step. Science, 1970, 167(3921): 1121–1123.
[7] Schippers A, Sand W. Bacterial leaching of metal sulfides proceeds by two indirect mechanisms via thiosulfate or via polysulfides and sulfur. Appl Environ Microbiol, 1999, 65(1): 319–321.
[8] Baker BJ, Banfield JF. Microbial communities in acid mine drainage. FEMS Microbiol Ecol, 2003, 44(2): 139–152.
[9] Johnson DB, Hallberg KB. The microbiology of acidic mine waters. Res Microbiol, 2003, 154(7): 466–473.
[10] Bonnefoy V, Holmes DS. Genomic insights into microbial iron oxidation and iron uptake strategies in extremely acidic environments. Environ Microbiol, 2012, 14(7): 1597–1611.
[11] Dopson M, Johnson DB. Biodiversity, metabolism and applications of acidophilic sulfur-metabolizing microorganisms. Environ Microbiol, 2012, 14(10): 2620–2631.
[12] Bond PL, Druschel GK, Banfield JF. Comparison of acid mine drainage microbial communities in physically and geochemically distinct ecosystems. Appl Environ Microbiol, 2000, 66(11): 4962–4971.
[13] Méndez-García C, Mesa V, Sprenger RR, et al. Microbial stratification in low pH oxic and suboxic macroscopic growths along an acid mine drainage. ISME J, 2014, 8(6): 1259–1274.
[14] Chen LX, Hu M, Huang LN, et al. Comparative metagenomic and metatranscriptomic analyses of microbial communities in acid mine drainage. ISME J, 2015, 9(7): 1579–1592.
[15] Méndez-García C, Peláez AI, Mesa V, et al. Microbial diversity and metabolic networks in acid mine drainage habitats. Front Microbiol, 2015, 6: 475.
[16] Kuang JL, Huang LN, He ZL, et al. Predicting taxonomic and functional structure of microbial communities in acid mine drainage. ISME J, 2016, 10(6): 1527–1539.
[17] Chen LX, Huang LN, Méndez-García C, et al. Microbial communities, processes and functions in acid mine drainage ecosystems. Curr Opin Biotechnol, 2016, 38: 150–158.
[18] Johnson DB, Hallberg KB. Carbon, iron and sulfur metabolism in acidophilic micro-organisms. Adv Microb Physiol, 2008, 54: 201–255.
[19] Rawlings DE, Johnson DB. The microbiology of biomining: development and optimization of mineral-oxidizing microbial consortia. Microbiology, 2007, 153(2): 315–324.
[20] Li KW, Zhang Q, Wang DP, et al. Isotope geochemistry of Bainiuchang silver polymetallic deposits in Mengzi, Yunnan. Mineral Deposits, 2010, 29(S1): 462–463 (in Chinese). 李開文, 張乾, 王大鵬, 等. 云南蒙自白牛廠銀多金屬礦床同位素地球化學研究. 礦床地質, 2010, 29(S1): 462–463.
[21] Chesnin L, Yien CH. Turbidimetric determination of available sulfates. Soil Sci Soc Am Proc, 1950, 15: 149–151.
[22] Kuang JL, Huang LN, Chen LX, et al. Contemporary environmental variation determines microbial diversity patterns in acid mine drainage. ISME J, 2013, 7(5): 1038–1050.
[23] Fuhrman JA, Comeau DE, Hagstr?m ?, et al. Extraction from natural planktonic microorganisms of DNA suitable for molecular biological studies. Appl Environ Microbiol, 1988, 54(6): 1426–1429.
[24] Edgar RC. Search and clustering orders of magnitude faster than BLAST. Bioinformatics, 2010, 26(19): 2460–2461.
[25] Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J, 2011, 17(1): 3.
[26] Edgar RC. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat Methods, 2013, 10(10): 996–998.
[27] Edgar RC. UNOISE2: improved error-correction for Illumina 16S and ITS amplicon sequencing. BioRxiv, 2016: 081257.
[28] Bolyen E, Rideout JR, Dillon MR, et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol, 2019, 37(8): 852–857.
[29] Wang Q, Garrity GM, Tiedje JM, et al. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol, 2007, 73(16): 5261–5267.
[30] Korzhenkov AA, Toshchakov SV, Bargiela R, et al. Archaea dominate the microbial community in an ecosystem with low-to-moderate temperature and extreme acidity. Microbiome, 2019, 7: 11.
[31] Faith DP. Conservation evaluation and phylogenetic diversity. Biol Conservat, 1992, 61(1): 1–10.
[32] Tan S, Liu J, Fang Y, et al. Insights into ecological role of a new deltaproteobacterial orderAcidulodesulfobacterales by metagenomics and metatranscriptomics. ISME J, 2019, 13(8): 2044–2057.
[33] Yelton AP, Comolli LR, Justice NB, et al. Comparative genomics in acid mine drainage biofilm communities reveals metabolic and structural differentiation of co-occurring archaea. BMC Genomics, 2013, 14: 485.
[34] Cao ZM, Banda JF, Pei LX, et al. Microbial community structure characteristics in different mine drainage lakes of an iron mine in Anhui Province. Acta Microbiol Sin, 2019, 59(6): 1076–1088 (in Chinese). 曹子敏, Banda JF, 裴理鑫, 等. 安徽某鐵礦不同礦山廢水庫中微生物群落結構特征. 微生物學報, 2019, 59(6): 1076–1088.
[35] Sánchez-Andrea I, Sanz JL, Stams AJM.gen. nov., sp. nov., a propionigenic bacterium isolated from sediments of an acid rock drainage pond. Int J Syst Evol Microbiol, 2014, 64(12): 3936–3942.
[36] Yeoh YK, Sekiguchi Y, Parks DH, et al. Comparative genomics of candidate phylum TM6 suggests that parasitism is widespread and ancestral in this lineage. Mol Biol Evol, 2016, 33(4): 915–927.
[37] Mesa V, Gallego JLR, González-Gil R, et al. Bacterial, archaeal, and eukaryotic diversity across distinct microhabitats in an acid mine drainage. Front Microbiol, 2017, 8: 1756.
[38] Amaral-Zettler LA, Zettler ER, Theroux SM, et al. Microbial community structure across the tree of life in the extreme Río Tinto. ISME J, 2011, 5(1): 42–50.
[39] Gavrilov SN, Korzhenkov AA, Kublanov IV, et al. Microbial communities of polymetallic deposits' acidic ecosystems of continental climatic zone with high temperature contrasts. Front Microbiol, 2019, 10: 1573.
[40] Johnson DB, Hallberg KB, Hedrich S. Uncovering a microbial enigma: isolation and characterization of the streamer-generating, iron-oxidizing, acidophilic bacterium “”. Appl Environ Microbiol, 2014, 80(2): 672–680.
[41] Hua ZS, Han YJ, Chen LX, et al. Ecological roles of dominant and rare prokaryotes in acid mine drainage revealed by metagenomics and metatranscriptomics. ISME J, 2015, 9(6): 1280–1294.
[42] Hallberg KB, Coupland K, Kimura S, et al. Macroscopic streamer growths in acidic, metal-rich mine waters in north wales consist of novel and remarkably simple bacterial communities. Appl Environ Microbiol, 2006, 72(3): 2022–2030.
[43] Falagán C, Moya-Beltrán A, Castro M, et al.sp. nov.: an extremely acidophilic sulfur-oxidizing chemolithotroph isolated from a neutral pH environment. Int J Syst Evol Microbiol, 2019, 69(9): 2907–2913.
[44] Schrenk MO, Edwards KJ, Goodman RM, et al. Distribution ofand: implications for generation of acid mine drainage. Science, 1998, 279(5356): 1519–1522.
[45] Golyshina OV, Timmis KN.and relatives, recently discovered cell wall-lacking archaea making a living in extremely acid, heavy metal-rich environments. Environ Microbiol, 2005, 7(9): 1277–1288.
[46] Golyshina OV, Pivovarova TA, Karavaiko GI, et al.gen. nov., sp. nov., an acidophilic, autotrophic, ferrous-iron-oxidizing, cell-wall-lacking, mesophilic member of the Ferroplasmaceae fam. nov., comprising a distinct lineage of the Archaea. Int J Syst Evol Microbiol, 2000, 50(3): 997–1006.
[47] Dopson M, Baker-Austin C, Hind A, et al. Characterization ofisolates andsp. nov., extreme acidophiles from acid mine drainage and industrial bioleaching environments. Appl Environ Microbiol, 2004, 70(4): 2079–2088.
[48] Edwards KJ, Bond PL, Gihring TM, et al. An archaeal iron-oxidizing extreme acidophile important in acid mine drainage. Science, 2000, 287(5459): 1796–1799.
[49] Hiraishi A, Nagashima KV, Matsuura K, et al. Phylogeny and photosynthetic features ofand related acidophilic bacteria: its transfer to the genusascomb. nov. Int J Syst Bacteriol, 1998, 48(4): 1389–1398.
[50] Johnson DB, Bacelar-Nicolau P, Okibe N, et al.gen. nov., sp. nov. andgen. nov., sp. nov.: heterotrophic, iron-oxidizing, extremely acidophilic actinobacteria. Int J Syst Evol Microbiol, 2009, 59(5): 1082–1089.
[51] Falagán C, Johnson DB.gen. nov., sp. nov.: an acidophilic ferric iron-reducing gammaproteobacterium. Extremophiles, 2014, 18(6): 1067–1073.
[52] Küsel K, Dorsch T, Acker G, et al. Microbial reduction of Fe(III) in acidic sediments: Isolation ofJF-5 capable of coupling the reduction of Fe(III) to the oxidation of glucose. Appl Environ Microbiol, 1999, 65(8): 3633–3640.
[53] Ziegler S, Waidner B, Itoh T, et al.gen. nov., sp. nov., an alkalinizing gammaproteobacterium isolated from an acidic biofilm. Int J Syst Evol Microbiol, 2013, 63(4): 1499–1504.
[54] Falagan C, Foesel B, Johnson B.sp. nov.,sp. nov., andsp. nov.: novel acidobacteria isolated from metal-rich acidic waters. Extremophiles, 2017, 21(3): 459–469.
[55] Bridge TAM, Johnson DB. Reduction of soluble iron and reductive dissolution of ferric iron-containing minerals by moderately thermophilic iron-oxidizing bacteria. Appl Environ Microbiol, 1998, 64(6): 2181–2186.
[56] Johnson DB, Joulian C, d’Hugues P, et al.sp. nov., an acidophilic facultative anaerobic Firmicute isolated from mineral bioleaching operations. Extremophiles, 2008, 12(6): 789–798.
[57] Druschel GK, Baker BJ, Gihring TM, et al. Acid mine drainage biogeochemistry at Iron Mountain, California. Geochem Trans, 2004, 5: 13.
Microbial community structure and function in acid mine drainage from Mengzi, Yunnan Province
Zonglin Liang1,2, Yaling Qin1,2, Pei Wang1,2, Baojun Wang1,Zhenghua Liu4,5, Huaqun Yin4,5, Shuang-Jiang Liu1,2,3, and Cheng-Ying Jiang1,2,3
1 State Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China 2 University of Chinese Academy of Sciences, Beijing 100049, China 3 RCEES-IMCAS-UCAS Joint Laboratory for Environmental Microbial Technology, Chinese Academy of Sciences, Beijing 100101, China 4 School of Minerals Processing and Bioengineering, Central South University, Changsha 410083, Hunan, China 5 Key Laboratory of Biometallurgy of Ministry of Education, Central South University, Changsha 410083, Hunan, China
In order to explore the microbial communities and functions in distinct environments of acid mine drainage (AMD) ecosystem and fully comprehend the patterns of AMD formation and development, high-throughput sequencing technology was used to study the prokaryotic community composition in AMD puddles and surrounding rills in a mining area in Mengzi, Yunnan Province, China. By combining with physicochemical properties of the samples, we distinguished the key factors affecting the community structure and analyzed the environmental functions of the microflora. We discovered that the main phyla in AMD puddles were Euryarchaeota, Proteobacteria (including the class α-, γ- and δ-Proteobacteria), Nitrospirae, Firmicutes, Actinobacteria, and Acidobacteria. Community structure of the AMD puddles apparently differed from that of the surrounding rills. Microbial diversity was significantly positively correlated with pH, whereas the relative abundance of Thermoplasmata was negatively correlated with pH and this class might play a predominant role in community structure. There was high relative abundance of the genus(6.60%–86.34%) in different samples of AMD puddles.spp. were the major iron- and/or sulfur-oxidizing bacteria in AMD solutions and sediments, whereas relative abundance of the obligate iron-oxidizerspp. was lower, andspp. were almost only present in the AMD solutions. In addition, acidophilic or acid-tolerant heterotrophic bacteria were widely distributed in the AMD solutions and sediments, which might promote the growth of iron- and/or sulfur-oxidizers and catalyze the oxidative/reductive dissolution of metal ores. Our results suggested that pH significantly impacted the microbial community structure of AMD environment by affecting the microbial diversity and microflora distribution.
acid mine drainage, high-throughput sequencing, microbial diversity, community structure, acidophile
November 12, 2019;
November 15, 2019
The Major Research Plan of National Natural Science Foundation of China (No. 91851206), National Natural Science Foundation of China (No. 31670124), National Basic Research Program of China (973 Program) (No. 2014CB846000), National Material Environmental Corrosion Platform of China.
Shuang-Jiang Liu. E-mail: liusj@im.ac.cn Cheng-Ying Jiang. Tel: +86-10-64807581; E-mail: jiangcy@im.ac.cn
梁宗林, 秦亞玲, 王沛, 等. 云南省蒙自酸性礦山排水微生物群落結構和功能. 生物工程學報, 2019, 35(11): 2035–2049.
Liang ZL, Qin YL, Wang P, et al. Microbial community structure and function in acid mine drainage from Mengzi, Yunnan Province. Chin J Biotech, 2019, 35(11): 2035–2049.
國家自然科學基金重點研發計劃 (No. 91851206),國家自然科學基金 (No. 31670124),國家重點基礎研究發展計劃(973計劃) (No. 2014CB846000),國家材料環境腐蝕平臺資助。
(本文責編 郝麗芳)
猜你喜歡
現代礦業(2021年12期)2022-01-17 07:30:32
中老年保健(2021年12期)2021-08-24 03:30:40
河北地質(2021年2期)2021-08-21 02:43:50
神劍(2021年3期)2021-08-14 02:30:08
昆鋼科技(2021年2期)2021-07-22 07:47:06
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
礦產勘查(2020年7期)2020-12-25 02:43:42
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
