歐洲溫室型黃瓜同源四倍體新種質(zhì)的創(chuàng)制與鑒定
2019-11-28 10:54:11武婭歌趙建華宋曉飛李曉麗孫成振閆立英鄭金雙
江蘇農(nóng)業(yè)科學 2019年18期
武婭歌 趙建華 宋曉飛 李曉麗 孫成振 閆立英 鄭金雙

摘要:為構建黃瓜同源四倍體誘導技術體系,創(chuàng)制新種質(zhì),以3個秋水仙素濃度(0.1、0.2、0.3 g/L)3個處理次數(shù)(3、5、7次)對二倍體歐洲溫室型黃瓜(2n=2x=14)幼苗生長點進行處理,通過植株形態(tài)鑒定與氣孔鑒定、DNA相對含量鑒定和細胞學鑒定等方法,篩選誘變植株并鑒定倍性。研究發(fā)現(xiàn),以2、3 g/L秋水仙素處理5次誘導效果較好,獲得了歐洲溫室型黃瓜同源四倍體(2n=4x=28)和非整倍體(2n=4x=26)。與二倍體親本相比,同源四倍體黃瓜子葉大小、葉片大小、主莖粗、雌雄花大小、保衛(wèi)細胞大小和保衛(wèi)細胞內(nèi)葉單位面積綠體數(shù)目等均呈極顯著增加,但氣孔密度極顯著減少。本研究獲得了歐洲溫室型黃瓜同源多倍體新種質(zhì),為黃瓜種質(zhì)創(chuàng)新與歐洲溫室型新品種選育奠定了基礎。
關鍵詞:歐洲溫室型黃瓜;秋水仙素;多倍體;二倍體;同源四倍體;氣孔;細胞學;誘導效果;形態(tài)特征
中圖分類號:S642.202.4文獻標志碼: A
文章編號:1002-1302(2019)18-0141-05
收稿日期:2018-06-08
基金項目:河北省自然科學基金(編號:C2015407058)。
作者簡介:武婭歌(1993—),女,河北邢臺人,碩士研究生,研究方向為蔬菜作物育種。E-mail:969361514@qq.com。
通信作者:鄭金雙,博士,講師,研究方向為蔬菜作物、藥用植物栽培和育種。E-mail:jinshuangk@163.com。
黃瓜(Cucumissativus L.)為葫蘆科甜瓜屬植物,原產(chǎn)于喜馬拉雅山南麓的熱帶雨林地區(qū)和中國的云南地區(qū),是世界上主要的蔬菜作物之一,也是設施栽培的主要蔬菜作物[1]。不同區(qū)域喜好不同,主栽品種和類型也不同,果實形態(tài)、口感差異較大。歐洲溫室型黃瓜以無刺、色深綠、肉質(zhì)軟、味甜等特點,成為重要的鮮食“水果型”黃瓜。目前,黃瓜遺傳基礎狹窄,我國黃瓜育種材料的親緣關系過于單一,遺傳育種研究相對落后[2]。
多倍體因其“巨大性”,能顯著提高作物的產(chǎn)量和品質(zhì),增強植物的抗病、抗逆能力[3-4]。多倍體育種已廣泛應用在糧食作物[5]、果蔬作物[6-8]、觀賞植物[9-11]以及藥用植物[12-14]的育種工作中。
目前,黃瓜多倍體育種方面的研究已經(jīng)開展。陳勁楓等利用秋水仙素溶液處理萌動種子,獲得了華北型黃瓜同源四倍體[6]。刁衛(wèi)平等采用離體誘導未授粉子房的方法,獲得了華北型黃瓜同源四倍體[15]。鄢郁霖等利用秋水仙素處理干種子和已露出0.5~1.0 cm胚根的萌動種子,獲得了露地型黃瓜同源四倍體[16]。劉永月等利用秋水仙素和二甲基亞砜混合液處理萌動種子,獲得了華北型無毛黃瓜同源四倍體[17]。但關于歐洲溫室型黃瓜多倍體的研究報道較少。本研究采用不同濃度和不同處理次數(shù)對歐洲溫室型黃瓜幼苗生長點進行誘導處理,旨在建立秋水仙素誘導黃瓜幼苗生長點技術體系及其倍性鑒定體系,創(chuàng)造同源四倍體新種質(zhì),豐富黃瓜種質(zhì)資源,為歐洲溫室型黃瓜新品種的選育奠定基礎。
1?材料與方法
1.1?試驗材料
以南京農(nóng)業(yè)大學黃瓜課題組陳勁楓教授提供的歐洲溫室型黃瓜‘南水3號種子為試驗材料。
1.2?多倍體誘導
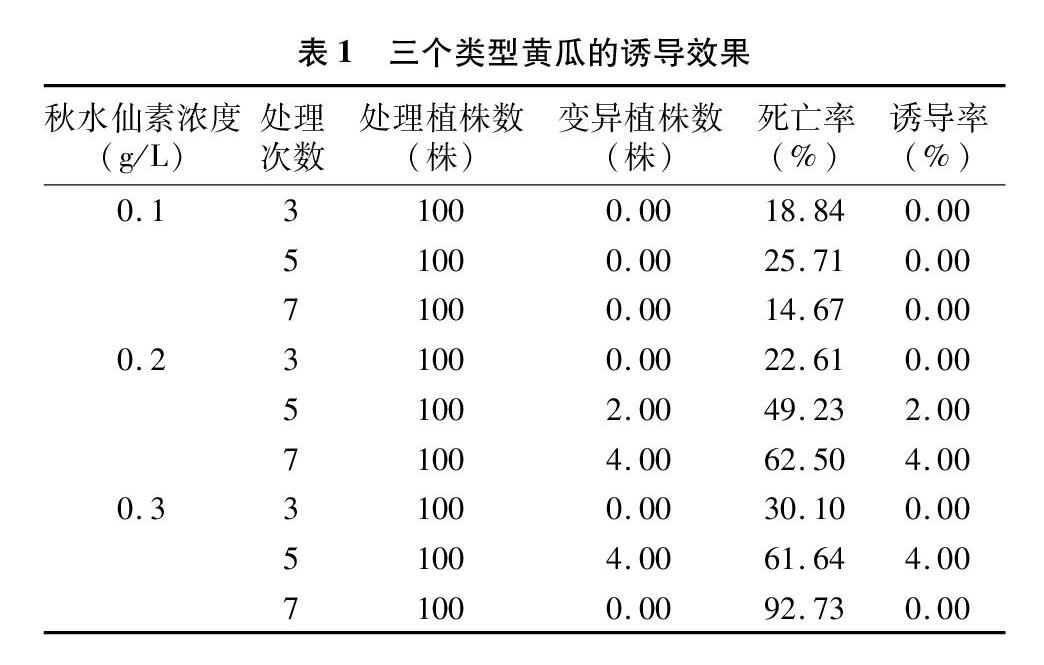
2015年秋,種子溫燙浸種后播于50孔穴盤中,置于日光溫室內(nèi)育苗。2張子葉完全展開后,將棉球置于生長點,用膠頭滴管吸取秋水仙素溶液,滴于生長點棉球上,完全浸濕棉球。于07:00—08:00和17:00—18:00進行0.1、0.2、0.3 g/L 3個秋水仙素濃度處理,3個處理次數(shù)為3、5、7次,共9個處理,以蒸餾水處理為對照,每個處理100株。處理結(jié)束后,棄去棉球。
1.3?多倍體植株的鑒定
1.3.1?植株形態(tài)鑒定與氣孔鑒定
形態(tài)鑒定:于同一時間觀測對照與處理植株的高度、葉片形態(tài),進行變異植株初步的形態(tài)鑒定。
氣孔鑒定:取完全展開的葉片,撕取下表皮,利用顯微鏡觀察氣孔的大小、氣孔密度、保衛(wèi)細胞內(nèi)葉綠體數(shù)量,進行氣孔鑒定。每個植株選取10個視野,統(tǒng)計(20倍視野下)氣孔密度,每個視野隨機選1個氣孔測量氣孔保衛(wèi)細胞的長度和寬度。
1.3.2?DNA相對含量鑒定和細胞學鑒定
通過形態(tài)鑒定和氣孔鑒定,對篩選出的候選變異植株進行自交留種。2016年春播種,對T1代植株進行DNA相對含量鑒定和細胞學鑒定。
DNA相對含量鑒定:取對照和候選變異植株生長點幼嫩葉片,利用流式細胞儀進行DNA相對含量鑒定。
細胞學鑒定:根據(jù)流式細胞儀檢測結(jié)果,對DNA相對含量變化的植株進行染色體計數(shù),進一步鑒定倍性。于晴天07:00—08:00取長度約1 mm的幼嫩雄花,于卡諾固定液(無水乙醇 ∶冰乙酸3 ∶1)中固定24 h,蒸餾水洗滌3次,75%乙醇中保存。采用常規(guī)壓片制片法[18],利用光學顯微鏡觀察花藥絨氈層體細胞染色體數(shù)量,進行細胞學鑒定。每個植株統(tǒng)計20個細胞,以加倍染色體數(shù)量所占比例高于85%作為同源四倍體純合體的鑒定標準。
根據(jù)秋水仙素處理后成苗株數(shù)計算死亡率,根據(jù)上述鑒定方法鑒定獲得的同源四倍體株數(shù)計算誘導率。
死亡率=死亡的幼苗數(shù)/總數(shù)×100%;
誘導率=產(chǎn)生變異的幼苗數(shù)/總數(shù)×100%。
1.4?二倍體與同源四倍體歐洲溫室型黃瓜比較
對鑒定為同源四倍體的歐洲溫室型黃瓜植株進行自交留種,2017年春對T3代植株進行二倍體與同源四倍體植株形態(tài)、顯微特征與果實品質(zhì)比較。
1.4.1?二倍體與同源四倍體歐洲溫室型黃瓜植株形態(tài)特征比較
營養(yǎng)器官:于子葉期和結(jié)果期分別測量二倍體與變異植株的形態(tài)特征,子葉期測定子葉長、子葉寬、子葉節(jié)長、子葉節(jié)粗,結(jié)果期測定植株的高度、中部的葉片長與葉片寬、主蔓節(jié)間長、主蔓節(jié)間粗、葉柄長、葉柄粗。
生殖器官:取當天開放的雌花和雄花進行拍照,分割雌花與子房,比較二倍體和同源四倍體雌雄花和子房大小。
1.4.2?二倍體與同源四倍體歐洲溫室型黃瓜顯微特征比較
氣孔特征比較采用的方法同“1.3.1”氣孔鑒定。取0.5 mm 左右大小的雄花,用卡諾固定液(無水乙醇 ∶冰乙酸=3 ∶1)固定24 h,75%乙醇保存。采用常規(guī)壓片制片法,利用光學顯微鏡觀察花粉粒形態(tài)。取當天開放雄花,利用KYKY-2800掃描電子顯微鏡觀察二倍體與同源四倍體的花粉粒形態(tài)。
1.4.3?二倍體與同源四倍體歐洲溫室型黃瓜果實特征比較
取二倍體與同源四倍體商品瓜,測定商品瓜果實形態(tài)特征(瓜長、瓜橫徑、瓜把長、中心腔、果肉厚)、含水量和含糖量。
1.5?數(shù)據(jù)分析
數(shù)據(jù)統(tǒng)計采用Excle 2010,數(shù)據(jù)分析采用DPS 14.50。
2?結(jié)果與分析
2.1?秋水仙素對歐洲溫室型黃瓜的誘導效果
由表1可知,隨著秋水仙素處理濃度的增加和次數(shù)的延長,死亡率呈增加趨勢。0.1 g/L秋水仙素處理時,黃瓜幼苗死亡率相對最低,變異率為0.00。0.3 g/L 秋水仙素處理時,隨著處理次數(shù)的增加,對生長點毒害作用增強,黃瓜幼苗死亡率相對較高,處理7次時死亡率達到92.73%。綜合誘導率和死亡率,以0.2、0.3 g/L秋水仙素處理5次誘導效果較佳。
2.2?多倍體黃瓜植株的獲得
2.2.1?植株形態(tài)鑒定與氣孔鑒定
根據(jù)植株形態(tài)進行變異植株初步篩選,結(jié)果表明,與對照相比,各處理組黃瓜植株高度、生長速度、葉片大小和形態(tài)均出現(xiàn)不同程度的變異現(xiàn)象,如植株矮小、葉色變深、葉片褶皺等(圖1-A和1-B)。
氣孔的巨大性是多倍體植株在解剖水平的重要表現(xiàn),也是初步篩選和鑒定多倍體的重要標準。篩選出氣孔面積明顯增大、氣孔密度減少的候選變異植株(圖1-C和1-D),并對其進行進一步鑒定。
2.2.2?DNA相對含量鑒定和細胞學鑒定
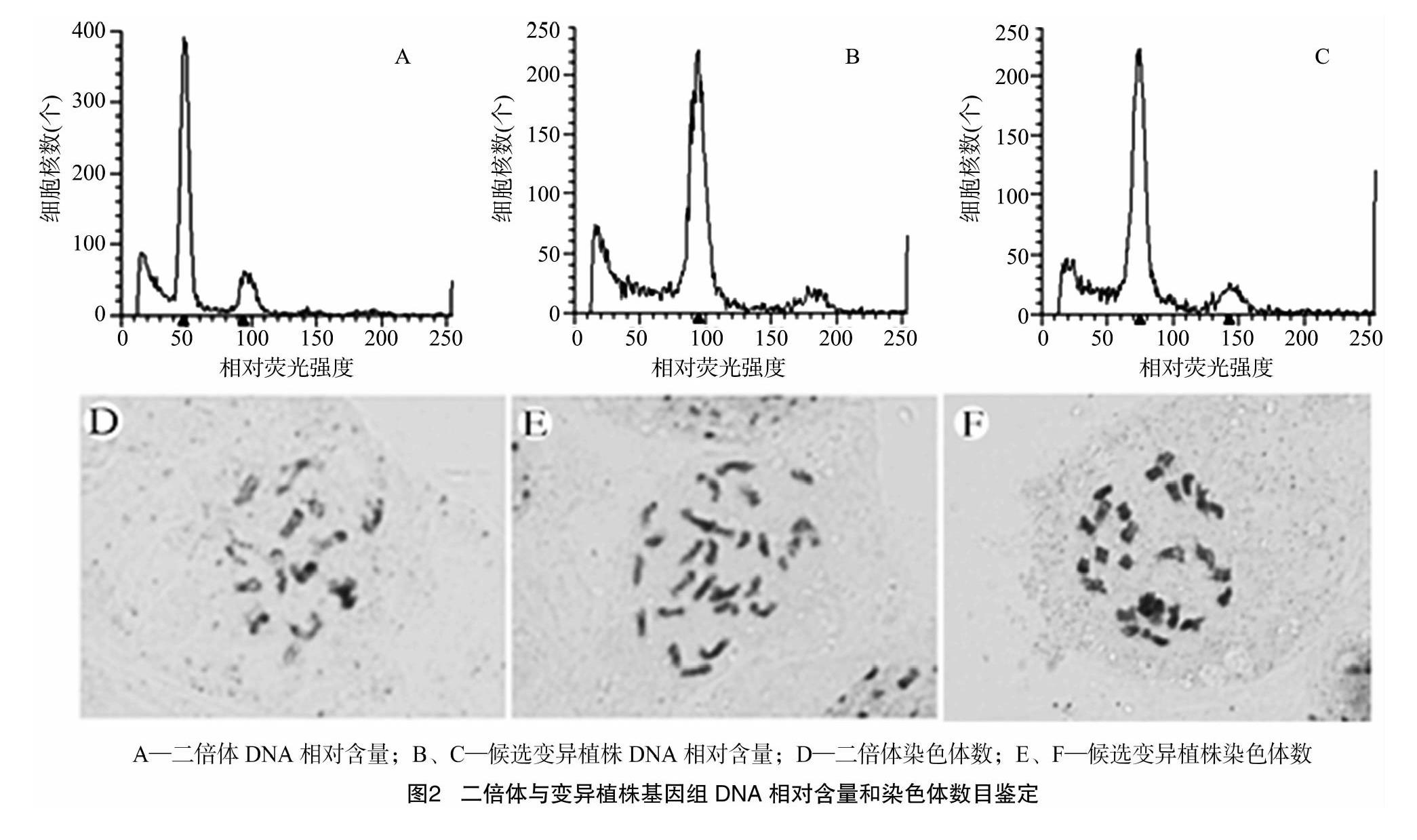
利用流式細胞儀對候選變異植株進行進一步倍性確定。研究發(fā)現(xiàn),二倍體歐洲溫室型黃瓜相對熒光強度為50(圖2-A),候選變異植株的相對熒光強度為100(圖2-B)、75(圖2-C)等。
采用常規(guī)染色體壓片、染色體計數(shù)技術,對變異植株染色體數(shù)目進行進一步確定。結(jié)果表明,二倍體黃瓜染色體為2n=2x=14(圖2-D),變異植株染色體為2n=4x=28(圖2-E)、2n=4x=26(圖2-F)。經(jīng)鑒定,共獲得歐洲溫室型黃瓜同源四倍體新種質(zhì)4個、非整倍體新種質(zhì)1個。
2.3?二倍體與同源四倍體歐洲溫室型黃瓜比較
2.3.1?植株形態(tài)比較
二倍體與同源四倍體黃瓜差異顯著性分析表明,子葉節(jié)長和株高達到極顯著差異,子葉節(jié)粗差異顯著(表2)。與二倍體相比,同源四倍體黃瓜的子葉大小、葉片大小呈極顯著增大,此外,同源四倍體黃瓜的葉片顏色加深、葉片厚度增加、葉片表面褶皺。
同源四倍體黃瓜與二倍體相比,除雌花子房長差異不顯著外,雄花和雌花花冠大小、花瓣大小均明顯增大,花期子房直徑和果柄長均達到極顯著差異,且花冠褶皺程度高(表3、圖3)。
2.3.2?顯微特征比較
由表4可知,與二倍體相比,同源四倍體歐洲溫室型黃瓜葉片下表皮氣孔保衛(wèi)細胞大小、單位面積葉綠體數(shù)量均呈現(xiàn)極顯著增加,分別為二倍體的1.52、203倍;氣孔密度極顯著減少,為二倍體的0.34倍。二倍體黃瓜花粉為三孔型(圖4-A和4-C),同源四倍體花粉除正常三孔型外,還出現(xiàn)四孔型、五孔型花粉(圖4-B、4-D和4-E),且敗育花粉比率較高,達70%以上(圖4-F)。
2.3.3?果實形態(tài)與品質(zhì)比較
在果實形態(tài)與品質(zhì)特征方面,與二倍體親本相比,同源四倍體黃瓜果實長度、直徑、中心腔直徑、果肉厚、瓜把長等均較小,但含糖量顯著提高(表5)。
3?討論與結(jié)論
人工誘導多倍體是植物育種的一個重要手段,也是創(chuàng)制新種質(zhì)的重要途徑。陳勁楓等對采用不同濃度秋水仙素溶液浸泡干種子的方法獲得黃瓜同源四倍體,發(fā)現(xiàn)4 g/L秋水仙素浸萌動種子4 h的染色體加倍率可達26.7%[6]。本研究采用秋水仙素處理幼苗生長點的方法,發(fā)現(xiàn)以2、3 g/L 秋水仙素處理5次誘導效果較好。低濃度處理次數(shù)較少時,成苗率較高,而在較高濃度處理次數(shù)較多時,成苗率降低,這與孔素萍等的研究結(jié)果[19]相一致。在本研究中,黃瓜幼苗的死亡率與秋水仙素處理時間和濃度呈正相關。
二倍體與同源四倍體植株在形態(tài)學、生理學、細胞學等方面均存在較大差異,研究表明,同源四倍體植株葉片氣孔大小、花粉粒大小、雌雄花大小、葉片大小與厚度、保衛(wèi)細胞內(nèi)葉綠體數(shù)量等均高于二倍體親本,而植株高度、氣孔密度均低于二倍體親本[13,20]。本研究中二倍體親本黃瓜通過秋水仙素誘變后,在形態(tài)學、解剖學、細胞學等方面均表現(xiàn)出不同程度的畸形現(xiàn)象。首先利用誘變植株的形態(tài)學特征、氣孔特征進行變異植株初步篩選,再對候選植株進行DNA相對含量和染色體數(shù)目鑒定確定其倍性水平,極大程度提高了多倍體植株篩選的工作效率。
多倍體雄花和花粉直徑均大于二倍體,而且花粉形態(tài)異常,萌發(fā)率低[21],多倍化容易導致植物育性降低[22-24]。陳勁楓等發(fā)現(xiàn),四倍體花粉粒大小不一,每一朵花中平均有2.3%的4孔花粉粒[6]。Diao等發(fā)現(xiàn),四倍體植株自交后代在細胞遺傳水平上較穩(wěn)定,但二倍體與同源四倍體黃瓜育性上存在較大差異[25]。本研究發(fā)現(xiàn),變異植株花粉出現(xiàn)四孔型、五孔型等形態(tài)變異類型,且同源四倍體黃瓜花粉有較高比率的敗育現(xiàn)象,可能是導致自交授粉后單瓜結(jié)籽率低的主要原因。本研究為黃瓜遺傳育種提供了新種質(zhì),為雙子葉植物多倍體育種提供了技術支持。
參考文獻:
[1]顧少涵,潘?俏,李子昂,等. 黃瓜耐低溫突變體材料的篩選與鑒定[J]. 南京農(nóng)業(yè)大學學報,2017,40(2):219-224.
[2]管?葦,張云霞,楊樹瓊,等. 黃瓜倍性材料創(chuàng)制及染色體組成的FISH鑒定[J]. 中國農(nóng)業(yè)科學,2014,47(17):3513-3522.
[3]徐麗娟,周鴻章,張蜀寧,等. 優(yōu)質(zhì)、抗熱同源四倍體不結(jié)球白菜的創(chuàng)制及鑒定[J]. 福建農(nóng)業(yè)學報,2014,29(5):449-453.
[4]呂?煒,戶秋穩(wěn),李艷艷,等. 優(yōu)質(zhì)四倍體蘿卜新種質(zhì)創(chuàng)新及鑒定[J]. 南京農(nóng)業(yè)大學學報,2016,39(1):48-54.
[5]羅小金,賀浩華. 水稻多倍性育種研究進展[J]. 中國農(nóng)學通報,2001,17(6):53-56.
[6]陳勁楓,雷?春,錢春桃,等. 黃瓜多倍體育種中同源四倍體的合成和鑒定[J]. 植物生理學通訊,2004,40(2):149-152.
[7]Blasco M,Luisa B M,Del M N M. Colchicine-induced polyploidy in loquat[Eriobotrya japonica (Thunb.) Lindl.][J]. Plant Cell Tissue and Organ Culture,2015,120(2):453-461.
[8]Amaral C,Silva S,Ledo C,et al. Agronomic characterization of autotetraploid banana plants derived from‘PisangLilin(AA)obtained through chromosome doubling[J]. Euphytica,2015,202(3):435-443.
[9]Xing S H,Guo X B,Wang Q,et al. Induction and flow cytometry identification of tetraploids from seed-derived explants through colchicine treatments in Catharanthus roseus (L.) G. Don[J]. Journal of Biomedicine & Biotechnology,2011(1):793198.
[10]李豆豆,劉芝龍,黃明忠,等. 鶴頂蘭四倍體植株的誘導與鑒定[J]. 園藝學報,2013,40(10):2033-2038.
[11]Chen R,Jiang W Z,Li Q L,et al. Comparison of seven colchicine-induced tetraploid clones with their original diploid clones in purple coneflower(Echinacea purpurea L.)[J]. Euphytica,2016,207(2):387-399.
[12]王朝梁,陳中堅,孫玉琴,等. 秋水仙堿誘導三七多倍體的初步研究[J]. 中國中藥雜志,2007,32(12):1222-1224.
[13]張海鳳,郭寶林,張成合,等. 陳新華.杜仲四倍體的誘導與鑒定[J]. 園藝學報,2008,35(7):1047-1052.
[14]Madani H,Hosseini B,Dehghan E,et al. Enhanced production ofscopolamine in induced autotetraploid plants of HyoscyamusreticulatusL.[J]. ActaPhysiologiaePlantarum,2015,37(3):55.
[15]刁衛(wèi)平,賈媛媛,江?彪,等. 黃瓜未授粉子房培養(yǎng)獲得同源四倍體[J]. 園藝學報,2008,35(12):1781-1786.
[16]鄢郁霖,王小蓉,唐海東,等. 秋水仙素處理對黃瓜生長發(fā)育及誘變效應研究[J]. 長江蔬菜,2010(6):8-11.
[17]劉永月,許建鵬,李田田,等. 無毛黃瓜同源四倍體誘導及鑒定[J]. 山東農(nóng)業(yè)科學,2016,48(11):21-25.
[18]鄭金雙,張蜀寧,孫成振,等. 二倍體及同源四倍體不結(jié)球白菜核型分析[J]. 南京農(nóng)業(yè)大學學報,2012,35(1):131-134.
[19]孔素萍,段乃彬,劉冰江,等. 不同誘變劑對大蒜四倍體誘導的影響[J]. 核農(nóng)學報,2016,30(6):1067-1073.
[20]Abdoli M,Moieni A,Badi H N. Morphological,physiological,cytological and phytochemical studies in diploid and colchicine-induced tetraploid plants of Echinacea purpurea (L.)[J]. ActaPhysiologiaePlantarum,2013,35(7):2075-2083.
[21]Lin L,Yao Q C,Xu H W,et al. Characteristics of the staminate flower and pollen from autotetraploid Betulaplatyphylla[J]. Dendrobiology,2013,69(1):3-11.
[22]軒淑欣,羅雙霞,張成合,等. 同源四倍體結(jié)球甘藍胚胎發(fā)育和結(jié)實性研究[J]. 河北農(nóng)業(yè)大學學報,2009,32(1):14-18.
[23]劉富強,張巍巍,蔡麗靜,等. 大白菜DH系同源四倍體的獲得及其細胞學研究[J]. 河北農(nóng)業(yè)大學學報,2013,36(5):24-28.
[24]張巍巍,劉富強,軒淑欣,等. 四倍體黑芥的獲得與鑒定[J]. 園藝學報,2015,42(12):2505-2511.
[25]Diao W P,Bao S Y,Jiang B,et al. Primary trisomics obtained from autotriploid by diploid reciprocal crosses in cucumber[J]. Sexual Plant Reproduction,2009,22(1):45-51.
