白樺8個MYB基因的克隆及其序列分析
2019-11-28 10:54:11王蓮萍王博楊春雨國會艷任如意
江蘇農業科學 2019年18期
王蓮萍 王博 楊春雨 國會艷 任如意
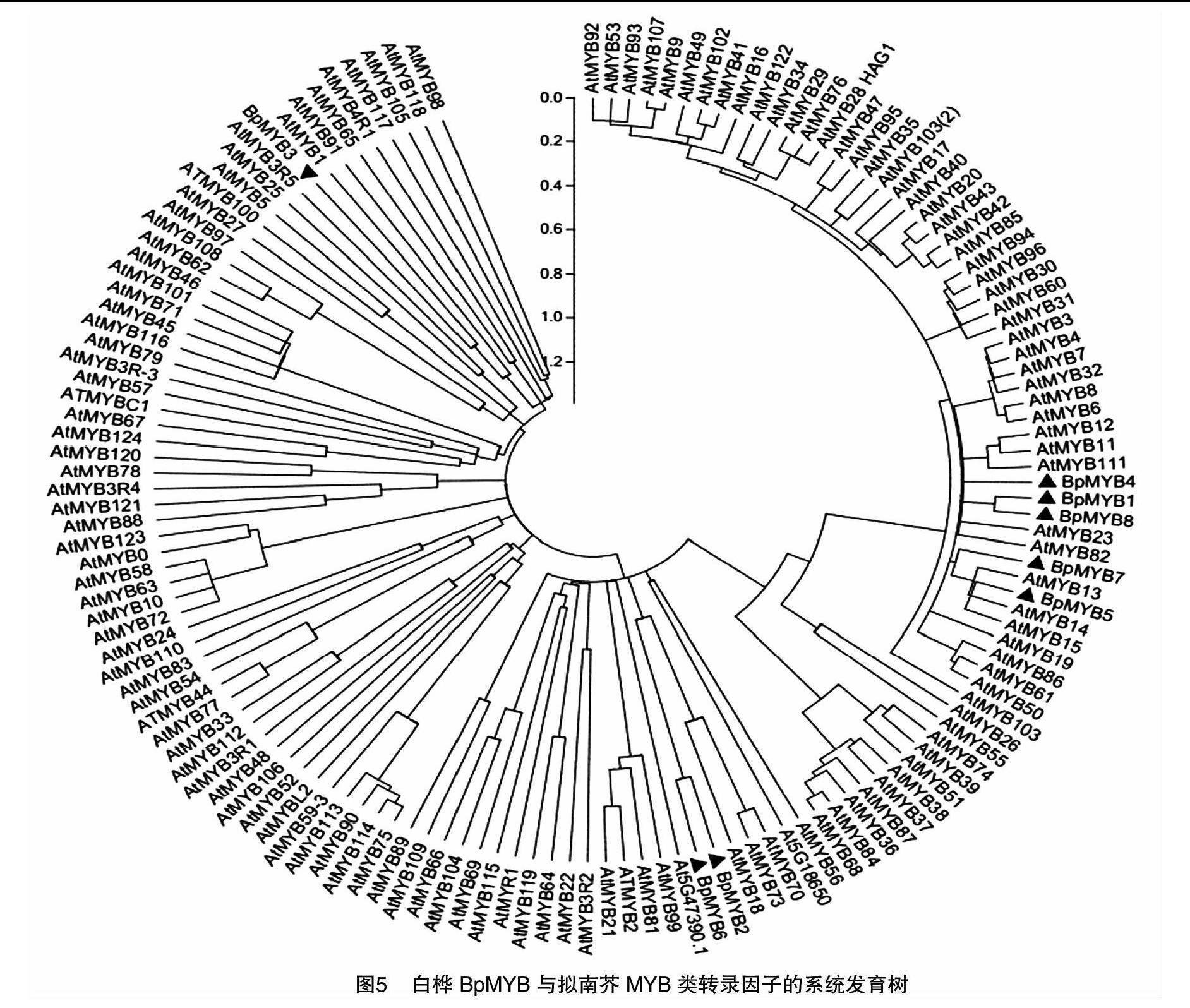
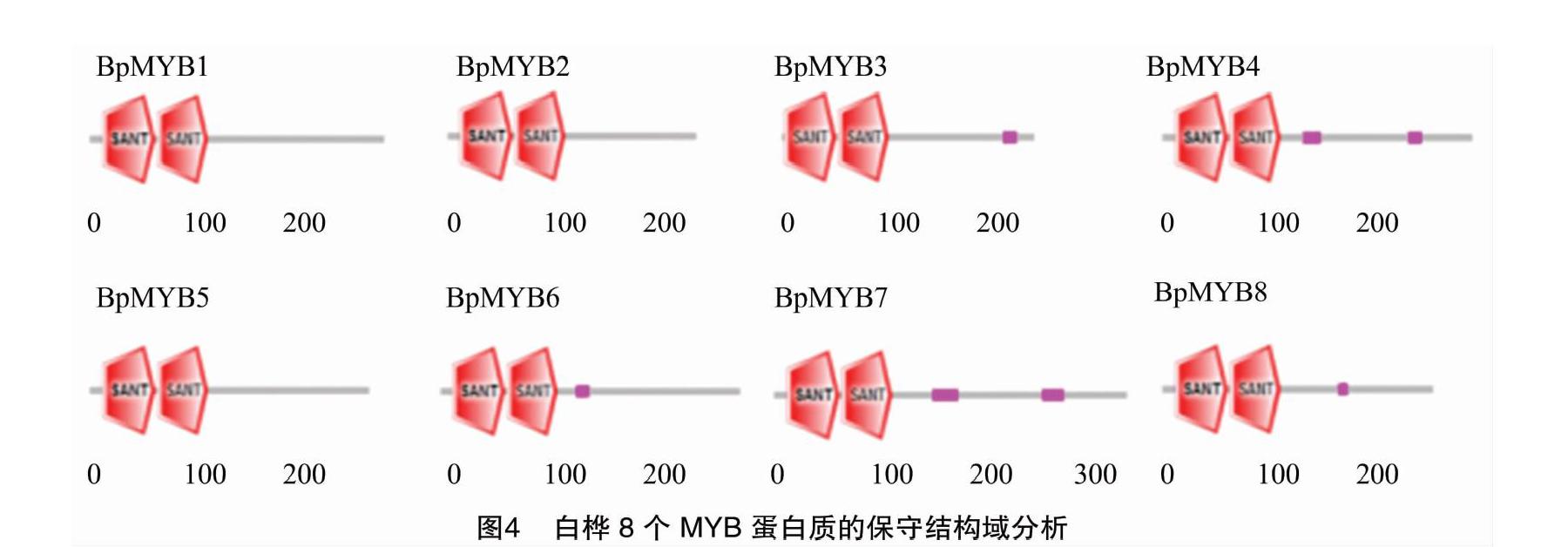

摘要:根據白樺MYB基因編碼區設計引物,以白樺的cDNA為模板,擴增出8個MYB基因,純化回收后與T載體進行連接并利用熱激法轉入大腸桿菌感受態細胞中,后經PCR檢測成功將8個MYB基因構建到T載體上。對8個MYB基因進行生物信息學分析,結果表明,8個MYB蛋白質的分子量在27.2~38.8 ku之間,等電點在5.38~8.66之間;并且,8個MYB均為核蛋白質,但都沒有跨膜結構域,屬于非跨膜蛋白質;系統進化樹分析結果預測8個MYB基因在植物生長發育的各個方面發揮著重要的作用,為后續研究8個MYB基因在白樺中的功能以及通過優良基因改良白樺的品質性狀提供前期數據支持。
關鍵詞:白樺;MYB基因;序列分析
中圖分類號: S188文獻標志碼: A
文章編號:1002-1302(2019)18-0093-05
收稿日期:2018-05-15
基金項目:黑龍江省教育廳項目(編號:1352MSYYB002);研究生創新項目(編號:kjcx2018-80mdjnu)。
作者簡介:王蓮萍(1994—),女,黑龍江省鶴崗人,碩士研究生,主要從事植物分子生物學與基因工程研究。E-mail:928718821@qq.com。
通信作者:任如意,博士,教授,主要從事植物分子生物學與基因工程研究。E-mail:swxrry@126.com。
轉錄因子也稱反式作用因子,是指能夠與基因啟動子區域中順式作用元件發生特異性相互作用的DNA結合蛋白質,通過它們之間以及與其他相關蛋白質之間的相互作用激活或抑制某些基因的轉錄[1]。根據轉錄因子保守域可分為很多家族,如bZIP、bHLH、MYB等。
MYB蛋白質普遍存在與動植物中,是植物最大的轉錄因子家族之一,它參與植物次生代謝調控、激素和環境的應答、并對細胞分化、細胞周期、器官的形成以及植物葉片的形態建成具有重要的調節作用[2-3]。自Paz-ares等首次報道玉米轉錄因子C1基因的克隆[4]以來,從番茄[5]、月季[6]、水稻[7]等多種植物中都分離克隆了MYB基因,并體現出MYB蛋白質功能的多樣性。植物體中大多數MYB蛋白質有2個重復MYB結構域(R2、R3),根據C端保守氨基酸序列的不同R2R3-MYB可分為22個亞類,同一亞類的基因往往具有相對保守的功能[8],有學者推測在擬南芥中有130多個R2R3-MYB基因[9],玉米中至少有80個MYB基因[10]。MYB家族轉錄因子有很多的成員被鑒定為是次生壁合成重要的調控因子[11],MYB轉錄因子在植物應答生物脅迫中具有重要的作用,在植物的生長過程中,幾乎參與了植物的發育和代謝的各個方面。楊樹的PtrMYB092和PtrMYBJ52基因參與了植物中木質素合成的調控,并能通過激活下游轉錄因子和關鍵酶基因的表達提高木質素的合成速率,促進植物細胞次生壁發生木質化[12]。有學者從桉樹克隆到了參與調控樹木木質部形成的關鍵基因,桉樹中的Eg MYB2基因可以調控木質素合成基因的轉錄,從而影響了桉樹木質素和次生壁的形成[13]。Zhong等研究表明擬南芥的AtMYB58和AtMYB63過表達時,植物細胞壁的木質素含量顯著增加,而其他細胞壁成分沒有變化[14]。擬南芥中HbMYB85a/b過表達都能使導管、木質纖維和維管束間纖維的細胞壁厚度增加,木質素含量升高,木質素合成相關酶基因表達升高,葉片量加厚,抽薹開花延遲[15]。McCarthy等在楊樹中克隆了MYB46、MYB83的同源基因Ptr MYB3、Ptr MYB20,最后通過功能分析證實了Ptr MYB3、Ptr MYB20參與了次生木質部的合成[16]。小麥中的TaMYB33受ABA介導的逆境響應信號調控,通過積累滲透壓調節物和提高細胞活性氧清除能力來提高植物對干旱和鹽的耐受性[17]。Yang等研究發現OsMYB2過表達植株中可溶性糖和脯氨酸含量上升,并發現其細胞內脯氨酸合成酶和轉運蛋白基因等逆境相關基因表達量上升,同時抗氧化酶的活性升高,對低溫、鹽害和干旱逆境脅迫的耐受性都明顯增強[18]。擬南芥中的MYB68基因在根中柱鞘細胞中特異型表達,在高溫脅迫下,根中MYB68表達會明顯增強,并且MYB68突變體的生長活力比野生型低,說明MYB68參與了擬南芥高溫脅迫的應答反應[19]。水稻Osmyb4基因的過表達大大提高了轉基因水稻對干旱、高鹽、紫外輻射等的耐受能力[20]。從以往研究中不難發現,MYB轉錄因子在植物中起著關鍵性作用,但其作用的詳細分子機理還沒有被完全闡述清楚,因此,有必要對MYB轉錄因子的功能和作用機制繼續進行深入研究。
白樺(Betula platyphylla Suk.)為樺木屬的落葉喬木,其木材可供一般建筑及制作器具,樹皮可以提煉樺油,樹汁在保健藥用上有抗衰老止咳等功效,植株本身可用于園林景觀。隨著對白樺不斷深入的研究,它的經濟市場價值也將不斷地被開發出來。本研究從白樺轉錄組中找到8個MYB基因,并以白樺cDNA為模板將其克隆出來,與T載體進行連接,轉化至大腸桿菌中。并對8個MYB基因進行生物信息學分析,為后續利用試驗手段深入研究這8個MYB基因的功能提供前期依據。
1?材料與方法
1.1?材料
1.1.1?試驗材料
白樺組培苗,由牡丹江師范學院生命科學與技術學院提供。
1.1.2?試驗試劑
大腸桿菌DH5α感受態細胞、2×Power Taq PCR MasterMIX混合酶(百泰克生物技術公司)、DL2000 DNA marker。膠回收試劑盒、PMD18-T載體試劑盒、質粒提取試劑盒(天根生物技術有限公司)。
1.1.3?培養基
培養基配方。1/2 MS固體培養基:1/2 MS粉(不含瓊脂粉和蔗糖)2.47 g,瓊脂粉8 g,蔗糖30 g,加蒸餾水定容至1 g,調節pH值為5.8。LB液體培養基:10 g蛋白胨、10 g NaCl、5 g酵母粉,加蒸餾水定容至1 L,調節pH值為7.0。
3?結論與討論
MYB作為最大的轉錄因子家族之一,根據所含保守結構域的不同分為1R、2R、3R、4R多個亞家族,不同的亞家族也具有著不同的結構特征和功能。從白樺轉錄組中找到8個MYB基因,在從白樺的cDNA中克隆出這8個基因,與擬南芥MYB家族基因進行系統進化分析,暗示這8個白樺MYB基因在白樺生長發育的各個方面具有不同的功能,但這只是一種預測,后續還需要進行轉基因試驗來證實,本研究為進一步利用試驗手段分析MYB基因在白樺中的功能提供前期基礎數據。
參考文獻:
[1]張椿雨,龍?艷,馮?吉,等. 植物基因在轉錄水平上的調控及其生物學意義[J]. 遺傳,2007,29(7):793-799.
[2]Martin C,Paz-Ares J. MYB transcription factors in plants[J]. Trends in Genetics,1997,13(2):67-73.
[3]王?棟,李利紅,陳志玲,等. 擬南芥根特異表達轉錄因子AtMYB305的鑒定及功能研究[J]. 科學通報,2001,46(21):1804-1809.
[4]Paz-Ares J,Ghosal D,Wienand U,et al. The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators[J]. EMBO J,1987,6(12):3553-3558.
[5]蔡新忠,徐幼平,樓?健. 番茄MYB基因片段的克隆及在過敏性反應時的表達[J]. 園藝學報,2003,30(5):589-591.
[6]謝吉容,熊運海,程在全,等. 月季MYB基因cDNA全長克隆和表達分析[J]. 中國農業科學,2008,41(12):4173-4179.
[7]陳?俊,朱?英,王宗陽. 水稻MYB cDNA的克隆和表達分析[J]. 植物生理與分子生物學學報,2002,28(4):267-274.
[8]王希慶,陳柏君,印莉萍. 植物中的MYB轉錄因子[J]. 生物技術通報,2003(2):22-25.
[9]Stracke R,Werber M,Weisshaar B. The R2R3-MYB gene family in Arabidopsis thaliana[J]. Current Opinion in Plant Biology,2001,4(5):447-456.
[10]Rabinowicz P D,Braun E L,Wolfe A D,et al. Maize R2R3 Myb genes:Sequence analysis reveals amplification in the higher plants[J]. Genetics,1999,153(1):427-444.
[11]郭?偉,趙樹堂,盧孟柱. 煙草和毛白楊次生維管發育相關MYB轉錄因子分析[J]. 林業科學研究,2016,29(1):25-33.
[12]李超鋒. 楊樹PtrMYB092和PtrMYB152轉錄因子在木質素合成中的功能研究[D]. 重慶:西南大學,2014.
[13]Monica G,Eric L,Sylvain L,et al. EgMYB2,a new transcriptional activator from Eucalyptus xylem,regulates secondary cell wall formation and lignin biosynthesis[J]. The Plant Journal,2005,43(4):553-567.
[14]李維靜,蘇衍菁,蘇相亭,等. MYB轉錄因子在植物木質素合成中的調控機理[J]. 山東畜牧獸醫,2013(11):69-71.
[15]劉?彤. 橡膠樹木質素合成相關MYB基因的功能鑒定[D]. 海口:海南大學,2015.
[16]McCarthy R L,Zhong R,Fowler S,et al. The poplar MYB transcription factors,PtrMYB3 and PtrMYB20,are involved in the regulation of secondary wall biosynthesis[J]. Plant & Cell Physiology,2010,51(6):1084-1090.
[17]Qin Y,Wang M,Tian Y,et al. Over-expression of TaMYB33 encoding a novel wheat MYB transcription factor increases salt and drought tolerance in Arabidopsis[J]. Molecular Biology Reports,2012,39(6):7183-7192.
[18]Yang A,Dai X,Zhang W H. A R2R3-type MYB gene,OsMYB2,is involved in salt,cold,and dehydration tolerance in rice[J]. Journal of Experimental Botany,2012,63(7):2541-2556.
[19]Feng C P,Andreasson E,Maslak A,et al. Arabidopsis MYB68 in development and responses to environmental cues[J]. Plant Science,2004,167(5):1099-1107.
[20]Vannini C,Locatelli F,Bracale M,et al. Overexpression of the rice Osmyb4 gene increases chilling and freezing tolerance of Arabidopsis thaliana plants[J]. The Plant Journal:for Cell and Molecular Biology,2004,37(1):115-127.
[21]Kirik V,Lee M M,Wester K,et al. Functional diversification of MYB23 and GL1 genes in trichome morphogenesis and initiation[J]. Development,2005,132(7):1477-1485.
[22]Kim J H,Nguyen N H,Jeong C Y,et al. Loss of the R2R3 MYB,AtMyb73,causes hyper-induction of the SOS1 and SOS3 genes in response to high salinity in Arabidopsis[J]. Journal of Plant Physiology,2013,170(16):1461-1465.
[23]Verma N,Burma P K. Regulation of tapetum-specific A9 promoter by transcription factors AtMYB80,AtMYB1 and AtMYB4 in Arabidopsis thaliana and Nicotiana tabacum[J]. The Plant Journal:for Cell and Molecular Biology,2017,92(3):481-494.
[24]Stracke R,Jahns O,Keck M,et al. Analysis of production of flavonol glycosides-dependent flavonol glycoside accumulation in Arabidopsis thaliana plants reveals MYB11-,MYB12- and MYB111-independent flavonol glycoside accumulation[J]. The New Phytologist,2010,188(4):985-1000.
[25]Chen Y,Chen Z,Kang J,et al. AtMYB14 regulates cold tolerance in Arabidopsis[J]. Plant Molecular Biology Reporter,2013,31:87-97.
[26]徐?玨. 轉錄因子AtMYB13抑制擬南芥防衛反應的研究[D]. 南京:南京農業大學,2009.
