我國棉花形態(tài)標(biāo)記性狀應(yīng)用研究進(jìn)展
2019-11-28 10:54:11陳瑩張法銘姜輝柴啟超王秀麗高明偉王家寶張超王永翠鄭錦秀趙軍勝
江蘇農(nóng)業(yè)科學(xué) 2019年18期
陳瑩 張法銘 姜輝 柴啟超 王秀麗 高明偉 王家寶 張超 王永翠 鄭錦秀 趙軍勝
摘要:本文綜述棉花主要形態(tài)標(biāo)記性狀的相關(guān)應(yīng)用研究進(jìn)展,旨在為形態(tài)標(biāo)記性狀在棉花遺傳育種和分子育種中的應(yīng)用提供參考。通過對我國目前具備形態(tài)標(biāo)記性狀的審定棉花品種進(jìn)行分類和統(tǒng)計(jì)發(fā)現(xiàn),芽黃標(biāo)記主要被用于與不育基因連鎖實(shí)現(xiàn)不育系在苗期的早期鑒定選擇,有助于提高棉花雜交制種產(chǎn)量和效益,育成品種1個(gè)。雞腳葉標(biāo)記棉具有良好的冠層結(jié)構(gòu)及光合生理特性,因此具有一定的早熟性、耐旱性和較好的抗病蟲性,在三系雜交棉生產(chǎn)上也有較好的應(yīng)用前景,育成品種3個(gè)。紅花標(biāo)記作為非常直觀的形態(tài)性狀標(biāo)記,可用于簡便高效地鑒別真假雜種,其中山東棉花研究中心培育審定了4個(gè)紅花標(biāo)記新品種。腺體標(biāo)記除了是直觀的形態(tài)性狀標(biāo)記以外,還具有低酚少腺體的特點(diǎn),山東棉花研究中心培育審定2個(gè)低酚標(biāo)記的抗蟲雜交棉新品種,邯鄲市農(nóng)業(yè)科學(xué)院與河北農(nóng)業(yè)大學(xué)審定了3個(gè)抗蟲低酚棉品種。其他形態(tài)標(biāo)記如窄卷苞葉性狀、紅葉性狀等在雜種優(yōu)勢利用方面也有較好的生產(chǎn)應(yīng)用潛力。以上各類形態(tài)性狀標(biāo)記在棉花生產(chǎn)上均具有廣泛的應(yīng)用前景,需進(jìn)一步提高標(biāo)記利用率,加強(qiáng)其應(yīng)用基礎(chǔ)研究。
關(guān)鍵詞:棉花;形態(tài)標(biāo)記;芽黃;雞腳葉;紅花標(biāo)記;腺體標(biāo)記
中圖分類號: S562.03文獻(xiàn)標(biāo)志碼: A
文章編號:1002-1302(2019)18-0046-04
收稿日期:2018-10-16
基金項(xiàng)目:國家自然科學(xué)基金(編號:31601348);國家現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系——棉花產(chǎn)業(yè)技術(shù)體系(編號:CARS-15-05);山東省重點(diǎn)研發(fā)計(jì)劃(編號:2017GNC10114)。
作者簡介:陳?瑩(1979—),女,山東濟(jì)寧人,碩士,副研究員,主要從事棉花育種及相關(guān)生物學(xué)研究及科研管理。E-mail:chenyingkb@126.com。
通信作者:趙軍勝,博士,研究員,主要從事棉花育種研究。E-mail:zhaojunshengsd@163.com。
在各種作物遺傳育種過程中,標(biāo)記性狀被廣泛用于品種純度鑒定和種質(zhì)資源的遺傳多樣性分析。目前,標(biāo)記主要包括形態(tài)標(biāo)記、細(xì)胞學(xué)標(biāo)記、生化標(biāo)記和DNA分子標(biāo)記,其中形態(tài)標(biāo)記和DNA分子標(biāo)記應(yīng)用較多。DNA分子標(biāo)記雖不受環(huán)境和組織特異性的影響,可客觀準(zhǔn)確地反映遺傳個(gè)體間的差異,但是需要借助于實(shí)驗(yàn)儀器,限制了其在大田生產(chǎn)中的應(yīng)用。雖然部分形態(tài)標(biāo)記易受環(huán)境影響,但是大部分較為穩(wěn)定,易于觀察,有助于田間的快速有效選擇。棉花雜種優(yōu)勢較強(qiáng),高純度的雜交種是實(shí)現(xiàn)棉花雜種優(yōu)勢利用的重要前提和保證。在大田制種過程中,借助于形態(tài)標(biāo)記可有效去除雜株,保證制種純度。由此可見,形態(tài)標(biāo)記研究對棉花生產(chǎn)具有重要意義。因此,本文就棉花主要形態(tài)標(biāo)記性狀的相關(guān)應(yīng)用研究進(jìn)展進(jìn)行綜述,旨在為其在遺傳育種理論研究和生產(chǎn)中應(yīng)用提供一定依據(jù)。
1?我國棉花形態(tài)標(biāo)記應(yīng)用研究現(xiàn)狀
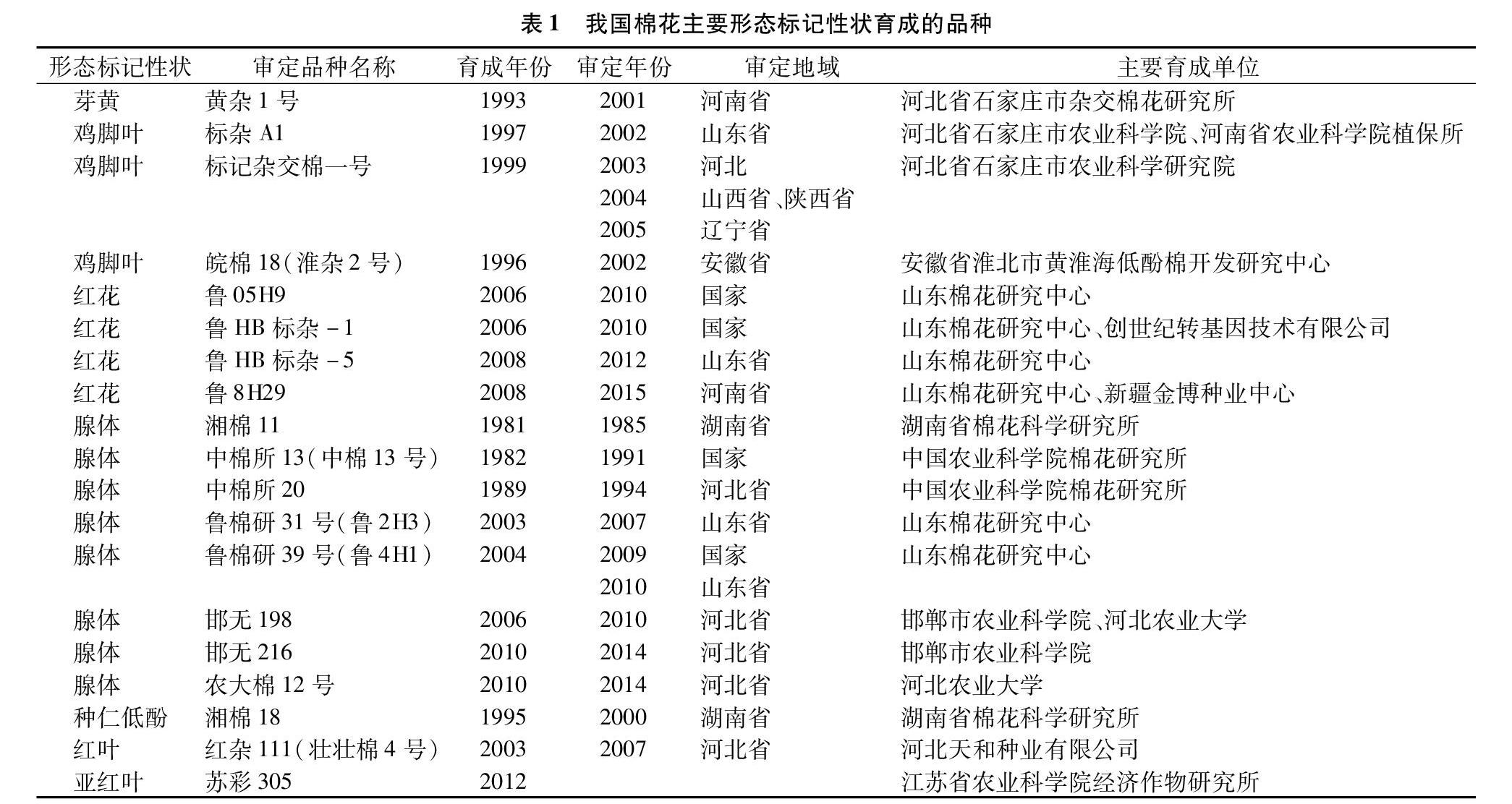
棉花形態(tài)標(biāo)記性狀大部分屬于質(zhì)量性狀[1],涉及18個(gè)性狀,其中研究較多的有芽黃、雞腳葉、紅花、腺體標(biāo)記性狀,研究較少的包括茸毛、紅株、紅葉、窄卷苞葉、黃花藥、光籽等標(biāo)記。在應(yīng)用層面上可分為2類:第1類,本身僅可作為形態(tài)標(biāo)記,如芽黃、紅花、植株茸毛、黃花藥等標(biāo)記;第2類,除了具有標(biāo)記作用外,還有其他功能性作用,如雞腳葉、腺體標(biāo)記等。經(jīng)不完全統(tǒng)計(jì),目前我國具備主要形態(tài)標(biāo)記性狀的育成、審定棉花品種見表1。
2?棉花形態(tài)標(biāo)記性狀類型
2.1?芽黃標(biāo)記
芽黃(virescent)是葉綠素含量較正常稀少的隱性突變,類型較多,主要特點(diǎn)是苗期真葉表現(xiàn)明顯的黃色,隨著植株生長,黃化葉片隨著葉綠素含量的不斷增加而逐漸變綠,達(dá)到正常水平。芽黃性狀表現(xiàn)明顯、穩(wěn)定且容易辨別,遺傳方式簡單,大多受1對隱性基因控制,是理想的標(biāo)記性狀,在棉花雜種優(yōu)勢利用領(lǐng)域已有很多研究。肖松華等研究發(fā)現(xiàn),同一芽黃基因?qū)氩煌z傳背景陸地棉后,對農(nóng)藝性狀影響的范圍和程度差異很大[2]。陸地棉芽黃品系與常規(guī)品系雜交,F(xiàn)1具有明顯的雜種優(yōu)勢,包括產(chǎn)量構(gòu)成因素、早熟性和纖維品質(zhì)性狀優(yōu)勢,尤其是產(chǎn)量性狀優(yōu)勢明顯[2-4]。王學(xué)德等經(jīng)親本配合力分析認(rèn)為,彭澤芽黃和湖北芽黃是較好的芽黃親本[5]。潘家駒等鑒定了6個(gè)芽黃基因?qū)ζ滢D(zhuǎn)育系經(jīng)濟(jì)性狀的影響并研究證實(shí)了這6個(gè)芽黃轉(zhuǎn)育系與陸地栽培品種的雜交組合的產(chǎn)量優(yōu)勢,證明了v10、v15、v20 3個(gè)芽黃基因作為標(biāo)記基因用于雜交棉制種的可能性[6]。
通過芽黃標(biāo)記與不育基因連鎖來實(shí)現(xiàn)不育系在苗期的早期鑒定選擇,可有效提高棉花雜交制種的產(chǎn)量和效益。其中,81A是攜帶芽黃標(biāo)記的陸地棉核雄性不育系,其在子葉期表現(xiàn)為淺黃色,1~3張真葉期表現(xiàn)為正常綠色,4葉期開始表現(xiàn)整株的淡黃色,7葉期后黃色漸褪,而到開花期表現(xiàn)為芽黃類型,即上部幾張葉片表現(xiàn)為淡黃色,下部葉片表現(xiàn)為正常綠色,遺傳研究證實(shí),其芽黃基因和雄性不育基因緊密連鎖,可以用于雜交制種母本篩選[7]。宇文璞等引入81A后,對芽黃A的溫敏感特性進(jìn)行了系統(tǒng)研究,發(fā)現(xiàn)并提出了芽黃A的溫控繁育法,選育出在子葉至3張真葉期就具有準(zhǔn)確芽黃標(biāo)記性狀、雜種優(yōu)勢顯著的溫敏性“芽黃A”雄性不育系;以此轉(zhuǎn)育成的芽黃8748A不育系配制的雜交種大部分組合優(yōu)勢明顯,產(chǎn)量高,抗逆性強(qiáng),其中用2145作父本配制的黃雜1號,表現(xiàn)出早發(fā)早熟、有效鈴多、鈴大、衣分高、增產(chǎn)明顯、穩(wěn)產(chǎn)性強(qiáng)、纖維品質(zhì)好等優(yōu)良性狀,生產(chǎn)示范應(yīng)用綜合表現(xiàn)突出[8-9]。1998年湖南農(nóng)業(yè)大學(xué)發(fā)現(xiàn)了高溫敏感型棉花雄性不育材料,培育出穩(wěn)定的棉花溫敏雄性不育系特棉S-1,其育性在日平均溫度<26.5 ℃時(shí)表現(xiàn)為可育,>27.0 ℃時(shí)表現(xiàn)為不育[10-11]。
棉花光溫敏雄性不育系是從海陸雜交種后代中發(fā)現(xiàn)的;具有芽黃標(biāo)記性狀的新型核雄性不育系,是對芽黃A經(jīng)過30年40多代選育出的棉花雄性不育系新類型[12-13]。芽黃396A是攜帶連鎖芽黃基因的棉花單隱性光溫敏核雄性不育系,品質(zhì)和農(nóng)藝性狀優(yōu)良,在高溫長日照或低溫短日照條件下表現(xiàn)為完全雄性不育,而在高溫短日照條件下表現(xiàn)為可育,可自交成鈴,不再需要專門的保持系,該不育系攜帶的芽黃標(biāo)記,在正常條件下,于出苗5~7 d的子葉期即可明顯區(qū)別可育株與不育株,在非正常條件下,最晚到3~5張真葉期即可區(qū)別[14]。牛金生等在新疆和三亞對396A的育性變化及其雜交種的產(chǎn)量性狀、抗病性、纖維品質(zhì)等開展試驗(yàn)研究,發(fā)現(xiàn)其雜交種F1代育性恢復(fù)好,花粉量大,全部能夠自然成鈴且雜種優(yōu)勢明顯[15]。張濤等對影響棉花光溫敏不育A系雜交成鈴的4個(gè)方面因素進(jìn)行分析研究,確定了雜交授粉的最佳授粉日期、時(shí)間段和日平均氣溫,發(fā)現(xiàn)以不同不育系為母本時(shí)的雜交成鈴率之間存在明顯差異[16]。
2.2?雞腳葉標(biāo)記
雞腳葉是陸地棉正常闊葉的顯性突變類型之一,具有裂刻極深、葉裂片窄的特點(diǎn)。雞腳葉標(biāo)記棉單株和群體通風(fēng)透光性好,因此具有良好的冠層結(jié)構(gòu)及光合生理特性。朱偉研究發(fā)現(xiàn),雞腳葉葉面積比正常葉縮小12.94%~25.59%,大大改善了棉田光照條件[17]。同鴨掌葉比較,雞腳葉葉綠素含量增加12.77%~39.36%,光合作用強(qiáng)度高17.5%,光照度增加12.77%~39.36%,透光度增加12.90%~39.45%,且中下層增加趨勢更明顯[17-18]。杜明偉等研究發(fā)現(xiàn),雞腳葉棉上部冠層光吸收率為38%~47%,中下部冠層為22%~31%,整體冠層的光分布均勻;而正常闊葉棉上部冠層光吸收率為65%~70%,極顯著高于中下部;雞腳葉棉冠層群體光合速率分布比正常闊葉棉更均衡,特別是在盛鈴后期,雞腳葉棉上部、中部、下部冠層的群體光合速率分別占總體光合速率的40.9%~46.7%、26.8%~29.6%、18.5%~21.5%,而正常闊葉棉則分別占61.1%~72.3%、21.7%~25.9%、102%~15.3%[19-20]。
雞腳葉棉還具有一定的早熟性、耐旱性。雞腳葉棉的果節(jié)出現(xiàn)速率比普通葉棉高50%~100%,開花前期每天開花3.9朵,而正常闊葉棉每天開花1.8朵;雞腳葉棉的霜前花率為81.8%,比普通葉棉高12.3%[21-22]。雞腳葉的蒸騰系數(shù)小于正常闊葉,可使葉片在干旱狀態(tài)下處于有利保持水分的狀態(tài)[23-24]。用雞腳葉(或超雞腳葉)作親本配制的雜交棉也顯示了早熟性,如標(biāo)雜A1在河南省區(qū)試中,霜前皮棉產(chǎn)量比對照極顯著增加36.6%[17]。
由于綜合了形態(tài)抗性、生態(tài)抗性和生理生化抗性,雞腳葉棉花一般抗棉鈴蟲、紅鈴蟲、棉大卷葉螟、斜紋夜盜蛾、抗葉螨等[17,25]。雞腳葉品種棉田透光性好、田間濕度低,所以喜濕或喜郁蔽條件的黏蟲、玉米螟等害蟲的生長發(fā)育也受到抑制[26];雞腳葉棉對角斑病的18個(gè)生理小種均具有抗性[25],極大地限制了棉鈴疫病的發(fā)生、發(fā)展[17]。另外,雞腳葉有利于農(nóng)藥穿透,能大大提高化學(xué)防治病蟲害效果;雞腳葉棉花一般較矮,株型緊湊,能抗風(fēng)暴[25]。
美國在1976、1981年先后發(fā)放了Gumbo等雞腳葉陸地棉品種[27]。澳大利亞從1984年開始發(fā)放育成的雞腳葉品種,至20世紀(jì)90年代,雞腳葉品種種植面積已經(jīng)達(dá)到澳大利亞植棉面積的45%以上,主要種植品種有Siokral-4、Siokra L23、Siokra S324、SiokraV1等[25]。我國曾育成超雞腳葉川簡3號、雞腳紫絮棉等品種,雞腳德字棉在四川省推廣種植10多萬hm2。目前,通過省級審定的品種中包括3個(gè)雜交種——通過河南省、山東省審定的標(biāo)雜A1[28],通過河北省、山西省、陜西省、遼寧省4個(gè)省審定的標(biāo)記雜交棉一號[29],通過安徽省審定的皖棉18(淮雜2號)[30]等。朱偉以超雞腳葉、雞腳葉和正常葉棉花不育系及其恢復(fù)系為雜交親本配制組合,驗(yàn)證發(fā)現(xiàn),雜種F1代由于單鈴質(zhì)量和衣分增加出現(xiàn)皮棉增產(chǎn)342%(增產(chǎn)極顯著)的情況,表明雞腳葉標(biāo)記在三系雜交棉生產(chǎn)上具有廣泛的應(yīng)用前景;另外發(fā)現(xiàn),雞腳葉雜交種雙親自交系可增強(qiáng)優(yōu)勢,通過自交系配組可使其功能充分展現(xiàn)[17]。
2.3?紅花標(biāo)記
20世紀(jì)80年代,由原中國科學(xué)院遺傳研究所牽頭的棉花遠(yuǎn)緣雜交協(xié)作組,用陸地棉與比克氏棉進(jìn)行遠(yuǎn)緣雜交,選育出了具有粉紅色花瓣和紅色大基斑的紅花純合材料,定名為“HB(hirsutum-bickii)紅花系”[31]。趙軍勝等利用4個(gè)陸地棉HB紅花近等基因系作父本,分別與輪回親本雜交組合,研究紅花標(biāo)記性狀棉花在產(chǎn)量雜種優(yōu)勢中的表現(xiàn),同時(shí)將HB紅花近等基因系及輪回親本作父本分別與共同母本雜交,比較各組F1有關(guān)性狀及產(chǎn)量等方面的超親、平均及競爭優(yōu)勢。結(jié)果表明,HB紅花標(biāo)記雜交組合籽棉與皮棉產(chǎn)量具有明顯的雜種優(yōu)勢,野生二倍體比克氏棉的漸滲片段對陸地棉雜種優(yōu)勢有重要的正向效應(yīng)[32]。山東棉花研究中心通過組配強(qiáng)優(yōu)勢雜交組合,育成并審定了魯05H9(國審棉2010003)、魯HB標(biāo)雜-1(國審棉2010004)、魯HB標(biāo)雜-5(魯農(nóng)審2012020號)、魯8H29(豫審棉2015010)等多個(gè)紅花標(biāo)記轉(zhuǎn)基因抗蟲雜交棉[33-34]。
2.4?腺體標(biāo)記
色素腺體為棉族植物特有,是棉屬植物分類的依據(jù)之一。棉花的色素腺體不僅是棉酚貯藏場所,還與棉酚的合成有著密切關(guān)系,但在不同類型的棉花之間,色素腺體與棉酚存在不同相關(guān)性[35]。棉酚對棉花抵抗某些病蟲害有重要作用,但是不利于種子營養(yǎng)成分的充分利用,因此,選育植株有腺體而種子無腺體或者有腺體無酚的品種是解決這一問題的有效方法。
腺體標(biāo)記可用于真假雜種的簡便辨認(rèn)。袁有祿等利用顯性無腺體陸地棉新品系N1為父本,以有腺體中棉所19和有腺體MS5MS6核不育系為母本,進(jìn)行不去雄人工輔助授粉,在后代中利用腺體標(biāo)記對真假雜種進(jìn)行分辨發(fā)現(xiàn),獲得真雜種率分別為52.4%和99.3%[36]。
棉酚對單胃動物具有毒害,選育腺體標(biāo)記的低酚棉品種是提高棉籽利用率的關(guān)鍵。我國的低酚棉育種研究開始于1972年,20世紀(jì)90年代達(dá)到高峰,當(dāng)時(shí)全國棉區(qū)育成不同類型低酚棉品種近60個(gè),推廣面積也快速增長;至1987年,我國低酚棉種植面積已經(jīng)超過美國和科特迪瓦共和國,居世界第1位,至1991年種植面積一直穩(wěn)定在10萬hm2以上,且以河北省、河南省、山東省棉區(qū)種植面積最大[37]。喻樹迅等成功選育了具有純合雙隱性基因gl2gl2gl3gl3的低酚棉品種中棉所20,該品種總棉酚含量僅為0.002 5%,遠(yuǎn)低于世界糧農(nóng)組織和衛(wèi)生組織規(guī)定的0.04%的標(biāo)準(zhǔn),而且還具有柱頭短、花絲長、散粉早的特點(diǎn),可大大降低異交率,在不隔離種植的條件下,純度可達(dá)98%以上[38]。祝水金等培育了具有子葉腺體延緩形成特性的陸地棉種質(zhì)系A(chǔ)BH-0318,該材料色素腺體性狀穩(wěn)定,休眠種子僅在子葉邊緣有少量腺體,棉酚含量僅為0.017 5%;種子萌發(fā)后,子葉和植株腺體數(shù)量與一般有酚品種相似,為培育植株有腺體而種子無腺體的棉花品種奠定了基礎(chǔ)[39]。山東棉花研究中心育成了具有低酚標(biāo)記性狀的轉(zhuǎn)基因抗蟲雜交棉品種魯棉研31號(山東省審定)和魯棉研39號(國家審定、山東省審定),并建立了魯棉研39號的指紋圖譜[40]。2010年河北省也首次審定了抗蟲低酚棉品種邯無198,2014年育成審定了邯無216、農(nóng)大棉12號2個(gè)抗蟲低酚棉[37,41]。劉芳等在新疆巴州對47個(gè)低酚棉品種資源的主要性狀進(jìn)行了調(diào)查分析,優(yōu)選了11個(gè)適宜的資源材料[42]。李函利等對179份不同來源的低酚棉種質(zhì)資源的耐鹽性進(jìn)行了評價(jià),篩選出8份達(dá)到了耐鹽和抗鹽水平的突出材料,為低酚棉育種篩選到了優(yōu)異種質(zhì)資源[43]。劉慧等研究了不同基因型棉花不同器官的色素腺體與棉酚旋光體含量間的相關(guān)性,結(jié)果發(fā)現(xiàn),低酚棉雖全株無色素腺體,但器官中也能檢測到低于0.02%的少量游離棉酚存在,有酚棉棉仁中色素腺體在各器官中最密,花瓣和棉仁色素腺體直徑最小,且棉仁和花瓣的總棉酚含量存在高度相關(guān)性,因此可以用花瓣的棉酚含量來判斷棉仁的棉酚含量[44]。
2.5?其他標(biāo)記
窄卷苞葉(Frego bract)是由美國阿肯色州棉農(nóng)Frego發(fā)現(xiàn)的,具有苞葉狹長、革質(zhì)化、向外扭翻生長,使花冠和棉鈴充分暴露的特性,受1對隱性基因(fg)控制。張金發(fā)等利用鄂沙78和7010 2個(gè)陸地棉品種作為母本,分別與窄卷苞葉棉Frego雜交組成1對近等基因系,與正常苞葉近等基因系試驗(yàn)比較,窄卷苞葉棉葉片光合速率下降、生長旺盛、不易早衰,對棉花產(chǎn)量、衣分、單株果枝數(shù)、脫落率和纖維品質(zhì)無明顯不良效應(yīng),在現(xiàn)蕾之前延遲棉花生育期,影響早熟,導(dǎo)致現(xiàn)蕾開花推遲,且對棉鈴象鼻蟲、棉鈴蟲等有一定的抗性[45-47]。
李俊蘭等在研究紅葉觀賞棉花的同時(shí),提出利用紅葉作為標(biāo)記性狀應(yīng)用于陸地棉雜種優(yōu)勢研究[48]。2006年狄佳春等報(bào)道了陸地棉亞紅株突變體,并對其光合特性進(jìn)行了初步研究[49]。王忠義等于2007年育成紅葉棉花品種紅雜111,霜前皮棉比DP99B增產(chǎn)11.3%[50]。萬艷霞等利用3個(gè)紅葉材料作父本,與15個(gè)綠葉棉花品種組配19個(gè)紅葉雜交組合,對具有紅葉標(biāo)記性狀棉花的產(chǎn)量雜種優(yōu)勢進(jìn)行研究發(fā)現(xiàn),19個(gè)紅葉雜交組合籽棉產(chǎn)量均具有明顯的雜種優(yōu)勢,皮棉產(chǎn)量具有正向中親優(yōu)勢、正向超親優(yōu)勢和負(fù)向競爭優(yōu)勢,衣分和鈴質(zhì)量具有正向的中親優(yōu)勢,單株鈴數(shù)表現(xiàn)正向中親、超親和競爭優(yōu)勢,并篩選出了4個(gè)產(chǎn)量競爭優(yōu)勢在5%以上、綜合性狀較好的優(yōu)勢組合[50]。陳旭升等選育出了具有亞紅葉標(biāo)記的優(yōu)質(zhì)高產(chǎn)彩色雜交棉組合蘇彩305,在生產(chǎn)上具有推廣潛力[51]。
為了克服胞質(zhì)雄性不育恢復(fù)系恢復(fù)基因鑒定難、保存難的問題,劉英新等選育了叢生鈴、雞腳葉、海島棉和無腺體4個(gè)標(biāo)記性狀恢復(fù)系,并進(jìn)行了產(chǎn)量、品質(zhì)和光合生理特性比較,發(fā)現(xiàn)它們在三系雜交棉育種、制種和繁種中都具有良好的利用價(jià)值[52]。
3?問題與展望
棉花形態(tài)標(biāo)記性狀在研究與應(yīng)用方面取得了一定成果,特別是在育種應(yīng)用研究方面成效顯著,已經(jīng)培育、選育出了一些具有標(biāo)記性狀的優(yōu)良種質(zhì)材料或品種,推動了棉花雜種優(yōu)勢在生產(chǎn)上的應(yīng)用。但目前形態(tài)標(biāo)記性狀的應(yīng)用還存在以下2個(gè)問題:
(1)形態(tài)標(biāo)記性狀利用率偏低。棉花具有豐富的形態(tài)標(biāo)記性狀,目前應(yīng)用比較成功的只有芽黃標(biāo)記、紅花標(biāo)記及腺體標(biāo)記,而大部分處于未開發(fā)狀態(tài)。因此,需要對形態(tài)標(biāo)記進(jìn)行進(jìn)一步的深入發(fā)掘,同時(shí)修改現(xiàn)行部分試驗(yàn)方法,促進(jìn)形態(tài)標(biāo)記的利用。(2)針對標(biāo)記性狀的基礎(chǔ)研究相對薄弱。相對于應(yīng)用研究,針對棉花形態(tài)標(biāo)記性狀遺傳特性解析及相關(guān)基因功能分析方面的研究較少,而且不夠深入,這也是阻礙形態(tài)標(biāo)記性狀在棉花育種中應(yīng)用的因素。隨著棉花基因組序列的公布,這一問題已逐步得到解決,受到廣泛關(guān)注的如色素腺體性狀標(biāo)記的遺傳基礎(chǔ)研究已經(jīng)有了一定突破,但其他性狀標(biāo)記的分子機(jī)制和遺傳研究都還比較滯后,有待進(jìn)一步深入挖掘。
基因工程和生物技術(shù)的不斷發(fā)展,勢必會推進(jìn)棉花形態(tài)性狀標(biāo)記的基礎(chǔ)和應(yīng)用研究更加深入,促進(jìn)棉花產(chǎn)業(yè)與其他產(chǎn)業(yè)的結(jié)合,不斷簡化生產(chǎn)成本,提高植棉收益,帶動植棉積極性,進(jìn)而有利于我國棉花產(chǎn)業(yè)的可持續(xù)發(fā)展。
參考文獻(xiàn):
[1]承泓良. 棉花質(zhì)量性狀遺傳研究進(jìn)展[J]. 江蘇農(nóng)業(yè)科學(xué),1984(7):46-49.
[2]肖松華,張?zhí)煺妫思荫x. 陸地棉芽黃近等基因系研究[J]. 棉花學(xué)報(bào),1996,8(5):235-240.
[3]肖松華,黃駿麒,潘家駒,等. 陸地棉芽黃品系和常規(guī)品種間雜種優(yōu)勢利用研究[J]. 棉花學(xué)報(bào),1996,8(2):71-76.
[4]閡留芳,何金龍,肖松華,等. 陸地棉芽黃品系在棉花雜種優(yōu)勢上的利用研究[J]. 棉花學(xué)報(bào),1996,8(3):113-119.
[5]王學(xué)德,潘家駒. 陸地棉芽黃指示性狀的雜種優(yōu)勢利用研究[J]. 南京農(nóng)業(yè)大學(xué)學(xué)報(bào),1989,12(1):1-8.
[6]潘家駒,閔留芳,劉?康,等. 陸地棉芽黃基因應(yīng)用于雜種棉的研究[J]. 南京農(nóng)業(yè)大學(xué)學(xué)報(bào),1998,21(3):7-14.
[7]張?zhí)煺妫思荫x,馮福禎. 一個(gè)有芽黃標(biāo)記性狀的棉花雄性不育系的遺傳鑒定[J]. 中國農(nóng)業(yè)科學(xué),1989,22(4):17-21.
[8]宇文璞,宇文綱,王淑霞,等. 溫敏型隱性核不育棉花應(yīng)用研究初報(bào)[J]. 河北農(nóng)業(yè)科學(xué),1994(4):38,24.
[9]宇文璞,宇文綱,王淑霞,等. 棉花溫敏型核不育芽黃A生產(chǎn)利用[J]. 中國棉花,1996,23(9):28.
[10]余筱南,陳金湘,李瑞蓮,等. 棉花溫敏雄性不育系的選育與應(yīng)用研究簡報(bào)[J]. 棉花學(xué)報(bào),2003,15(6):380-381.
[11]唐海明,余筱南,陳金湘,等. 棉花溫敏雄性不育系特棉S-1的利用[J]. 湖南農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版),2005,31(5):30-33.
[12]王凱輝,郭寶生,劉素恩,等. 棉花光溫敏核雄性不育系育性及其雜種優(yōu)勢研究[J]. 河北農(nóng)業(yè)科學(xué),2013,17(3):55-59.
[13]宇文小崗,王淑霞,宇文璞,等. 棉花雄性核不育系光溫A系簡介[J]. 中國棉花,2006,33(11):18.
[14]張建豐,張戰(zhàn)奇,宇文璞. 棉花單隱性光溫敏細(xì)胞核雄性不育系芽黃396A[J]. 農(nóng)業(yè)科技通訊,2006(4):33.
[15]牛金生,韓光明,藍(lán)家樣,等. 棉花光溫敏核不育系396A育性特征及雜種優(yōu)勢分析[J]. 棉花科學(xué),2016,38(6):7-12.
[16]張?濤,孟慶忠,王玉美,等. 棉花光溫敏雄性核不育系雜交成鈴率的影響因素研究[J]. 農(nóng)村經(jīng)濟(jì)與科技,2017,28(增刊1):23-24.
[17]朱?偉. 雞腳葉標(biāo)記的三系雜交棉雜種優(yōu)勢機(jī)理的研究[D]. 杭州:浙江大學(xué),2006.
[18]吳巧娟,肖松華,劉劍光. 雞腳葉對陸地棉光合特性的影響[J]. 江西棉花,2007,29(5):11-13.
[19]杜明偉,馮國藝,姚炎帝,等. 雜交棉標(biāo)雜A1和石雜2號超高產(chǎn)冠層特性及其與群體光合生產(chǎn)的關(guān)系[J]. 作物學(xué)報(bào),2009,35(6):1068-1077.
[20]馮國藝,羅宏海,姚炎帝,等. 新疆超高產(chǎn)棉花葉、鈴空間分布及與群體光合生產(chǎn)的關(guān)系[J]. 中國農(nóng)業(yè)科學(xué),2012,45(13):2607-2617.
[21]Andries J A,Jones J E,Sloane L W,et al. Effects of okra leaf shape on boll rot,yield,and other important characters of upland cotton,Gossypium hirsutum L.[J]. Crop Science,1969,9(6):705-710.
[22]Karami E,Weaver J B J. Dry-matter production,yield,photosynthesis,chlorophyll content and specific leaf weight of cotton in relation to leaf shape and colour[J]. Journal of Agricultural Science,1980,94(2):281-286.
[23]Baker D N,Myhre D L. Effects of leaf shape and boundary layer thickness on photosynthesis in cotton(Gossypium hirsutum)[J]. Physiologia Plantarum,1969,22(5):1043-1049.
[24]Karami E,Krieg D R,Quisenberry J E. Water relations andcarbon-14 assimilation of cotton with different leaf morphology[J]. Crop Science,1980,20(4):421-426.
[25]王淑民. 澳大利亞棉業(yè)[M]. 北京:中國農(nóng)業(yè)出版社,1995.
[26]王忠義,蘆立婷,樊建英. 超雞腳葉棉花資源育種潛力研究[J]. 中國棉花,2009,36(9):25-26.
[27]張雪林,韓世杰,彭凡嘉,等. 雞腳葉陸地棉育種研究進(jìn)展[J]. 中國棉花,2016,43(10):17-20.
[28]王忠義,趙敬霞,王興三,等. 超雞腳葉類型棉花的利用價(jià)值[J]. 中國棉花,2007,34(11):30-31.
[29]王忠義. 標(biāo)雜A1和標(biāo)記雜交棉一號的選育體會[J]. 中國棉花,2005,32(6):38-39.
[30]張家憲,蔣國柱,劉?偉. 高產(chǎn)優(yōu)質(zhì)短季低酚雜交棉——淮雜2號[J]. 中國棉花,2002,31(4):19-20.
[31]何鑒星,姜茹琴,張欣雪,等. 比克氏棉漸滲到陸地棉的花色基因的遺傳分析[J]. 遺傳學(xué)報(bào),2000,27(4):344-350.
[32]趙軍勝,高明偉,張興居,等. HB紅花性狀對陸地棉雜種優(yōu)勢的影響[J]. 山東農(nóng)業(yè)科學(xué),2011(3):11-14.
[33]趙軍勝,王秀麗,高明偉,等. 紅花性狀標(biāo)記雜交棉新品種魯05H9 SSR指紋圖譜構(gòu)建及應(yīng)用[J]. 山東大學(xué)學(xué)報(bào)(理學(xué)版),2014,49(1):44-49.
[34]王家寶,趙軍勝,高明偉,等. 紅花標(biāo)記雜交棉魯8H29的選育及配套栽培技術(shù)[J]. 中國棉花,2015,42(12):33-34.
[35]錢玉源,張海娜,崔淑芳,等. 棉花色素腺體的研究進(jìn)展[J]. 作物研究,2017,31(2):205-210.
[36]袁有祿,靖深蓉,張?zhí)煺妫? 陸地棉顯性無腺體指示性狀的雜種優(yōu)勢利用研究[J]. 種子,1997(6):15-17.
[37]權(quán)月偉,李繼軍,唐光雷,等. 河北省低酚棉育種研究進(jìn)展及應(yīng)用前景[C]//2015全國棉花青年學(xué)術(shù)研討會論文集. 新疆石河子:中國棉花雜志社,2015.
[38]喻樹迅,原日紅,余學(xué)科,等. 低酚棉中棉所20遺傳特異性與豐產(chǎn)性機(jī)理的研究[J]. 中國農(nóng)業(yè)科學(xué),1999,32(5):16-22.
[39]祝水金,蔣玉蓉,Reddy N,等. 陸地棉子葉色素腺體延緩形成種質(zhì)系的育成及其遺傳研究[J]. 科學(xué)通報(bào),2004,49(19):1987-1992.
[40]陳?瑩,趙軍勝,高明偉,等. 腺體標(biāo)記棉花雜交種魯棉研39號指紋圖譜構(gòu)建[C]//中國棉花學(xué)會2014年年會論文集. 呼和浩特:中國棉花雜志社,2014.
[41]翟雷霞,米換房,李繼軍,等. 抗蟲低酚棉品種邯無216的選育及栽培技術(shù)要點(diǎn)[J]. 中國棉花,2016,43(7):33-34.
[42]劉?芳,譚忠寧,比買熱木·阿不都艾海提. 47個(gè)低酚棉品種資源性狀觀察與分析[J]. 中國棉花,2015,42(7):15-17.
[43]李函利,王省芬,王國寧,等. 低酚棉種質(zhì)資源耐鹽性鑒定[J]. 植物遺傳資源學(xué)報(bào),2016,17(6):1099-1105.
[44]劉?慧,季靈艷,趙天倫,等. 陸地棉色素腺體與不同棉酚旋光體含量之間的相關(guān)性研究[J]. 棉花學(xué)報(bào),2017,29(5):437-446.
[45]張金發(fā),孫濟(jì)中,劉金蘭. 棉花窄卷苞葉基因?qū)Ξa(chǎn)量品質(zhì)的影響[J]. 作物學(xué)報(bào),1993,19(4):315-320.
[46]張金發(fā),孫濟(jì)中,劉金蘭. 窄卷苞葉基因?qū)γ藁ü夂闲阅艿挠绊慬J]. 華中農(nóng)業(yè)大學(xué)學(xué)報(bào),1993,12(4):328-332.
[47]張金發(fā),孫濟(jì)中,劉金蘭. 窄卷苞葉基因?qū)γ藁ㄔ缡煨缘挠绊慬J]. 華中農(nóng)業(yè)大學(xué)學(xué)報(bào),1991,10(4):321-326.
[48]李俊蘭,崔淑芳,韓擇林. 棉花新類型——觀賞棉花[J]. 中國棉花,2003,30(9):29-51.
[49]狄佳春,殷劍美,陳旭升,等. 陸地棉亞紅株突變體光合特性初步研究[J]. 江蘇農(nóng)業(yè)科學(xué),2006(2):33-34.
[50]萬艷霞,馬峙英,王國印,等. 紅葉棉花的產(chǎn)量雜種優(yōu)勢研究初探[J]. 棉花學(xué)報(bào),2010,22(2):186-188.
[51]陳旭升,狄佳春,趙?亮. 具有亞紅葉標(biāo)記的抗蟲彩色雜交棉選育[J]. 江西農(nóng)業(yè)學(xué)報(bào),2015,27(4):1-3.
[52]劉英新,王學(xué)德,倪?密,等. 四個(gè)帶有標(biāo)記性狀的棉花CMS恢復(fù)系的主要農(nóng)藝與光合生理特性[J]. 棉花學(xué)報(bào),2010,22(5):448-453.
