噬菌體在食品應用中的安全性
2019-11-28 03:40:04叢聰袁玉玉王麗麗李曉宇李淑英徐永平
國外醫藥(抗生素分冊) 2019年5期
關鍵詞:李斯特
叢聰,袁玉玉,王麗麗,2,4,李曉宇,2,4,李淑英,徐永平,,*
(1 大連理工大學生物工程學院, 遼寧大連 116024;2 動物性食品安全保障技術教育部工程研究中心, 遼寧大連 116600;3 大連賽姆生物工程技術公司, 遼寧大連 116620;4 遼寧省大連賽姆噬菌體應用工程技術研究中心,遼寧大連 116600)
1 前言
近幾年來,每每發生由食源性病原菌引起的各種疾病,尤其是食物中毒和腸道傳染病。由于全球性食品貿易的快速增長,導致了飲食習慣、食品加工方式的變化,食源性致病菌不斷出現以及細菌耐藥等一系列問題,這些均成為人類必須面對的嚴峻挑戰[1]。食品安全問題備受關注,由于食品生產技術中不可能完全殺滅食物中的病原菌,因此國內外一直在研究并完善殺滅食源性致病菌的話題[2]。人們逐漸開始接受將噬菌體(Bacteriophage,簡稱phage)應用于食品工業中控制食源性致病菌,這不僅能保障食品的安全性、延長食品保質期,同時也促進了人們對噬菌體自身安全性領域的研究。
通常在食品生產中用于控制食源性致病菌的方法包括物理(蒸汽、干熱和紫外線照射)和化學(殺菌劑、防腐劑)兩種處理方法,采用這兩種方法處理的食品感官品質會受到影響,且因不屬于綠色食品而使消費者產生排斥心理[3]。熱殺滅技術屬于傳統的物理殺菌方法,但因加熱溫度過高,食品的色澤、風味會發生明顯變化,營養價值也大幅降低;冷殺菌技術在包裝打開以后,又有再次染菌而腐敗的可能[4]。另外,某些抑制食源性致病菌的手段不能直接應用于新鮮水果蔬菜和即食食品中。目前常見的食源性致病菌包括沙門菌(Salmonella)、大腸埃希菌(Escherichia coli) O157:H7、單增李斯特菌(Listeria monocytogenes)、金黃色葡萄球菌(Staphylococcus aureus)和彎曲桿菌(Campylobacter)等。
因此人們開始使用噬菌體作為食品中的天然抗菌劑來殺滅或抑制食源性致病菌,從而保證食品安全[5-7]。本文旨在探討噬菌體安全性的問題、運用噬菌體控制食品中致病菌的理論依據,以及對致病菌進行控制的研究現狀。
2 噬菌體概述
噬菌體是一種能夠感染細菌、放線菌或螺旋體等微生物的病毒。它不具有感染哺乳動物細胞的能力,僅能特異性地侵染其宿主菌。因其具有嚴格的宿主特異性特點,一種噬菌體往往只能感染和裂解某種細菌,甚至只裂解種內的某些菌株[5]。噬菌體只寄居在易感的細胞宿主內,它可以在脫離寄主細胞的狀態下維持一段時間的活性,若一旦脫離了宿主細胞,既不能生長也不能復制[8]。噬菌體主要有兩種類型——裂解性噬菌體(virulent phage)和溶原性噬菌體(溫和噬菌體,temperate phage)。溶原性噬菌體一般不引起宿主菌的裂解[9],而應用于食品工業的噬菌體是裂解性噬菌體,它在侵染宿主菌后會引起宿主菌裂解,同時釋放子代噬菌體,進而達到殺菌作用。
大量研究和應用實例已經表明,利用噬菌體對食品中的致病菌進行控制是一個可行、高效,且有著廣闊發展空間的方法[10]。噬菌體作為細菌的天敵被用于食品上時,由于其高度特異性而只會殺滅目標宿主菌,不會對食品的固有菌群產生影晌,這正是食品抗菌劑所需要的品質,它有助于維持食品特別是發酵食品的產品品質[11]。因此,從環境中篩選出能特異性侵染食物中病原菌的噬菌體,并利用它們對食品中的致病菌進行控制,這對改善食品的品質、提高食品安全性等方面具有重大意義。
3 噬菌體在食品工業中的應用
存在于食品原輔料采集、生產、加工、儲藏等環節的食源性致病菌會引發食品污染,造成食品腐敗變質并傳播疾病,噬菌體可以通過殺滅食源性致病菌來保證食品安全。
3.1 原料采集時殺滅病原菌
宰殺動物時,有害細菌會隨血液、糞便污染尸體,因而可在屠宰后用噬菌體對動物尸體進行消毒;也可在原料采集(擠奶或屠宰)前,通過給動物口服噬菌體的方法殺滅其體內病原菌。例如,Bach等[12]發現在人工瘤胃中的噬菌體DC22可有效清除E.coliO157:H7,而得了瘋羔病的羊口服噬菌體制劑后卻不能減少其糞便中細菌的含量,這可能是因為沒有足夠的噬菌體到達羊腸道所導致。Raya等[13]給羊口服噬菌體CEV1來殺滅E. coliO157:H7,服用2 d后羊腸內的E. coliO157:H7的數量明顯降低。
3.2 生產及加工環節的消毒
噬菌體可對生產環境(工作表面、地面、墻壁、加工設備等)進行凈化和消毒。例如,布盧維爾等[14]用噬菌體混合物處理被阪崎腸桿菌(Enterobacter Sakazakii)污染的不銹鋼盤表面后,沒有再發現該菌,而未被處理的對照組仍有該菌存活甚至可以輕微生長。Viazis等[15]用噬菌體混合物BEC8處理3種不同材質工作表面上的E. coliO157:H7,室溫1 h后噬菌體能起到有效的殺菌效果。
在食品生產中可直接將噬菌體與食物混合使用。O'Flynn等[16]用3種不同噬菌體來處理被103CFU/gE. coliO157:H7污染的牛肉,在37 ℃儲存時,78 %的牛肉未發現有大腸埃希菌,而剩余的22 %的牛肉中大腸埃希菌的數量都低于10 CFU/g。Goode等[17]發現將噬菌體應用在食品中可以減少95 %的空腸彎曲桿菌,同時還發現了能夠殺滅和抑制沙門菌的噬菌體,并且即使是沙門菌的抗性菌株在高濃度的噬菌體作用下也會被殺滅,兩個或兩個以上的噬菌體可直接殺死被侵染的宿主菌。Higgins等[18]成功用噬菌體殺滅了禽肉產品上的腸炎沙門菌,Humberto等[19]對雞肉表面也做過類似的實驗。
3.3 原料消毒并延長食品儲藏期
噬菌體作為天然的抗菌劑和防腐劑可直接對新鮮的水果蔬菜進行消毒。例如沙門菌是引起人類食物中毒的主要致病菌之一,在實驗中用噬菌體混合物對已感染沙門菌的哈密瓜和蘋果進行噬菌體處理后,哈密瓜上的沙門菌明顯減少;然而,蘋果片上的沙門菌似乎未受噬菌體影響,可能由于蘋果片上的pH較低導致噬菌體數量快速減少造成的[20]。
噬菌體還能延長食品的保質期,例如阪崎腸桿菌能引起新生兒腦膜炎,主要通過嬰兒配方奶粉進行感染。由于目前生產奶粉時不能做到完全無菌,所以當奶粉在室溫存放沒有被及時食用時,少量致病菌大量繁殖,最終會對嬰幼兒健康構成潛在危險。為了可以安全儲存嬰兒配方奶粉,雀巢技術公司分離出了對阪崎腸桿菌具有強裂解力的無毒噬菌體ESP 732-1,用以防止阪崎腸桿菌污染,這種T4噬菌體能在24 ℃~37 ℃有效抑制該菌的生長,其濃度與抑菌效果成正比,當濃度達到109PFU/mL時能夠完全殺滅阪崎腸桿菌[14]。Whichard等[21]用宿主譜廣的沙門菌噬菌體Felix-O1對香腸中的鼠傷寒沙門菌(Salmonella typhimurium)進行侵染,結果發現鼠傷寒沙門菌的數量明顯減少。另有研究發現,在干酪生產的發酵液中人為地加入沙門菌及其噬菌體SJ2,儲存數月后,沙門菌依然不能生長[22]。
3.4 檢測食源性致病菌
噬菌體的宿主特異性在食品工業中可用于檢測食源性致病菌,如噬菌體擴增法檢測技術:在待檢測食品中加入某種噬菌體,若其宿主菌存在于食品內則會被噬菌體迅速侵染,隨后用特異性高的殺病毒劑清除所有未侵染的噬菌體,已侵染的噬菌體則繼續擴增,進而裂解細胞,釋放子代噬菌體繼續侵染周邊宿主細胞,最終在培養皿中展現出噬菌斑,此種方法可證明食品中存在食源性致病菌。其他檢測食源性致病菌的方法還有熒光染料標記法、檢測報告基因法等[23]。
4 已開發的食品級噬菌體制劑產品
在國外,很多生物技術公司對噬菌體的研發非常重視,目前已有多家公司參與研制并生產噬菌體成品制劑,部分公司信息詳見表1。噬菌體在食品上用來對食源性致病菌進行生物防控的研究取得了很大進展,市場上已有一些噬菌體的產品被商業使用,并已通過美國食品藥品管理局(Food and Drug Administration, FDA)認證后上市,有9種已通過GRAS (Generally Regarded As Safe,公認安全)認證級別,還有2種待批準為GRAS產品。
2006年,FDA批準李斯特菌噬菌體制劑Listex?P100在即食食品和禽類產品中使用,它由荷蘭EBI食品安全公司(EBI Food Safety)研發,可有效避免肉類及奶酪類產品中李斯特菌的污染,并且對人體無害,這是世界上第一個通過FDA認證的用于食品工業的產品[24]。這個產品標志著FDA首次將噬菌體視為了GRAS級添加劑,隨后獲得了歐洲、澳大利亞和新西蘭的批準應用于食品中[25],之后不斷有針對不同致病菌的噬菌體產品通過FDA GRAS的認證并獲得應用批準,所有GRAS級噬菌體制劑詳見表2,部分抑制食源性致病菌的噬菌體研究詳見表3。
在食品工業中,噬菌體自身的抗微生物性質可減少人們對抗生素的依賴和利用,并能顯著降低制藥公司開發新藥的數量,降低開發總成本,更容易滿足消費者對不含病原體和合成化學品食物的需求。另外,由OmniLytics公司生產的噬菌體制劑Agriphage?是第一個由美國環保署(US EPA)正式批準用于農業的噬菌體產品,主要用來治療農作物的番茄細菌斑病[51]。
5 噬菌體的特征
5.1 宿主特異性
噬菌體的宿主特異性可以保證其在裂解宿主菌的同時不殺死食品生產中的發酵菌株,因此在使用噬菌體對生產設備等進行消毒時,不用擔心噬菌體會影響發酵生產。此外,噬菌體僅能侵染并裂解細菌細胞,而不會侵入或殺死哺乳動物的細胞,因此對人體無害。
5.2 增殖能力強
一個適當條件的裂解周期內,每個噬菌體可產生約200個子代噬菌體,這表明它可以以200n的速度進行增殖。經過不斷地復制和增殖,進而裂解大量的宿主細胞,因此可快速地抑制并殺滅食源性致病菌。

表1 部分噬菌體制劑研發公司
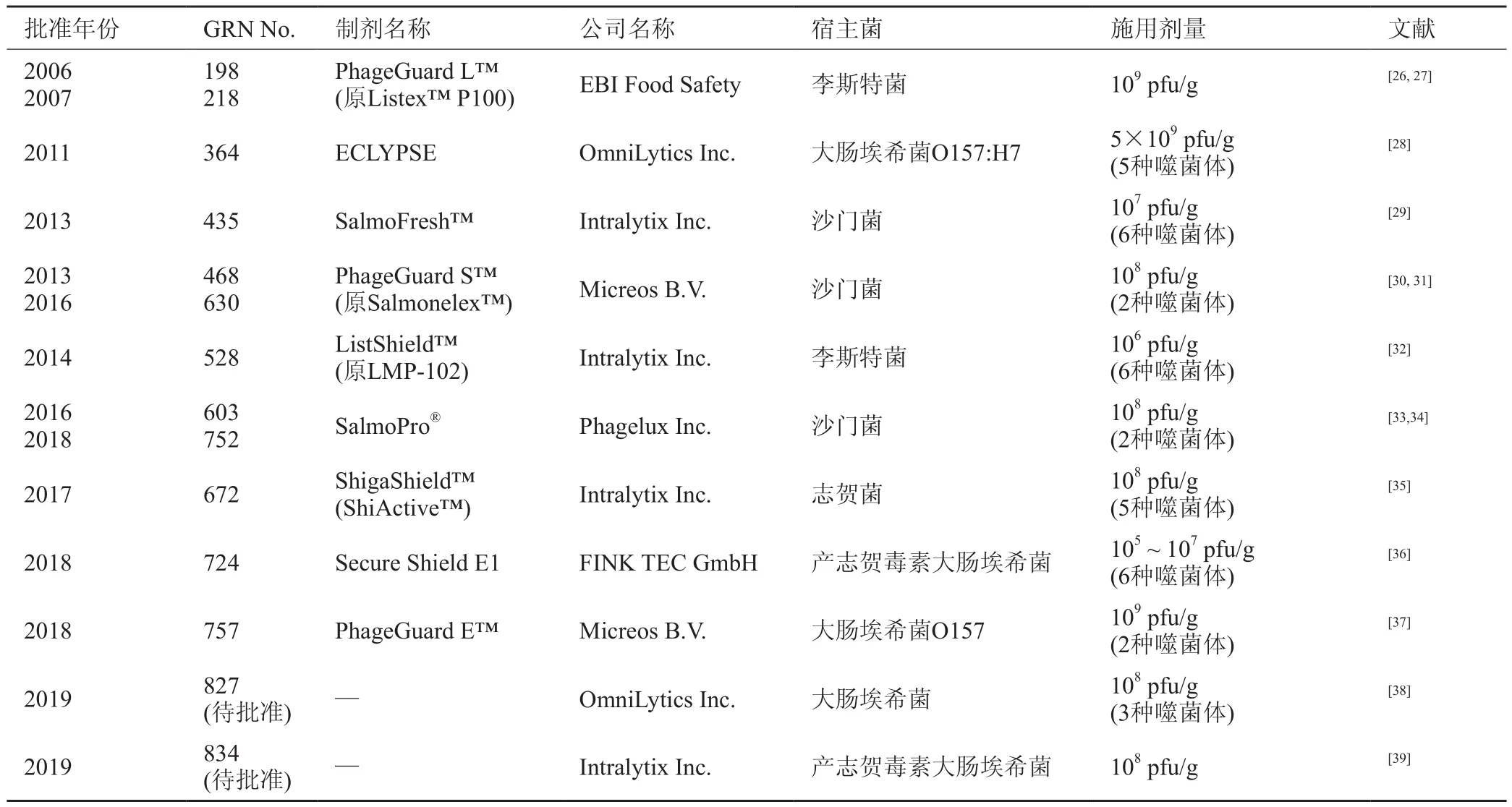
表2 所有FDA批準的GRAS級噬菌體制劑

表3 部分食品級研究所用噬菌體
5.3 安全無毒副作用
5.3.1 自然界中廣泛存在
自然環境中含有高豐度的噬菌體,數量大、種類多。干凈的水中噬菌體含量為109PFU/mL,表層海水含量為107PFU/mL,陸生系統也含有大量的噬菌體[52]。同樣地,食物中也有大量的噬菌體。研究人員從泡菜中分離出了26種不同的噬菌體[53]。每克新鮮的肉類、果蔬以及熟食中含有104個E. coli噬菌體[54]。很明顯,我們每天通過飲食、飲水甚至呼吸都會攝入大量的噬菌體。
5.3.2 對腸道菌群無害
噬菌體僅專一性地尋找對應的宿主菌,而對腸道內其他的細菌無害,因此不會干擾體內正常的菌群及代謝,并且生物機體自身也能通過免疫系統清除體內的噬菌體。研究人員用E. coli噬菌體做了安全性研究,給小鼠及人類志愿者直接喂食大劑量的E. coli噬菌體,這些噬菌體能夠在體外感染E. coli菌株,結果顯示該噬菌體無論對小鼠還是人類的機體都沒有明顯影響[55,56],且對消化道內的微生物菌群也沒有影響,可能是由于E. coli菌群生活在噬菌體不容易到達的位置,也可能是機體的免疫系統清除了體內的噬菌體,使之減少到不能殺滅細菌的水平[57]。
噬菌體本身是由核酸和蛋白質構成,其成分對人體不具有毒害作用,而且也不會改變食物的顏色、結構和味道[58]。活性噬菌體被機體攝入后通過消化分解,產生出核酸和蛋白質,若再進一步可分解為磷酸、堿基、氨基酸等,均是無毒的代謝產物。因此,有活性的噬菌體僅侵染宿主菌,失去活性的噬菌體也不會對機體產生任何毒害作用。
5.3.3 有長期使用的歷史
自從1919年Felix d'Herelle第一次采用噬菌體完成了對痢疾的治療后,東歐和前蘇聯就一直研究和生產用于治療疾病的噬菌體制劑[59]。噬菌體的給藥途徑大部分是口服和表面擦藥,也有肌肉注射、靜脈注射甚至是頸動脈注射[60],但并沒有關于噬菌體對機體產生不良影響的報道。
García等[61]以金黃色葡萄球菌的溫和噬菌體ΦH5和ΦA72為試驗材料,通過DNA隨機缺失法得到與兩株溫和噬菌體ΦH5和ΦA72相對應的烈性噬菌體,并將這兩株烈性噬菌體用于干酪的制作中,有效地抑制了干酪中的金黃色葡萄球菌。Edita等[62]用兩種噬菌體混合物來滅活新鮮和硬型奶酪中的金黃色葡萄球菌,結果發現噬菌體的加入使宿主菌顯著減少,并且沒有影響發酵劑菌群結構,同時也維持了奶酪預期的物理化學特性。Pasternack和Sulakvelidze[63]申請了6個單增李斯特菌噬菌體的專利,這些噬菌體能夠有效控制由單增李斯特菌引起的食物污染。美國Omnilytics公司銷售的噬菌體混合制劑用在溫室和田間已有10多年的歷史,已證明該產品對細菌斑點感染病的治療是安全及有效的,已被美國EPA批準使用。在歐洲,EBI食品安全委員會授權認同芬蘭一家公司生產的噬菌體產品LISTEX?是安全的[64]。
所以通過以上的證據我們可以得知,吃到以噬菌體作為食品添加劑的食物是不需要擔驚受怕的,因為噬菌體是“可以被吃的”。
6 噬菌體對食源性致病菌的裂解機制
已有文獻報道過噬菌體侵染及裂解宿主菌的機制,使用噬菌體控制目標菌主要有兩種方式:
(1)直接用分離出的噬菌體控制目標菌,如Carlton等[24]研究了單增李斯特菌的噬菌體P100,全基因組測序后給小鼠服用大劑量的噬菌體P100,口服獨立性試驗驗證應用噬菌體P100是安全可靠的,小鼠沒有表現出副作用;同時,他發現用噬菌體P100能夠完全清除軟干酪中的李斯特菌屬,并且在食物中P100和A511都能殺滅李斯特菌,因此他認為該噬菌體可以安全有效地抑制李斯特菌[24]。Guenther等[65]報道單增李斯特菌的噬菌體A511在肉制品和乳制品中對單增李斯特菌有一定的控制作用。Abuladze等[40]將三種噬菌體的混合物作用于污染了大腸埃希菌O157:H7的食品中,結果發現噬菌體能夠顯著降低食品中大腸埃希菌O157:H7的量。Modi等[22]發現在制作Cheddar干酪之前向原料奶中添加噬菌體SJ2能有效抑制沙門菌屬,在干酪99 d的儲存期內能使沙門菌濃度保持在50 CFU/g以下,而不加噬菌體SJ2的干酪在99 d的儲存期內沙門菌的濃度為103 CFU/g。Carvalho等[66]成功分離出了43株噬菌體,它們能有效抑制結腸彎曲桿菌(Campylobacter coli)和空腸彎曲桿菌(Campylobacter jejuni),從而為保障食品安全提供可能。Bigwood等[43]將噬菌體Cj6接種于污染了空腸彎曲菌的牛肉中,結果顯示空腸彎曲菌的數量有一定程度的減少,同時他將沙門菌噬菌體P7接種于污染了沙門菌的牛肉中,結果發現沙門菌數量顯著降低,當噬菌體的侵染數量越多,沙門菌的減少量越多。
(2)在分離得到目標菌的噬菌體后,通過分子生物學手段獲得噬菌體的裂解酶,利用裂解酶來控制目標菌的效果尤為顯著。已發現噬菌體K能夠抑制葡萄球菌的活性,其內溶素Lys K已被克隆并用乳酸乳球菌表達。人們發現Lys K可以殺死9種葡萄球菌,其中包括耐甲氧西林金黃色葡萄球菌(MRSA)[67]。Donovan等[68]將金黃色葡萄球菌噬菌體phi11應用于牛奶中,其裂解酶對金黃色葡萄球菌和其他6種凝固酶陰性的葡萄球菌均有裂解作用,這說明該酶可直接用于牛奶中控制致病菌。Celia等[69]將產溶菌素的基因重組到質粒上后轉入到大腸埃希菌中表達,純化出溶菌素Ply700后對其性質進行了測定,Ply700對有乳鏈球菌(Streptococcus uberis)、釀膿鏈球菌(Streptococcus pyogenes)和停乳鏈球菌(Streptococcus dysgalactiae)均能裂解。Obeso等[70]用大腸埃希菌表達并純化了金黃色葡萄球菌噬菌體ΦH5的裂解酶LysH5,將其用于巴氏消毒奶中,結果表明未在樣品中檢測到金黃色葡萄球菌。Loessner等[71]將單增李斯特菌噬菌體A118編碼的噬菌體內溶素Ply118用于提取DNA、RNA和破碎單增李斯特菌的細胞壁上;乳酸乳球菌中的乳球菌素啟動子啟動并表達出克隆的Ply118和Ply511,它們可以有效抑制單增李斯特菌的活性。Hu等[72]對銅綠假單胞菌(Pseudomonas aeruginosa)噬菌體PaP3的裂解酶進行了鑒定分析,結果表明噬菌體PaP3的裂解酶對金黃色葡萄球菌具有顯著的抑制作用,但對銅綠假單胞菌和大腸埃希菌無抑制作用。
7 噬菌體的潛在風險
以噬菌體為基礎的生物防控手段在提高食品微生物安全方面的應用具有很大的潛力,因為它們安全使用的歷史悠久、操作處理相對簡單,并且其抗菌活性具有高度特異性。噬菌體的安全使用主要體現在:噬菌體在自然界中無處不在,能在動物和人體中自然共生,包括在食品生態系統(Food Ecosystems)中也是,而且在東歐地區臨床上已被廣泛應用。噬菌體的高特異性表現在:對哺乳動物細胞無害,對腸道菌群沒有不利的影響,是其它高效抗菌劑的來源。但不可否認的是,任何事物都有其弊端,噬菌體也具有相應的潛在風險。
7.1 噬菌體的溶原現象
不是所有的噬菌體都適于殺滅細菌病原體。溶原性噬菌體不會殺死宿主菌,而是將其基因組整合到宿主染色體上(溶原現象),并能使宿主菌抵抗其它噬菌體的溶菌作用[9];另外,整合的基因組可以改變宿主的表型,甚至能夠增強宿主菌的致病性或毒力。例如霍亂弧菌(Vibrio cholerae)的霍亂毒素CTX是由整合的噬菌體CTX的ctxA和ctxB兩個基因所編碼的[73];又如由E. coli的志賀樣毒素是由其整合基因組中的溶原性噬菌體基因stx1和stx2編碼的[74]。
7.2 噬菌體的轉導作用
轉導是以噬菌體為媒介,把供體菌的小片段DNA攜帶到受體菌中,經過交換及整合使受體菌獲得供體菌的部分遺傳性狀的現象,這個過程很有可能會把新的基因傳遞給受體菌[75]。如果噬菌體在致病菌(供體菌)中增殖后攜帶了致病基因,則很有可能把受體菌也轉變成有致病性的致病菌[76]。因此,若不得已使用致病性宿主來增殖噬菌體,則應選用不具有轉導作用的噬菌體。研究發現,在李斯特菌和梭狀芽孢桿菌(Clostridium Prazmowski)的噬菌體中,有明確固定基因組末端的噬菌體是不具有轉導作用的,而含有末端冗余并有環狀排列基因次序的噬菌體是能夠進行轉導作用的[77,78]。
7.3 出現抗噬菌體菌株
細菌可自發突變產生抗噬菌體的菌株(抗性菌)。例如Park等[79]研究變形假單胞菌(Pseudomonas plecoglossicida)時發現了抗噬菌體的菌株。大部分細菌的突變率為10?6,一般不會產生大量的抗噬菌體菌株,因此對噬菌體的殺菌效果不會產生明顯影響。但若長期多次使用該噬菌體,在自然選擇的影響下,只有抗性菌菌株能夠存活下來并不斷增殖,這時該種噬菌體就很難對抗性菌進行殺滅,只能尋找方法與細菌共進化產生能殺滅抗性菌的噬菌體。
8 總結
人們最早用噬菌體來治療細菌性疾病,后來逐漸將其應用在食品工業上,目前已篩選出可用來抑制阪崎腸桿菌、E. coliO157:H7、傷寒沙門菌等食源性致病菌的噬菌體。噬菌體制劑Listex? P100是世界上第一個通過FDA的GRAS級認證產品,并被批準作為食品生產中的加工助劑使用。利用噬菌體對食品中的致病菌進行控制是一種高效、快速、發展前景廣的生物防控手段,在研究和應用領域中均值得進一步地探索。噬菌體的裂解譜較窄使其應用受到了限制,因此我們需要篩選出更多的廣譜噬菌體。另外,噬菌體應用的安全性也將是以后持續關注的重點。針對目前國內外頻發的由食源性致病菌引起的食品安全問題,特異性噬菌體的研究與開發勢必成為未來的趨勢和熱點,這些都預示著噬菌體在食品安全中的應用前景非常廣闊。
但我們還應留意噬菌體的溶原現象和轉導作用等的潛在風險,它們會改變宿主菌的表型,具有增強宿主菌的致病性或毒力的可能。因此我們在研究及使用噬菌體時,應避免使用溶原性噬菌體和具有轉導作用的噬菌體。
對于某些食品,噬菌體不能對病原菌起到完全地殺滅或抑制作用[20],另外細菌若自發突變可能會出現抗噬菌體菌株。因此若能將噬菌體制劑與其它保鮮防腐方法結合使用則可更有效地抑制病原菌,從而保障食品的安全性和穩定性。
猜你喜歡
藝術評鑒(2022年6期)2022-05-10 10:07:27
樂府新聲(2019年2期)2019-11-29 07:34:30
幼兒園(2019年7期)2019-09-05 17:49:18
北方音樂(2017年4期)2017-05-04 03:40:15
中國音樂教育(2015年8期)2015-05-16 03:22:06
小小說月刊(2013年6期)2013-05-14 14:55:19
食品科學(2013年23期)2013-03-11 18:30:07
食品科學(2013年23期)2013-03-11 18:30:05
食品科學(2013年15期)2013-03-11 18:25:40
食品科學(2013年15期)2013-03-11 18:25:25
