岷縣生荒地和熟地育成當歸苗抗逆生理特性的差異
2019-11-18 06:18:00白剛郭鳳霞陳垣袁洪超肖婉君
草業學報 2019年11期
白剛,郭鳳霞*,陳垣,2*,袁洪超,肖婉君
(1.甘肅農業大學生命科學技術學院,農學院,甘肅省中藥材規范化生產技術創新重點實驗室,甘肅省藥用植物栽培育種工程研究中心,甘肅省干旱生境作物學重點實驗室,甘肅 蘭州 730070;2.甘肅省特色藥用植物資源保護與利用工程實驗室,甘肅省特色藥材規范化可追溯栽培工程技術研究中心,甘肅中天藥業有限責任公司,甘肅 定西 748100)
當歸(Angelicasinensis)為傘形科當歸屬草本藥用植物,以干燥根入藥,具有補血活血等多種功效,是我國傳統大眾中藥材品種之一[1]。當歸適宜氣候冷涼,降水充沛的生態環境,并適宜排水良好,有機質含量較高的黑褐類土壤[2]。當歸野生資源已趨瀕危,全國第四次中藥資源普查僅在甘肅省岷縣和宕昌縣人跡罕至的高寒陰濕灌木林發現極其稀有的野生當歸資源[3]。現市售當歸主要為栽培品種,地處青藏高原東部邊緣地帶的甘肅省岷縣、漳縣、卓尼縣和宕昌縣一帶氣候高寒陰濕,土層深厚肥沃,降水充沛,地勢陡險,土壤不宜積水,屬于我國優質當歸道地產區,已有1500多年栽培史,年產量和銷量均占全國當歸的90%以上[4],2001年被中國特產之鄉推薦暨宣傳活動組織委員會命名為“中國當歸之鄉”。在限定使用范圍和劑量內當歸既是保健食品又是中藥材物質的中藥材品種。
當歸栽培期3年,第1年種子育苗,第2年種苗成藥栽培,第3年留株生產種子。當歸一旦進入生殖生長其肉質根逐漸木質化。早期抽薹也稱未熟抽薹,即當歸在成藥栽培期尚未成藥而植株提早通過春化作用而抽薹開花,造成藥材根木質化失去藥用價值的物候特征[5]。隨著中藥現代化和藥食同源的普及,當歸需求量增大,種植面積擴大。然而,適宜當歸育苗的生態區域卻極其狹窄,僅局限于甘肅省岷縣和漳縣高海拔草原區,育成種苗質量優異,根病率和早期抽薹率均低[6]。因此,長期以來供苗基地一直沿用生荒地育苗,不占耕地,是農民傳統的育苗方法。然而,隨著全球氣候變暖,當歸早期抽薹率提高。另外,重迎茬栽培導致當歸根病加重,死苗率高,嚴重影響栽培成效[7-11]。這些問題加重了藥農對生荒地育苗的主觀依賴性。生荒地育苗嚴重破壞了天然植被和草皮生態,極不利于保護野生物種的繁衍[12]。當歸道地產區生態環境建設與傳統育苗方式嚴重矛盾,可開墾山地幾乎用盡,育苗向更高海拔荒地延伸,生態環境日趨惡化[13],大面積泥石流災害頻繁,農田生態系統功能受損,恢復機制的研究顯得至關重要,探尋可替代生荒地當歸育苗的熟地農茬資源是需要迫切解決的關鍵科技問題。盡管有人利用岷縣熟土進行當歸設施育苗探索,發現早播搭網育苗有一定成苗率[7],熟地黃芪(Astragalusmembranaceus)茬土與生荒土盆栽育成當歸苗抗逆生化基礎相當[4,14],根際微生物多樣性相似[5]。宕昌縣日光溫室黃芪茬土也較利于當歸苗的生長發育[8],但設施育苗因成本高對當歸生地育苗現狀的改變成效甚微。近年來,張新慧等[15]調查發現,熟地農茬口影響當歸產量和質量。在作物茬口特性研究基礎上[16],梁偉等[17]首次在卓尼縣8種作物茬口熟地進行當歸育苗研究。岷縣熟地黃芪與小麥(Triticumaestivum)茬口育成當歸種苗抗病性較強[6]。上述研究說明當歸依賴生荒地育苗的傳統方式是有望通過科學研究改變的,但熟地育苗科學問題尚不明確,缺乏與生荒地育苗的系統比較。逆境是指對作物生長不利的各種綜合環境因素,抗逆植物具有發達的抗氧化系統[18-19],根病率、返青率和早期抽薹率均是反映當歸苗抗逆性強弱的主要指標,豌豆(Pisumsativum)具有勻田和培肥地力的雙重作用。因此,在岷縣系統研究生荒地和熟地豌豆-黃芪雙豆輪作茬育成當歸種苗抗逆特性及其差異性具有重要意義,可為篩選替代生荒地當歸育苗的熟地茬口資源提供科學依據,以改變生荒地育苗的傳統格局,促進當歸產業的可持續化生態發展。
1 材料與方法
1.1 試驗區概況及當歸種子來源
當歸育苗試驗設在甘肅省定西市岷縣禾馱鄉石家臺村大石溝大灣梁,屬高原型大陸性季風氣候,境內平均海拔2865 m,年均氣溫5.5 ℃,年均日照時長2214.9 h,年均降水量800 mm,無霜期123 d,最熱7月平均氣溫16 ℃,最冷1月平均氣溫-6.9 ℃,平均相對濕度68%,植被覆蓋良好,屬典型高寒陰濕區,高寒草甸土壤類型,為當歸傳統道地育苗區和栽培產區。
當歸生荒地(uncultivated fields, SH )和熟地黃芪茬口(cultivated residue ofA.membranaceus, HQ)育苗試驗地分別設在岷縣禾馱鄉石家臺村大石溝大灣梁山頂和半山腰,位于同一山脈,均屬溫帶大陸性季風氣候,但海拔高度、降水量和土壤及植被類型均有差異。山頂生荒地土壤類型為高寒草甸土壤,半山腰熟地農田土壤類型為灰棕壤,均適宜當歸生長發育。當歸育苗前對植被觀察,育苗期采用地溫自動記錄儀(RC-4HC Data Logger,成都華恒儀器有限公司)對生荒地和熟地試驗地中心地表10 cm 處溫度自動記錄(表1)。
育苗試驗用當歸種子為2016年在生荒地留苗地3年生當歸種株采集的種子,經甘肅農業大學陳垣教授鑒定為當歸,掛涼保藏備用于生荒地和熟地黃芪茬當歸育苗試驗。

表1 2016年9月18日至2017年9月17日岷縣生荒地與熟地當歸育苗田地表溫度比較Table 1 Comparison of annual soil surface temperatures in the uncultivated and cultivated fields for A. sinensis seedling cultivation in Min County, during September 18, 2017 to September 17, 2018
1.2 岷縣熟地豌豆-黃芪茬口培育及當歸育苗
在岷縣禾馱鄉石家臺村半山腰熟地農田2013-2014年當歸繁種結束,2015年種植豌豆品種隴豌2號勻田倒茬,2016年采用種子播種蒙古黃芪(A.membranaceusvar.mongholicus)育苗,培育黃芪茬口,以確保茬口土壤均勻一致。2017年當歸播種育苗。黃芪茬口培育試驗采用隨機區組設計,3次重復,即在上年豌豆勻田基礎上,2016年4月10日將試驗地按坡度劃分為3個區組(3次重復),小區面積4.8 m2(4.0 m×1.2 m),區組間隔0.5 m,四周設地埂,地埂寬0.5 m,每小區施用有機肥1 kg(施用量2083.3 kg·hm-2),采用開溝條播法播種蒙古黃芪種子,播深3 cm,行距10 cm,播種后耙平,以培育黃芪茬口,每小區種子播種量75 g(折合播量156.3 kg·hm-2),黃芪育苗期田間管理同大田。黃芪種苗采挖期2016年10月10日按小區用叉锨逐區采挖收獲,維持小區自然狀態越冬,對小區按坡度走向分別編號黃芪茬(HQ1、HQ2、HQ3),備用于次年當歸熟地育苗。
熟地黃芪茬口當歸育苗試驗于2017年6月1日在上述已培育的茬口進行,按小區撿拾石礫及草根后,每小區施有機肥2.5 kg,用鐵锨翻耕后耙平,每小區當歸種子播種量為25 g(折合播量52.0 kg·hm-2),采用撒播方法播種,播種后覆蓋麥草保墑增溫,覆蓋厚度以遮住土壤為宜。
1.3 岷縣生荒地當歸育苗
生荒地當歸育苗試驗于2017年6月1日開挖草皮后碎土撿拾草根,用鐵锨翻耕后耙平,育苗方法與熟地育苗方法一致,采用隨機區組設計,3次重復,即按坡度走向劃分為3個區組(3次重復),除生境不同外,小區面積、每小區施用有機肥量、當歸種子播種量、播種育苗方法均與熟地黃芪茬口完全一致。
上述生荒地和熟地黃芪茬口當歸育苗期均不施除草劑和農藥,其他農事田間管理均與大田一致。
1.4 當歸種苗抗逆指標測定
當歸育苗結束,2017年10月7日田間采挖種苗時對生荒地和熟地黃芪茬3個重復小區分別隨機取樣20苗,每處理共取樣60苗,立即拌鮮土放至采樣箱內低溫帶回實驗室,備用于病株統計和抗逆生理指標測定。其余種苗拌鮮土扎把越冬貯藏,備用于次年成藥栽培期返青成活率和早期抽薹率的測定。
當歸種苗細胞膜穩態性按照Guo等[19]的方法以測定相對電導率和氨基酸泄漏率衡量,滲透調節能力以測定可溶性糖和可溶性蛋白質含量衡量。當歸種苗脂質過氧化程度以測定丙二醛(malondialdehyde, MDA)含量和組織自動氧化速率(autoxidation rate, AR)衡量。AR測定5 h后的丙二醛含量,以每小時MDA含量的平均增加量來表示組織自動氧化速率,用于衡量活性氧自由基(reactive oxide species, ROS)對細胞的氧化攻擊程度[18-19]。采用氮藍四唑(nitro-blue tetrazolium, NBT)光化還原法測定超氧物歧化酶(superoxide dismutase, SOD)活性,采用愈創木酚顯色法測定過氧化物酶(peroxidase, POD)活性,采用紫外吸收法測定過氧化氫酶(catalase, CAT)活性,測定在240 nm處每分鐘光密度的變化量[18-19]。上述指標每重復小區測定3次,每處理共測定9次。
1.5 當歸種苗返青成活率和早期抽薹率的測定
種苗返青結束按重復小區依次統計返青成活株數和早期抽薹株數,最后分別計算返青率和早期抽薹率。
返青成活率(regreen-survival rate)=(移栽后返青成活株數/移栽總株數)×100%
早期抽薹率(early-bolting rate)=(早期抽薹株數/返青總株數)×100%
1.6 數據統計分析
采用Excel 2003進行處理平均數間的t檢驗并制圖。
2 結果與分析
2.1 岷縣生荒地和熟地豌豆勻田后黃芪育苗田植被和生境比較
岷縣禾馱鄉石家臺村大石溝大灣梁生荒地和熟地生境差異較大,生荒地地勢陡險(坡度30°),植被覆蓋度好,半山腰熟地農田以農作物為主,地勢較為平緩(坡度20°)。盡管生境有一定差異,但生荒地和熟地農田四周地埂均以艾草(Artemisiaargyi)為優勢種。圖1顯示,山頂生荒地當歸育苗區屬典型高寒草甸植被,以草本植物為優勢種群,種群結構復雜,物種多樣性豐富,與處于同一山脈半山腰的熟地農田豌豆輪作黃芪育苗地具有明顯差異,熟地豌豆茬口黃芪育苗田由于定期雜草管理,植物種群結構單一,黃芪優勢種生態位突出。由于生荒地大氣濕度大,當歸種子播種后較熟地出苗率高,群體長勢良好,但熟地種苗個體生長優勢較強,在2017年極端干旱條件下也獲得一定數量的當歸種苗(圖1)。
2.2 岷縣生荒地和熟地育成當歸種苗根系相對電導率和氨基酸泄露率的比較
生荒地和熟地豌豆-黃芪茬育成當歸種苗根系相對電導率和氨基酸泄漏率雖均具有一定差異,但差異性均未達到顯著水平(P>0.05),黃芪茬育成當歸種苗相對電導率更低(圖2)。然而,在生荒地育成的當歸種苗根系氨基酸泄露率更低。熟地黃芪茬較生荒地育成當歸種苗根系相對電導率下降15.29%(t=0.926,P>0.05),但氨基酸泄露率提高9.92%(t=1.518,P>0.05)。
生荒地和熟地黃芪茬育成當歸種苗根系可溶性糖含量和可溶性蛋白質含量雖有一定差異,但差異性也未達到顯著水平(P>0.05),黃芪茬育成當歸種苗可溶性糖含量與生荒地育成種苗的水平基本一致,但可溶性蛋白質含量低于生荒地的水平,說明黃芪茬熟地與生荒地育成當歸種苗糖類滲透調節能力一致,但蛋白質滲透調節能力稍有差異。生荒地育成種苗的蛋白質滲透調節能力更強。與生荒地育成種苗相比較,熟地黃芪茬育成當歸種苗可溶性糖含量僅提高0.44%(t=0.040,P>0.05),但可溶性蛋白質含量卻下降23.68%(t=1.240,P>0.05)(圖2)。
2.3 岷縣生荒地和熟地育成當歸種苗根系丙二醛含量與組織自動氧化速率的比較
熟地黃芪茬育成當歸種苗根系丙二醛含量和根系組織自動氧化速率均較生荒地育成種苗的水平提高,但兩者差異性不顯著(P>0.05),黃芪茬與生荒地育成當歸種苗MDA含量水平差異性很小,但生荒地種苗AR較低,說明熟地育成當歸種苗與生荒地育成當歸種苗根系膜脂質抗氧化能力略有差異,生荒地育成種苗的抗氧化能力更強,使根系MDA含量處于較低水平(圖3)。與生荒地育成種苗相比較,黃芪茬熟地育成當歸種苗MDA含量僅提高2.06%(t=0.478,P>0.05),而根系AR提高75.67%,但因熟地區組間變異程度大,變異系數48.14%,使兩者差異性未達到顯著水平(t=1.542,P>0.05)。
2.4 岷縣生荒地和熟地育成當歸種苗根系抗氧化酶活性的比較
不同生境條件下育苗對當歸苗根系抗氧化酶活性具有一定影響(圖4)。熟地黃芪茬口條件下培育的當歸種苗根系SOD和POD活性均較強,CAT活性較弱,但與生荒相育成種苗間的差異性均不顯著(P>0.05),熟地育成當歸種苗根系SOD活性較生荒地的水平提高7.15%(t=0.909,P>0.05),POD活性提高33.96%(t=0.601,P>0.05), CAT活性卻降低16.92%(t=0.903,P>0.05)。熟地育成種苗個體差異性較大,抗氧化酶類活性重復小區間的變異度均大于生荒地育成種苗的水平。
2.5 岷縣生荒地和熟地育成當歸種苗根病發病率和病情指數的比較
不同生境條件下育成當歸種苗越冬后種苗根部發病率均在25%~35%(圖5)。黃芪茬口熟地條件下培育的當歸種苗根部病害發病率相對較低,較生荒相育成種苗下降10.71%(t=0.802,P>0.05),病情指數下降25.00%(t=1.375,P>0.05),但各小區種苗發病株數差異性較大,導致小區間變異度大,降低了不同來源差異顯著性。
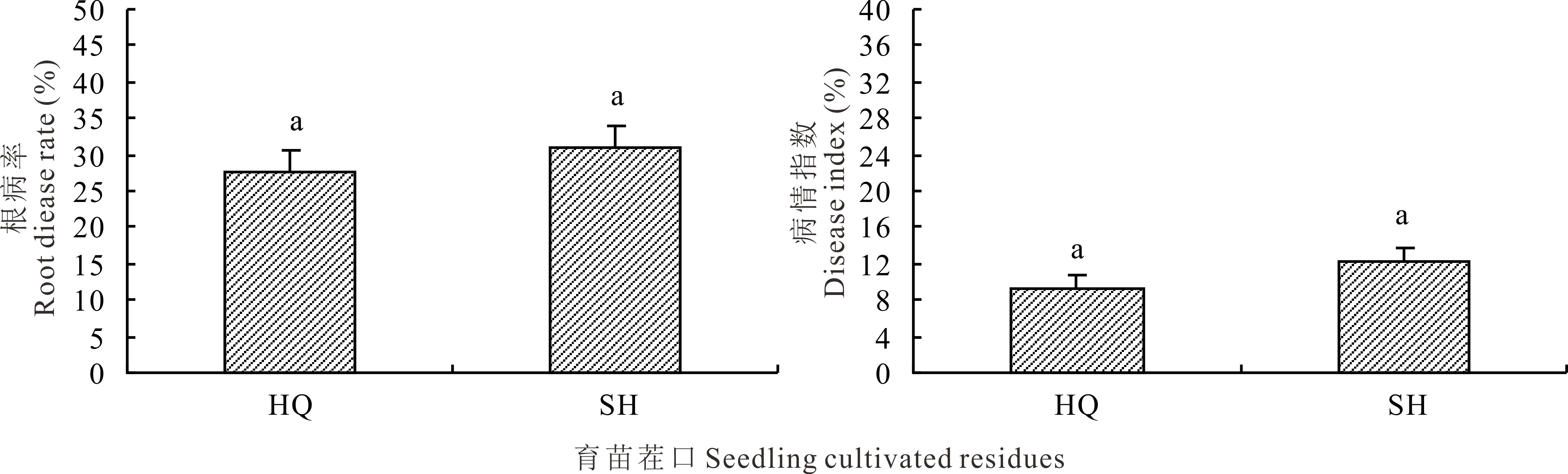
圖5 生荒地和熟地育成當歸種苗根病率和病情指數的比較Fig.5 Comparison of root disease rate and disease index in A. sinensis seedlings from the uncultivated and cultivated fields
2.6 岷縣生荒地和熟地育成當歸種苗移栽返青率和早期抽薹率動態的比較
不同土壤條件下育成當歸種苗春季移栽成藥栽培返青成活率和返青株早期抽薹率的變化動態差異均不顯著(圖6)。黃芪茬口熟地培育的當歸種苗移栽后返青勢強,至返青結束返青率始終高于生荒地種苗的水平,終返青率較生荒地育成種苗增加3.13%(t=0.402,P>0.05),但返青株早期抽薹率卻有所提高,平均早期抽薹率較生荒地種苗的水平提高5.32%(t=0.869,P>0.05),但各小區返青株中早期抽薹率差異性大,導致小區間變異度較大,兩者差異性未達到顯著水平。
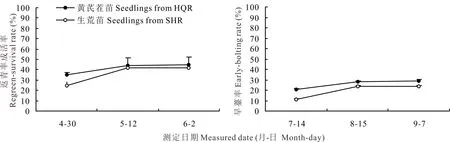
圖6 生荒地和熟地育成當歸種苗返青成活率和早期抽薹率的比較Fig.6 Comparison of regreen-survival rate and early-bolting rate in A. sinensis seedlings from the uncultivated and cultivated fields
種苗返青成活率與其根系CAT呈正相關,與POD活性的正相關達到顯著水平(P<0.05),與SOD活性的正相關達到極顯著水平(P<0.01),說明種苗抗氧化酶類活性的增強可提高當歸種苗返青成活率,SOD作用更大,POD次之,CAT相對較弱。早期抽薹率與返青率呈極顯著正相關(P<0.01),與抗氧化酶活性也均呈正相關,相關程度依次為POD>SOD>CAT,與POD和SOD間的正相關性達到顯著水平(P<0.05)。種苗丙二醛含量與組織自動氧化速率呈極顯著正相關(P<0.01),MDA含量愈高,AR越大,對種苗返青及生長發育越不利(表2)。
3 討論
3.1 熟地豌豆-黃芪茬育苗可維持當歸苗根系膜系統的穩態性
相對電導率和氨基酸泄露率是衡量植物膜系統完整性的重要指標,細胞膜穩態性差,膜系統易被氧化損傷,引起電解質泄露,相對電導率增大[18-19]。蛋白質的不穩定性可使氨基酸泄漏率增加[14]。植物在逆境下產生的大量活性氧自由基會引起膜脂質過氧化,進而損傷膜系統,MDA是膜脂過氧化的最終產物,濃度過高有毒害作用,但低濃度具有信號誘導生物體增強抗逆性的功能,MDA含量與膜穩態性和蛋白質穩定性相結合可衡量植物抗逆性,組織自動氧化速率是單位時間內MDA產生的速率,可直接反映細胞膜脂質過氧化反應的程度[18-19]。本研究中,當歸種苗組織自動氧化速率與MDA含量呈極顯著正相關進一步說明了兩者在抗逆性評價體系中的內在聯系,生荒地育成當歸苗蛋白質的穩定性更強,而熟地育成當歸苗膜系統穩態性更佳,因為生荒地育成苗根系氨基酸泄漏率和組織自動氧化速率略低,而熟地育成苗根系相對電導率和MDA含量略低,說明不同來源當歸種苗抗逆生理機制可能存在一定差異。2017年6-10月育苗期間降水不均勻,在6-7月正值當歸播種出苗期干旱少雨,但育苗后期降水多,可能導致熟地當歸苗逆境壓力勢更大,但種苗并沒有受到嚴重傷害,因為根系通過消耗可溶性蛋白質提高抗逆性,MDA處于較低濃度范圍,有利于激活其抗氧化系統機制,才得以維持種苗生物膜系統的穩態性。
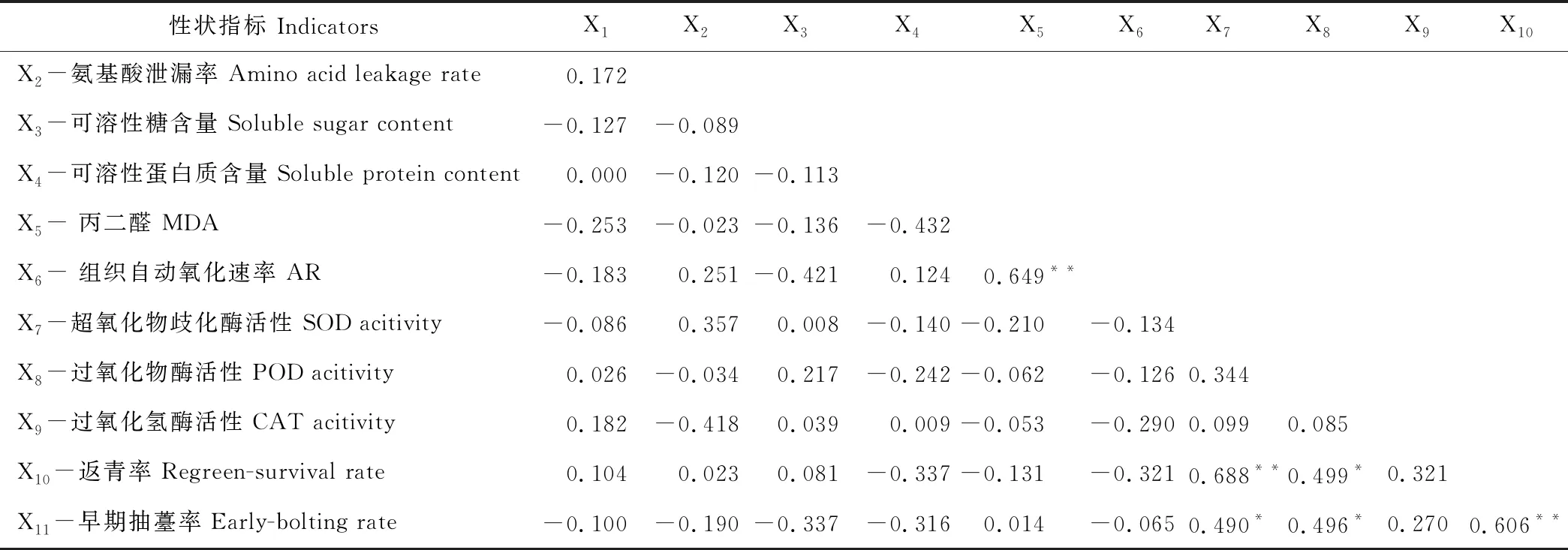
表2 生荒地和熟地育成當歸種苗抗逆生化指標與返青率和早期抽薹率的相關性分析Table 2 Relation of physicochemical stress resistant indicators to the regreen rate and early-bolting rate of A. sinensis seedlings from the uncultivated and cultivated fields
注: X1, 相對電導率。** 表示相關性達極顯著水平(P<0.01), * 表示相關性達顯著水平(P<0.05),n=18,R16,0.05=0.468,R16,0.01=0.590。
Note: X1, relative conductivity. ** means great significant correlation atP<0.01, * means significant correlation atP<0.05,n=18,R16,0.05=0.468,R16,0.01=0.590.
3.2 熟地豌豆-黃芪茬育苗可改善當歸苗根系糖類滲透調節能力
可溶性糖擔負著滲透調節作用,在植物旺盛光合作用期間通過滲透調節作用維護著細胞滲透勢、細胞水分平衡和光合作用[19]。一般植物的抗逆性與可溶性糖的大小呈正相關[20]。極端逆境影響植物的生長發育,可溶性蛋白作為保護性物質也參與植物細胞的滲透調節作用,可溶性蛋白含量與植物的抗逆性呈正相關[21-22]。本研究表明,豌豆-黃芪雙豆間迎茬條件下當歸種苗可溶性糖和可溶性蛋白含量均接近生荒地種苗的水平,但可溶性糖較生荒地種苗提高0.11%,可溶性蛋白質含量下降4.58%,盡管差異性未達到顯著水平,但可說明在遇干旱氣候條件下,生荒地育苗對當歸種苗有更適宜的環境優勢,環境壓力較小,也可說明熟地農茬口可造就環境壓力勢,雙豆茬輪作造就的環境壓力勢在一定程度上提高了當歸種苗的滲透調節功能及生理調控保護作用,從而有利于增強當歸種苗的抗逆性。
3.3 熟地豌豆-黃芪茬育苗可增強當歸種苗根系SOD和POD酶促抗氧化系統并降低根病率
生物自由基損傷理論證明當植物受到各種非生物因子脅迫時,細胞內活性氧(ROS)包括超氧陰離子(O2·-)、過氧化氫、羥基自由基(·OH)和單線態氧(1O2)等積累過多,其高度的氧化活性可破壞細胞膜結構,使植物抗逆性下降,導致植株發病甚至死亡,抗逆植物體內可進化出高效抗氧化機制,及時調控體內ROS以減輕這種氧化傷害[18]。SOD主要將最具有毒性的O2·-分解為H2O2和H2O,H2O2進一步由POD和CAT協同清除[19]。抗氧化系統發達的植株可抵抗氧化脅迫,有效保護膜系統[21-22]。本研究中,熟地豌豆-黃芪茬與生荒地育成當歸種苗根系內MDA含量基本一致,但抗氧化酶類活性存在一定差異。熟地育成種苗SOD和POD活性均高于生荒地種苗的水平,CAT活性卻低于生荒地的水平,盡管差異性未達到顯著水平,但也可以反映出熟地黃芪茬育苗環境對當歸具有一定環境脅迫勢,因為CAT是抗逆性最弱的抗氧化酶類,逆境條件下一般CAT最先受到影響,易失活。盡管黃芪茬育成當歸種苗CAT下降了16.92%,但也維持了一定的CAT活性,較生荒地種苗下降程度未達到顯著水平,這與金彥博等[23]在岷縣蠶豆(Viciafaba)-黃芪茬口基礎上研究的結果相似,進一步說明黃芪茬口培育的當歸種苗可以增強SOD和POD維持根系的生物抗氧化功能。這從種苗返青成活率得到進一步解釋,即豌豆-黃芪茬口育成當歸種苗較生荒地育成種苗早期抽薹率雖增加5.32%,但返青成活率提高3.13%,這可能是由于熟地育苗期間種苗受到干旱脅迫,加速了種苗的自我繁殖保護能力的根源所在,因為早期抽薹也是植物應對環境變遷的抗逆表現,營養生長期的干旱脅迫可誘發當歸物候的轉變,即從營養生長及早進入生殖生長,完成生活史。抗氧化系統發達的種苗返青成活率和早期抽薹率均高的結果也證明了這一點。由于土壤是植物生長生存的環境,植物可以改變土壤的生態環境,進而影響后茬植物的生長發育。植物對茬口的適應也是一種抗逆表現[24]。一般在田間最大持水量的60%范圍內,土壤含水量越高,土壤微生物的活性愈強,從而利于植物的生長發育[25]。雖然當歸重迎茬在土壤中積累了自毒物質,豆科作物豌豆和黃芪均屬于綠肥作物,可能對降解當歸自毒物質、促進種苗生長發育均具有重要作用。
4 結論
茬口形成的因素較為復雜,但茬口特性的好壞最終表現在后茬作物的產出性能上,盡管熟地豌豆-黃芪間茬口育成當歸種苗早期抽薹率較生荒地苗提高5.32%,但根病率下降10.71%,病情指數下降25.00%,由于根系SOD和POD活性較強,返青率提高7.52%,成藥率提高11.39%,有效補償了早期抽薹率偏高造成的產出損失,說明熟地適宜作物輪作茬口育成當歸種苗具有與生荒地種苗相似但有差異的抗逆生理機制,豌豆-黃芪倒茬輪作可替代生荒地進行當歸育苗。一般6月中旬麥草覆蓋育苗,生長期100 d即可采挖種苗,拌土入窖越冬,次年春季移栽大田進行成藥栽培,但熟地育苗期需加強水肥管理,底肥強化有機肥,根據天氣情況及時遮陽為宜。
致謝:岷縣禾馱鄉郎銀忠和李雪芳提供試驗地和合作,隴西縣鞏昌鎮郭志軍、郭天明和郭太平協助試驗地田間管理。中國科學院大學博士生陳翔,蘭州大學碩士生郭愛峰,甘肅農業大學碩士生梁偉、金彥博、許美玲、王雪琴參與育苗試驗,碩士生郭一青、王小琴、陳永中、劉蘭蘭,本科生甄世偉參與指標測定。在此一并致謝。
