毛白楊樹干呼吸及其溫度敏感性季節變化特征
2019-11-07 06:43:36郭麗麗楊慶朋喬雅君張茜茜梁偉佳鄭云普
生態學報 2019年18期
劉 亮,李 菲,郭麗麗,楊慶朋,喬雅君,聶 唯,楊 彬,張茜茜,曹 旭,梁偉佳,鄭云普,*
1 河北工程大學水利水電學院, 邯鄲 056038 2 中國科學院沈陽應用生態研究所/中國科學院森林生態與管理重點實驗室,沈陽 110164 3 河北雄安新區生態環境局,雄安 071700 4 中國農業大學水利與土木工程學院,北京 100083 5 中國水利水電科學研究院水利研究所,北京 100048
森林自養呼吸過程是陸地生態系統碳循環的重要組成部分[1],已經逐漸成為當前全球變化生態學研究領域的熱點問題之一[2,3]。以往的研究結果表明,森林生態系統總初級生產力的50%—70%通過自養呼吸的形式進入大氣[4- 6]。同時,樹干呼吸作為森林生態系統碳循環過程的重要組成部分,約占整個森林生態系統自養呼吸的12%—42%[7]。盡管如此,樹干呼吸相關研究卻在整個森林生態系統碳循環研究中被關注的相對較少,以至于目前對于森林生態系統碳儲量的估算還存在很大的不確定性[4,6]。
樹干呼吸(Es)是一個非常復雜的生物學過程,CO2氣體主要是由樹干木質部、韌皮部和形成層等組織的活細胞通過生理的有氧呼吸代謝過程而產生[5]。樹干呼吸過程不僅受到溫度、水分和光輻射等多種環境因素的影響[2,7],而且還會根據樹干液流、生長速率和林分狀況的不同而發生明顯變化[1,3,6]。因此,難以準確判斷究竟哪些關鍵因素直接決定著樹干的呼吸過程。通常而言,溫度作為重要的環境因素可能直接影響Es[8- 10]。目前,大多數學者認為溫度可能通過多個不同的途徑對樹干呼吸產生影響,主要包括溫度升高增加細胞的呼吸速率,增加CO2的擴散系數或者降低液流中溶解CO2的濃度[10-14]。以往許多研究已經發現Es和樹干溫度之間具有較好的相關性[11],故研究者往往利用以溫度為變量的函數估算Es,且認為Es隨著樹干溫度呈現指數增加的趨勢,其與樹干溫度的關系一般表示為溫度敏感指數(Q10)[12- 15]。然而,另外的一些相關研究卻發現Es和樹干溫度的相關性并不顯著,甚至根本不存在相關性[9,16],即使將Es進行溫度標準化校正后,Es在樹種和林分間卻仍存在較大的時空變率[17]。因此,樹干Es及其對溫度的響應(Q10)呈現出較大的時空變異性,以至于對森林生態系統碳儲量的估算造成偏差[3,6],這在一定程度上限制了森林碳平衡模型的發展[4]。
以往大多數的研究結果表明,樹干呼吸及其溫度的敏感性隨季節的不同而呈現出明顯的動態變化特征[10,13,16]。在不同季節里林木的光照條件、土壤水分狀況、氮素利用率等都可能存在較大的差異[15,17],從而導致樹干內部的非結構性碳水化合物含量的改變[12],最終造成樹干Es和Q10的顯著變異[16,17]。楊慶朋[18]對我國亞熱帶典型人工林連續2年的監測數據發現,3個樹種生長季的Q10值均明顯低于非生長季,且將生長季和非生長季的數據整合一起得到的Q10與單獨生長季或非生長季的數據推求出的Q10存在很大差異。Es和Q10的季節變異可能與樹干體內非結構性碳水化合物的含量及形態的季節性變化存在一定關系,但還需要進一步驗證[18]。另外,為了深入理解樹干呼吸不同組分對溫度變化的響應機理,學者們從生理上將樹干呼吸劃分為提供能量合成新細胞的生長呼吸和維持細胞生命活動的維持呼吸兩個部分[19],并假設秋冬季樹木停止生長后的樹干呼吸即為維持呼吸,并認為維持呼吸對溫度的響應在年際尺度上是恒定的[19]。以往大多數研究發現維持呼吸對溫度的響應較為敏感,而生長呼吸則對溫度并不敏感[20],故不同季節生長呼吸和維持呼吸所占的比例并不相同,也可能造成Q10和Es的季節性變異。此外,盡管Q10和Es作為生態系統過程模型的重要參數被廣泛應用于預測未來氣候變暖對陸地生態系統影響的模擬研究[21],但該參數的估算卻僅依賴于樹干組織內的氮素含量[3,8,15],而并未考慮Q10和Es的季節變異,從而導致對于陸地生態系統碳儲量的估算存在很大的不確定性[4,6,18]。
毛白楊(Populustomentosa)作為華北平原區農田防護林的主栽樹種之一,在該區域具有較大的分布面積和栽培規模。同時,農田防護林也是陸地生態系統的重要組成部分,但長期以來人們主要關注的是其經濟價值和防護效益,而往往忽視其在陸地生態系統碳循環方面所起到的關鍵性作用。本研究以毛白楊為研究對象,連續監測毛白楊樹干呼吸速率和樹干溫度變化,深入分析樹干呼吸速率的晝夜變異以及季節性動態變化規律,旨在揭示影響毛白楊樹干呼吸速率及其溫度敏感性的潛在機理,以期為準確估算陸地生態系統碳儲量和預測全球變暖進程提供基本的數據支撐和理論依據。
1 材料與方法
1.1 研究區概況
本研究樣地位于河北省邯鄲市南郊區的典型毛白楊農田防護林內(東經114°30′,北緯36°25′)。研究區域屬典型暖溫帶半濕潤大陸性季風氣候,日照充足,雨熱同期,四季的交替明顯。該地區多年平均降雨量548.9 mm,主要集中在7—8月份,年平均氣溫14℃,最冷月份(1月)平均氣溫為-2.5℃,最熱月份(7月)平均氣溫27℃,全年無霜期200 d,年日照2500 h。研究樣地內的土壤質地以粉砂和輕壤為主;pH值較高,為7.5—8.0;土壤有機質含量較低,為6.0—10.0 g/kg; 全N含量為0.5—0.7 g/kg。
1.2 樹干呼吸的測定
于2015年11月在試驗區分別選取3個區域,在3個區域內分別選取生長良好的6棵毛白楊林木(共計18棵)用于樹干呼吸速率和樹干溫度的連續監測(2016年1月份—2017年1月份,共計監測23次,具體日期見圖3)。其中2016年4月26日(116 d)對樹干呼吸進行了24 h的監測,其余的樹干呼吸監測時間為8:00—20:00。
樹干呼吸測定采用Li-Cor 6400便攜式光合作用測量儀配套使用的呼吸測量氣室。先將PVC環的一端切割成弧形以匹配樹干的弧度,另一端磨平用以連接呼吸測量室。在選擇的毛白楊樹干1.5 m高度處安裝PVC環(內徑10 cm,深度5 cm)。安裝前輕微刮掉表層樹皮,盡量確保樹干表面的平整性,但不能損傷到形成層組織。用硅膠將呼吸環固定在樹干表面上,檢查其密封性,確保不漏氣。在呼吸環的右側5 cm位置鉆取一個細孔,使Li-Cor 6400自帶的溫度探頭剛好插進樹干,在監測樹干呼吸的同時測量樹干溫度。
測定樹干呼吸時,將Li-Cor 6400呼吸測量室扣在樹干上固定的呼吸環,每間隔2—3 h對18棵毛白楊樹干呼吸及樹干溫度進行1次測定。對18棵樹干呼吸及樹干溫度取平均值,作為每次測定的均值;對每天內的數據取平均值作為當天測定的均值。樣地內林木及其測定位置的基本特征見表1。根據毛白楊在本研究區的物候特征(展葉期為4月份,落葉期為11月份)將整個研究期劃分為生長季(2016年5月份—10月份)和非生長季(2016年11月份—2017年1月份)。進入非生長季后樹木開始落葉,研究區進入秋季,樹木停止生長。本研究認為在非生長季測定的呼吸即是維持呼吸,而在生長季測定的呼吸則是生長呼吸和維持呼吸的總和[19]。

表1 毛白楊樹干呼吸監測林木及樣地特征
1.3 樹干可溶性糖和淀粉的含量分析
每塊樣地附近隨機選取2棵與測定樹相似的樣樹,共計6棵白楊樹,分別在生長季(2016年5月18日,第138天)和非生長季(2016年11月23日,第327天)使用生長錐鉆取樹芯樣品,取樣孔徑12 mm,取樣深度4 cm。考慮到樹干內碳水化合物的日變化,故取樣時間統一在9:00—11:00進行[1]。將樹芯樣品于烘箱內在75 ℃條件下烘干72 h至恒重。樣品烘干后粉碎封閉儲存。采用高壓液相色譜法(HPLC)測定樹芯樣品中的淀粉和可溶性糖(蔗糖、葡萄糖、果糖)含量。
1.4 數據分析
為了計算樹干呼吸速率,需要獲得PVC環所圍樹干面積以及氣室插入PVC環的有效深度。根據公式(1)計算PVC環所圍的樹干面積[18]。
(1)
A指PVC環所圍的樹干面積,Dc指PVC環的直徑,Ds指樹干直徑。
根據公式(2)計算氣室插入PVC環的有效深度[18]。
H=(Vc-(Dc/2)2πd)/(Dc/2)2π
(2)
H指有效插入深度,Vc和Dc指樹干上PVC環的體積和直徑。
本研究采用公式(3)擬合樹干呼吸速率和樹干溫度之間的關系[22],按公式(4)計算溫度敏感系數Q10[23]。
ES=β0eβ1T
(3)
式中:β0為當樹干溫度為零時的樹干CO2釋放通量;β1為溫度系數。
Q10=e10β1
(4)
式中:Q10為樹干溫度每增加10℃呼吸速率增加的倍數。
為了消除溫度的干擾,比較相同樹干溫度時樹干呼吸速率在不同時間和季節上的差異,需將毛白楊樹干呼吸統一矯正為15℃時的呼吸速率(R15)[21]。R15由樹干呼吸和樹干溫度的回歸方程(E-T曲線)得到。
本研究數據利用單因素方差分析的方法(ANOVA)比較林木組間的顯著性差異(n=18)。所有的統計分析均利用SPSS 13.0 (Chicago, IL) 統計軟件完成,所有作圖均利用Excel軟件實現。
2 結果與分析
2.1 毛白楊樹干呼吸的日變化特征

圖1 生長季樹干溫度和樹干呼吸速率的日變化 (±標準誤,n=18)Fig.1 Diurnal variation of stem respiration and stem temperature during growing season (±SE,n=18)
2016年4月26日(116天),對毛白楊樹干呼吸及樹干溫度進行24 h監測,結果顯示毛白楊樹干溫度呈現出先升高后降低的變化趨勢,且樹干溫度的最高值為22.8℃,出現在下午13:00,最低值為15.3℃,出現在次日凌晨5:00時(圖1)。然而,毛白楊樹干呼吸速率的變化趨勢同樹干溫度的變化并不一致,具體表現為樹干呼吸速率的最大值出現在傍晚約19:00,樹干呼吸速率最大值的出現相比樹干溫度最高值后滯了8 h,且樹干呼吸速率在白天隨時間推移呈現出微弱降低的趨勢,從早晨7:00的1.0 μmol CO2m-2s-1逐漸降低為下午15:00的0.76 μmol CO2m-2s-1,盡管在下午17:00樹干呼吸速率略微升高。本研究還發現毛白楊在夜間的樹干呼吸速率明顯高于白天,具體表現為毛白楊樹干呼吸速率從下午17:00到傍晚19:00出現明顯的跳躍,樹干呼吸速率由大約0.8 μmol CO2m-2s-1突然升高到最高值1.42 μmol CO2m-2s-1,相比樹干呼吸速率最低值增加87%(圖1)。
對毛白楊樹干溫度和樹干呼吸速率進行指數函數擬合的結果顯示,毛白楊樹干溫度與樹干呼吸速率存在較好的指數關系(E-T曲線),但在白天(7:00—19:00)和夜間(19:00—7:00)存在明顯的差異(圖2)。白天的毛白楊樹干溫度與樹干呼吸速率呈現負相關的關系,樹干溫度解釋了樹干呼吸速率變化的88%。然而,在夜間樹干呼吸速率與溫度之間表現為正相關關系,樹干溫度解釋了樹干呼吸速率96%的變異(圖2)。
2.2 毛白楊樹干呼吸及溫度敏感性的季節動態特征
毛白楊樹干呼吸速率在季節尺度上同樹干溫度的變化趨勢具有較好的一致性(圖3)。在整個觀測期內,毛白楊樹干溫度和樹干呼吸速率均隨季節變化呈現出先升高后降低的變化趨勢,樹干呼吸的變化范圍為0.11—2.63 μmol CO2m-2s-1,樹干溫度的波動范圍為1.8—33℃。毛白楊樹干呼吸在春季(4月初)開始升高,并在夏季(8月末)達到最高值2.63 μmol CO2m-2s-1,而后隨氣溫的降低逐漸下降,在冬季(12月中旬)達到最低值0.11 μmol CO2m-2s-1(圖3)。在整個觀測期內毛白楊樹干呼吸速率同樹干溫度之間也存在著較好的指數函數關系(E-T曲線),樹干溫度可以解釋毛白楊樹干呼吸速率季節變異的93%,毛白楊樹干呼吸的溫度敏感性指數(Q10)為2.62(圖4)。
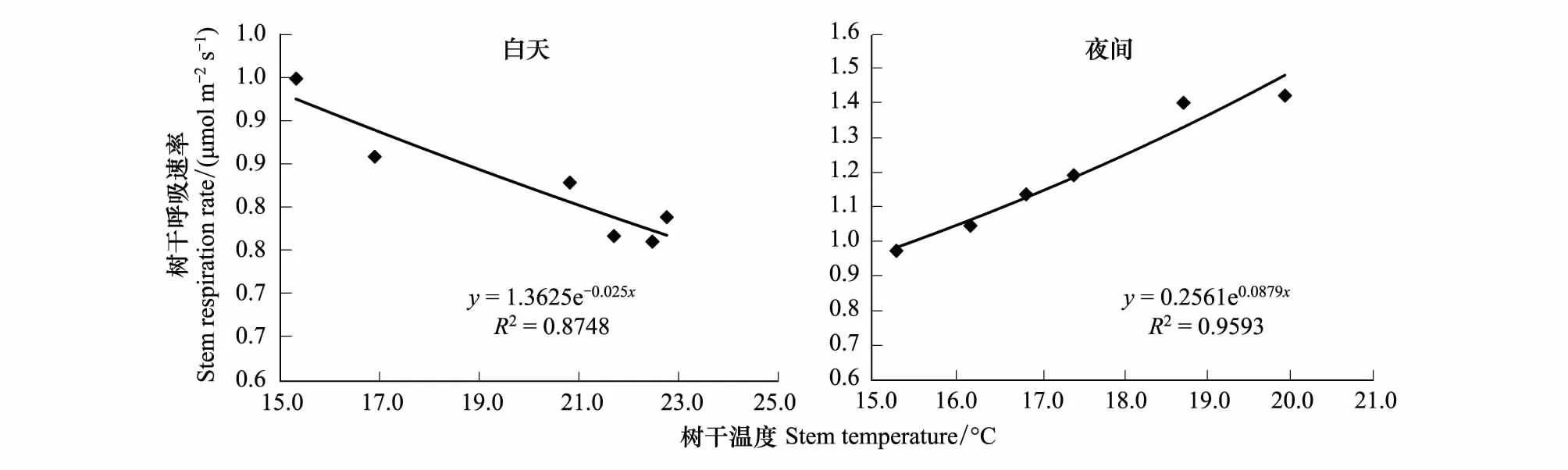
圖2 日變化尺度上樹干呼吸和樹干溫度的關系Fig.2 Relationships between stem respiration and stem temperature during diurnal variation

圖3 樹干呼吸和樹干溫度的季節變化(±標準誤,n=18)Fig.3 Seasonal variation of stem respiration and stem temperature (±SE,n=18)

圖4 整個研究期樹干呼吸和樹干溫度的關系 Fig.4 Relationships between stem respiration and stem temperature during the observation periods
根據毛白楊物候特征,將整個研究期劃分為生長季和非生長季,結果顯示生長季和非生長季的樹干呼吸和樹干溫度之間都存在指數函數關系(生長季R2=0.68,非生長季R2=0.79),表明樹干溫度在生長季和非生長季可以分別解釋樹干呼吸季節變化的68%和79%(圖 5)。另外,本研究結果還顯示,非生長季樹干呼吸的溫度敏感性系數(Q10=3.00)明顯高于生長季(Q10=1.95)(圖5)。
本研究結果顯示,毛白楊樹干呼吸在夜間的R15為0.58 μmol CO2m-2s-1,顯著高于白天的R15值0.35 μmol CO2m-2s-1(P<0.01,圖 6)。在季節尺度上R15的變化卻與溫度變化呈現比較一致的趨勢。生長季毛白楊樹干呼吸(生長呼吸與維持呼吸之和)的R15為0.65 μmol CO2m-2s-1,顯著高于非生長季毛白楊樹干呼吸(維持呼吸)的R15值0.52 μmolCO2m-2s-1(P<0.05,圖 6)。
2.3 樹干呼吸速率與樹干非結構性碳水化合物的關系
本研究結果顯示,毛白楊樹干可溶性糖和淀粉的含量在不同季節間的差異顯著,且生長季毛白楊樹干可溶性糖及淀粉的含量均明顯高于非生長季(P<0.05,圖 7)。具體而言,生長季毛白楊樹干的可溶性糖含量為86.69 mg/g,而非生長季的可溶性糖含量降低為63.83 mg/g,即毛白楊生長季的樹干可溶性糖含量比非生長季增加36%。相似地,生長季毛白楊樹干的淀粉含量為26.24 mg/g,明顯高于非生長季的淀粉含量(18.24 mg/g)44%。
本研究分別擬合了生長季和非生長季毛白楊樹干呼吸(R15)與樹干可溶性糖或淀粉含量的線性關系(圖 8)。擬合結果顯示,就生長季而言,毛白楊樹干的可溶性糖含量與R15的相關性(R2=0.52)高于淀粉(R2=0.35);相反,毛白楊樹干的淀粉含量與R15在非生長季的線性關系(R2=0.59)相比可溶性糖含量與R15的相關性更好(R2=0.50)。

圖5 生長季和非生長季樹干呼吸與樹干溫度的關系Fig.5 Relationships between stem respiration and stem temperature during growing season and non-growing season
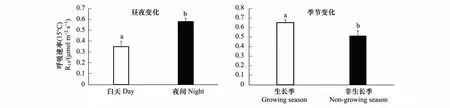
圖6 溫度矯正樹干呼吸的晝夜變化和季節變異(±標準誤,n=18)Fig.6 Diurnal and seasonal variations of temperature-normalised stem respiration (±SE, n=18)

圖7 樹干可溶性糖和淀粉含量的季節變化(±標準誤,n=6)Fig.7 Seasonal variations of the stem soluble sugar and starch contents (±SE,n=6)

圖8 樹干可溶性糖和淀粉濃度與R15在不同季節的關系Fig.8 Relationships of soluble sugars and starch with R15 in the growing season and non-growing season
3 討論
3.1 樹干呼吸日變化分析
樹干呼吸日變化往往會受到樹干溫度和樹干液流的影響,且樹干溫度和樹干液流存在明顯的晝夜變化[24],導致影響樹干呼吸日變化的機理極其復雜,以至于目前對于森林碳儲量的估算還存在很大的不確定性[8,10]。以往的大多數研究結果表明,森林樹干呼吸日變化曲線為雙峰型[19]和單峰形[25],且樹干呼吸速率最大值出現在16:00—20:00時,而最小值則出現于4:00—10:00[25]。同時,另有相關研究結果發現樹干呼吸通常滯后于樹干溫度約2—3個小時[26]。然而,本研究結果顯示毛白楊樹干呼吸晝夜動態變化呈單峰型曲線,其峰值出現在傍晚大約19:00時,且樹干溫度與樹干呼吸在白天呈現出相反的變化趨勢。以往的相關研究也發現類似的現象,Zhu等[27]在對木荷樹干呼吸的研究中發現在7月份,樹干呼吸和樹干溫度呈現出負相關的現象,并解釋為由于高溫抑制了樹干呼吸作用,導致了樹干呼吸速率的降低。王秀偉等[28]的研究也發現,3個樹種(落葉松、水曲柳、白樺)的樹干呼吸與樹干溫度在6月份呈現出相反的趨勢,其原因可能是白天樹干液流速度大,樹干呼吸產生的部分CO2可能通過植物的蒸騰作用而進入大氣中,故低估了樹干呼吸的測量值,從而導致樹干表面CO2釋放量在夜間高于白天。本研究監測顯示在4月份樹干溫度最大值為22.76℃,故不可能是高溫抑制樹干呼吸,而應該是白天樹干液流帶走部分CO2氣體而造成的,即造成樹干呼吸峰值出現在傍晚時的主要原因是由于白天樹體內部生成的部分CO2氣體被樹干液流運輸至葉片并通過蒸騰作用釋放到大氣中[29]。事實上,以往的大多數研究也發現樹干呼吸與樹干液流之間存在顯著的負相關關系[27,30],樹體內部細胞代謝過程產生的CO2氣體并不會全部通過樹皮擴散到外界大氣中,而是一部分CO2氣體溶解于樹干液流中并向上運輸,最終通過葉片氣孔釋放到大氣環境,以至于許多研究發現樹干呼吸峰值出現在溫度較低的夜間[29,31]。另外,本研究分別擬合白天和夜間毛白楊樹干呼吸對溫度響應的曲線,結果顯示樹干呼吸和樹干溫度在夜間的相關系數高于白天,這可能是由于夜間受到液流的干擾小,樹干呼吸對溫度的響應更加敏感,而白天液流對樹干呼吸的干擾較大,從而降低了樹干呼吸對溫度響應的敏感性。然而,液流對樹干呼吸速率的影響程度在不同季節、不同氣候條件甚至不同樹種間存在很大的變異[27,29]。因此,目前關于液流對樹干呼吸產生影響的潛在機理還存在較大爭議[27,28],有待于在不同森林類型中更進一步的深入探討和研究。
3.2 樹干呼吸及溫度敏感性的季節變化規律
在整個觀測期內樹干呼吸呈現出明顯的季節變化規律(圖3),且同樹干溫度具有很好的相關性(R2=0.93,圖4),表明在季節變化的尺度上樹干溫度可能是控制樹干呼吸的主要因素之一,因為在較高樹干溫度時樹體內部細胞的線粒體呼吸速率增大,從而增加CO2氣體的釋放量,而且還會提高CO2氣體從樹體內部到外界大氣的擴散系數[21]。另外,本研究分別擬合生長季和非生長季E-T曲線的結果表明,相對于生長季而言,非生長季毛白楊樹干呼吸與溫度之間具有更好的指數回歸關系(非生長季時的R2高于生長季),該結果與Ryan的研究結論一致[32]。本研究結果表明,在非生長季樹干呼吸全部為維持呼吸,維持呼吸對溫度的響應更加敏感,故其相關性更高,回歸系數更大。然而,在生長季所測樹干呼吸為生長呼吸與維持呼吸之和,生長呼吸對溫度的響應不敏感。因此,生長季樹干呼吸與溫度的回歸系數較小。同樣,王淼等的研究也發現幼樹的樹干呼吸與溫度的相關性比老樹較差,主要是由于幼樹處于快速生長階段,故其生長呼吸部分占總呼吸的比例較大,從而導致樹干呼吸與樹干溫度相關性的下降[25]。此外,以往大多數研究結果表明樹干呼吸溫度敏感性(Q10)隨著物種、氣候條件、底物供應狀況等的變化而發生改變[30,33],樹干呼吸Q10值的變化范圍為1.0—4.4[34-35]。本研究結果顯示,毛白楊樹干呼吸非生長季的Q10值(3.00)高于生長季的Q10值(1.95),非生長季樹干呼吸主要為維持呼吸,這也表明維持呼吸對溫度的響應更加敏感。然而,利用整個觀測期數據擬合得到的Q10為2.62,與在生長季和非生長季擬合出的Q10也存在很大的不同,表明在不同的時間尺度上,利用E-T曲線擬合出的溫度敏感指數存在很大的變異。因此,基于生態系統過程模型預測森林生態系統碳循環過程對氣候變暖響應的相關研究應該充分考慮到森林樹干呼吸過程在不同季節(生長季和非生長季)響應溫度變化存在差異的情況。
3.3 可溶性糖與淀粉的季節變化對樹干呼吸的影響
毛白楊生長季的R15顯著高于非生長季,表明除溫度以外的其他因素也可能在毛白楊樹干呼吸季節變化中起作用。本研究對不同季節樹體內非結構性碳水化合物含量(可溶性糖和淀粉)測定的結果顯示,生長季毛白楊樹干中的可溶性糖和淀粉含量均顯著高于非生長季,表明毛白楊生長季R15高于非生長季的原因可能是由于生長季可利用的呼吸底物(非結構性碳水化合物)含量比非生長季更高而造成的。以往的一些研究結果也表明非結構性碳水化合物含量與樹干呼吸之間存在著密切的相關性[21,36]。例如,Maier等發現火炬松樹體組織中的非結構性碳水化合物含量與樹干呼吸存在明顯的正相關關系[21]。在生長季樹干中可溶性糖與R15存在更好的相關關系;而在非生長季樹干中的淀粉與R15存在更好的相關關系。這表明在非生長季樹干呼吸的主要底物形式為淀粉,樹體內儲存的大量淀粉物質被消耗用于林木維持呼吸過程,以至于樹干組織的非結構性碳水化合物含量降低[37];然而,生長季樹干呼吸的底物主要以可溶性糖為主,且林木通過光合作用過程生成大量非結構性碳水化合物[37],并迅速以可溶性糖的形式運送至樹體用于樹干呼吸過程,進一步為林木新陳代謝過程提供能量。
4 結論
本研究發現毛白楊樹干呼吸的Q10在不同季節存在明顯變異,且生長季Q10值(1.95)顯著低于非生長季Q10值(3.00),表明毛白楊樹干呼吸不同組分對溫度響應的敏感程度并不相同。另外,生長季樹體內的可溶性糖含量與樹干呼吸具有較好的相關性,但非生長季淀粉含量同樹干呼吸的相關性更加明顯,表明呼吸底物類型及其供應狀況(非結構性碳水化合物)在一定程度上決定著毛白楊的樹干呼吸速率及其溫度敏感性,即毛白楊樹干呼吸的溫度敏感性(Q10)其實并非為呼吸過程對溫度的真實響應,而是受其他多種因素(如呼吸底物類型、底物供應狀況等)和生理過程共同影響的表觀溫度敏感性(ApparentQ10)。因此,利用生態系統過程模型估算森林生態系統呼吸非常有必要將樹干總呼吸區分為生長呼吸和維持呼吸,并分別計算兩者的溫度敏感性,避免利用樹干總呼吸的表觀Q10直接估算森林生態系統呼吸而帶來的誤差,以至于高估氣候變暖對樹干呼吸過程的影響,進而低估森林生態系統呼吸對未來氣候變暖的反饋能力。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
