外源ABA對干旱脅迫下多年生黑麥草光合特性的影響
2019-11-07 10:41:42許喆,任健,田英,米楠*
草地學報 2019年5期
許 喆,任 健,田 英,米 楠*
(1.寧夏大學農學院,寧夏 銀川 750021; 2. 云南農業大學草業科學系,云南 昆明 650201)
植物在生長中受到各種環境因素的影響,干旱是植物生長發育最重要的限制因子之一,對植物生長和生物量的形成均有一定影響[1]。當植物處于干旱脅迫時,體內的脫落酸(Abscisic acid,ABA)會大量增加,從而提高對逆境的抗性[2],降低干旱對植物細胞膜的傷害[3]。ABA是公認的五大植物激素之一,在調控植物生長發育、應對非生物脅迫中發揮著重要作用,能調節植物氣孔開閉、休眠、葉片衰老等[4-5]。研究發現,噴施外源ABA能延緩番茄(LycopersiconesculentumMill.)[6]、玉米(ZeamaysL.)[7]、香蕉(MusananaLour.)[8]、水稻(OryzasativaL.)[9]等植物葉片中葉綠素的降解速度,增加脯氨酸含量和保護酶的活性。外源ABA可以提高植物的抗冷性和保水能力,促使氣孔關閉、降低蒸騰速率、提高光合作用及葉片含水量,減少干旱脅迫對植物細胞與組織的傷害[10-12]。
多年生黑麥草(Loliumperenne)屬禾本科黑麥草屬植物,是全世界種植最廣泛的冷季型草種之一[13-14],在我國主要分布在華東、華中和西南等地。由于黑麥草再生性強、耐刈割、耐放牧[15],具有成坪快、品質優良、耐踐踏等優點,是草坪建植重要的先鋒草種[16],也是草食家畜的優質牧草和養魚的好飼料[17]。然而,多年生黑麥草的根系較淺,主要分布在15厘米深的表層土壤中[18],所以很容易受到干旱脅迫的影響。有研究表明,在干旱環境下多年生黑麥草的過氧化物酶(Peroxidase,POD)、超氧化物歧化酶(Superoxide Dismutase,SOD)等酶活性升高,并帶動其他酶活性的變化[19-20]。目前有關干旱對多年生黑麥草的影響方面的研究報道較少,生長調節劑對多年生黑麥草抗旱性的影響尚不清楚。因此,本研究以多年生黑麥草為材料,探究干旱脅迫下外源ABA對多年生黑麥草耐旱性的影響,尤其是光合能力的變化,為干旱氣候下多年生黑麥草草坪管理提供理論基礎與參考。
1 材料與方法
1.1 供試材料和試驗設計
供試材料為多年黑麥草(品種:冬牧)。選取大小、形狀、色澤一致且飽滿的種子,先用0.1%HgCl溶液消毒15 min,清水沖洗干凈,播種至育苗盆(19 cm×15 cm×6 cm)中培養,培養基質為蛭石。2017年4月12日于人工氣候箱中發芽,人工氣候箱培養條件:白天溫度(25±1)℃,夜晚溫度(20±1)℃,每天光照12 h,黑暗12 h,光強4 000 lx,濕度70%。發芽后用Hoagland營養液培養15 d后,平均株高10 cm時開展干旱脅迫。利用聚乙二醇(Polyethylene glycol,PEG 6000)模擬干旱脅迫,該化合物是一種高分子滲壓劑,性質穩定,方便易行[21]。脫落酸ABA為國藥集團化學試劑有限公司生產。
采用雙因素隨機區組試驗,試驗因素包括ABA和PEG,其中ABA設3個梯度:0(A0),200 μmol·L-1(A2)和500 μmol·L-1(A5),葉面噴施,對照組以等量蒸餾水代替。PEG設3個梯度:正常澆水(PEG 0%)、中度干旱(PEG濃度10%)和重度干旱(PEG濃度30%),對照以等量的Hoagland營養液代替。土壤水分傳感器測得土壤水分的體積百分比分別為65%,51%和34%。試驗共9個處理,即ABA 0 μmol·L-1+PEG 0%(CK),ABA 200 μmol·L-1+PEG 0%(A2P0),ABA 500 μmol·L-1+PEG 0%(A5P0),ABA 0 μmol·L-1+PEG 10%(A0P10),ABA 0μmol·L-1+PEG 30%(A0P30),ABA 200 μmol·L-1+PEG 10%(A2P10),ABA 200 μmol·L-1+PEG 30%(A2P30),ABA 500 μmol·L-1+PEG 10%(A5P10),ABA 500 μmol·L-1+PEG 30%(A5P30),每處理重復三次。ABA2 d噴施一次,每次各噴施15 ml,PEG培養液3 d澆灌一次,每次各200 ml,脅迫時間為1個月后測定相關指標。
1.2 葉綠素含量
采用N-N-甲基甲酰胺浸提法測定[22]。
1.3 光合參數
2017年6月,選擇葉片全展開、無病蟲害的葉片進行光合作用測定。利用LI-6400XT光合-熒光測定儀(美國LI-COR公司)上午9:00-11:00在室內測定葉片凈光合速率(Net photosynthetic rate,Pn)、胞間CO2濃度(Intercellular carbon dioxide concentration,Ci)、蒸騰速率(Transpiration rate,Tr)、氣孔導度(Stomatal conductance,Gs)等光合生理參數,計算出水分利用效率(Water use efficiency,WUE=Pn/Tr),使用開放式氣路,CO2濃度400 μmol·mol-1,飽和光強設定為800 μmol· (m2·s)-1,測定時光照強度穩定3min[23]。
1.4 葉綠素熒光
采用LI-6400-40葉綠素熒光系統葉室,選擇無傷害有代表性的葉片完全暗適應12 h。測定光合的初始熒光(Basic fluorescence,Fo)、最大熒光(Maximum fluorescence,Fm)和可變熒光(Variable fluorescence,Fv),計算PSII最大光化學量子產量(PSII Maximum photochemical quantum yield,Fv/Fm)、PSII潛在活性(PSII potential activity,Fv/F0)。葉片光適應20min后進行測定,非光化學猝滅(Photochemical quenching coefficient,qP)、光化學猝滅(Non-photochemical quenching,NPQ)等熒光參數。每處理測定3個葉片,各熒光參數重復記錄6次。
1.5 數據處理與統計方法
利用SPSS19.0統計分析軟件對數據進行方差分析(ANOVA),不同處理間差異比較(Mean±SE)采用Duncan法,數據均用EXCEL軟件作圖。
2 結果與分析
2.1 對葉綠素含量的影響
干旱脅迫顯著影響了多年生黑麥草的葉綠素含量(P<0.05)(圖1)。與CK相比,中度干旱脅迫使葉綠素a,b和總葉綠素含量分別提高了13.7%,15.5%和13.8%,而重度干旱脅迫下外源ABA 500 μmol·L-1與未噴施ABA處理相比葉綠素a,b和總葉綠素含量分別上升77.9%,75.9%和77.2%。說明ABA能夠抑制正常生長和中度干旱脅迫下的葉綠素含量,但是,適度干旱脅迫能增加多年生黑麥草的葉綠素含量,在重度干旱脅迫下噴施外源ABA,尤其是ABA 500 μmol·L-1使葉綠素a,b和總葉綠素含量變化顯著(P<0.05)。
2.2 對光合參數的影響
干旱脅迫對多年生黑麥草的光合參數影響顯著(圖2),導致葉片的光合機構數量和活性下降,導致光合作用減弱。在重度干旱脅迫下與CK相比,多年生黑麥草Pn,Gs和WUE分別下降124.8%,155.8%和57.9%,原因是氣孔導降低(P<0.05),從而抑制了植物的光合作用。
ABA僅對多年生黑麥草Gs產生了明顯的影響(P<0.05)。與對照相比,噴施外源ABA 200 μmol·L-1對凈光合速率(Pn)的影響不明顯,但顯著提高了水分利用效率(WUE)(P<0.05),相反胞間CO2濃度(Ci)下降了23.7%(P<0.05)。外源ABA 500 μmol·L-1使凈光合速率(Pn)下降了47.6%,ABA能夠抑制正常生長條件下的凈光合速率(Pn)。
由于多年生黑麥草在中度干旱脅迫下具有抗旱性,因此,噴施不同濃度的ABA產生的影響不顯著。氣孔是植物對干旱脅迫反應的重要標志,是植物氣體交換的“窗口”,在重度干旱脅迫下,噴施ABA 500 μmol·L-1與未噴施ABA處理相比Gs顯著上升(P<0.05),Pn和WUE分別上升85.1%和89.5%,降低葉片的失水速度,提高葉片的抗氧化能力,減緩因干旱脅迫造成的傷害。在重度干旱脅迫下噴施ABA 500 μmol·L-1能緩解干旱對多年生黑麥草光合作用的傷害。
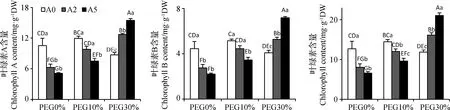
圖1 干旱脅迫下ABA對多年生黑麥草葉綠素a,b及葉綠素總量的影響
2.3 對葉綠素熒光的影響
干旱脅迫對多年生黑麥草Fo,Fm,Fv,Fv/Fm,Fv/Fo和NPQ均有著顯著影響(表1,2)。與CK相比,重度干旱脅迫(PEG 30%)造成了光抑制現象,使Fv/Fm和Fv/Fo分別下降2.6%和19.4%(P<0.05);NPQ上升97.3%(P<0.05),ABA對正常生長的多年生黑麥草Fv/Fm和Fv/Fo有顯著影響(P<0.05)。但在干旱脅迫下外源ABA在Fo,Fm,Fv,Fv/Fm,Fv/Fo和NPQ方面存在的交互作用(P<0.05),即噴施外源素ABA影響了干旱對光反應中心PSII的作用。
中度干旱脅迫下表現出較強的抗逆性,隨著噴施ABA濃度的升高Fv/Fm和Fv/Fo下降;中度干旱下噴施ABA 200 μmol·L-1能提高電子傳遞速率,而噴施ABA 500 μmol·L-1則能夠緩解對葉綠體光合機構的破壞。
重度干旱下噴施ABA 500 μmol·L-1能緩解干旱對多年生黑麥草的傷害,與未噴施ABA處理相比,Fv/Fo顯著升高21.1%(P<0.05)PSII電子傳遞活性升高,植物光合活性的水平上升,葉片利用光能的能力增強。
3 討論與結論
干旱是不可避免的自然災害,在自然界植物會遭受周期性或長期性干旱脅迫[24-25]。受到嚴重干旱脅迫后,植物在形態結構、生理代謝和產量都會受到不同程度的影響[26-28],從而直接或者間接影響到光合色素及光合特性。本研究中,多年生黑麥草在中度干旱脅迫下葉綠素含量出現了升高,相反,重度干旱脅迫下葉綠素降低,這與干旱脅迫對甘蔗葉綠素的影響結果一致[29],也進一步證明了張艷俠[30]對花生的研究結果。葉綠體是植物光合作用的主要場所,干旱脅迫影響了植物進行光合作用,使植物的葉綠體結構造成破壞導致葉綠素含量降低[31]。多年生黑麥草在重度干旱下噴施ABA 200 μmol·L-1和500 μmol·L-1提高葉綠素的含量,使葉綠素a、葉綠素b和總葉綠素上升。重度干旱脅迫下噴施ABA可以防止葉綠素降解引起的最大光能轉化效率有明顯的緩解作用[12,32]。
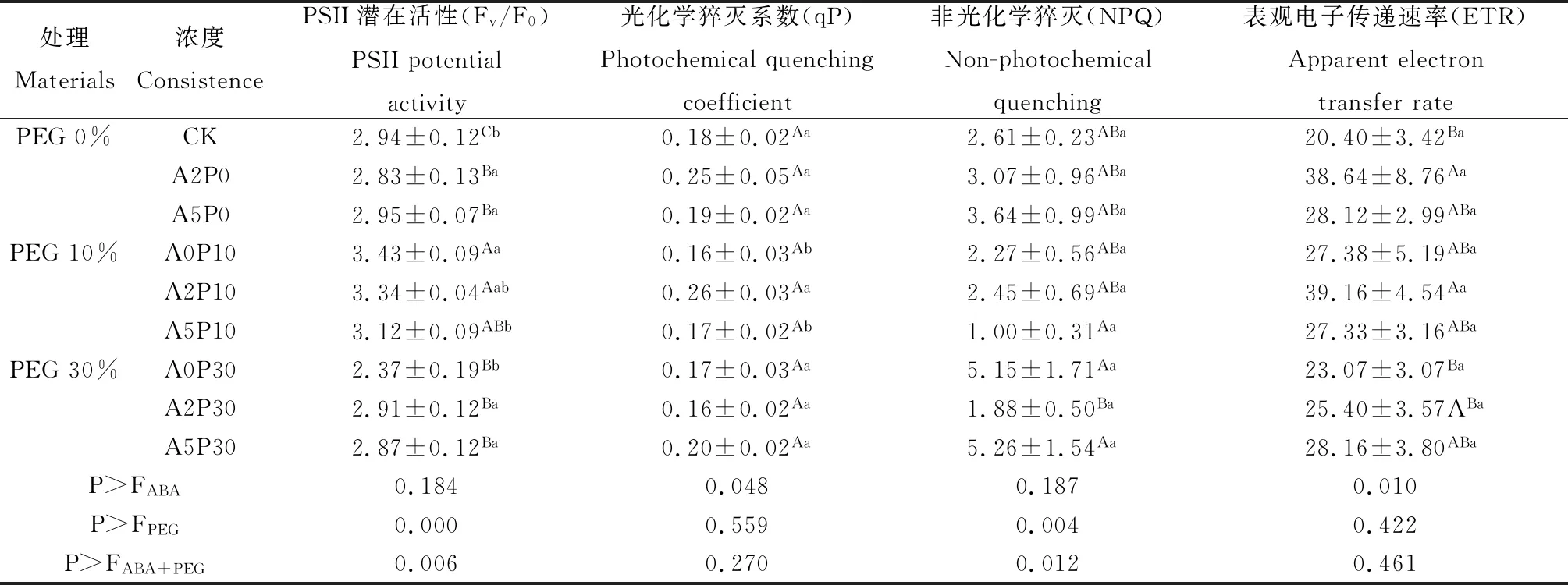
表2 干旱脅迫下ABA對多年生黑麥草Fv/Fo,qP,NPQ和ETR的影響
干旱導致的水分脅迫使得植物光合作用減弱,凈光合速率(Pn)降低[33],導致植物減產[34-35]。氣孔導度影響著植物的蒸騰和光合作用,氣孔導度降低,葉綠體光合能力受到明顯抑制[36],氣孔部分關閉或全部關閉[13],降低蒸騰速率(Tr),減少葉片內水分蒸發,CO2的進入阻力增大,胞間CO2濃度(Ci)上升,光合速率下降。
多年生黑麥草作為冷季型草坪草屬于C3植物,CO2補償點較高,胞間CO2增加并不是因為光合作用限制,而是受非氣孔因素的限制,這與顧振瑜等[37]研究結果一致。王書宏等[38]研究發現外源ABA能夠顯著提高干旱脅迫下草莓(Fragaria×ananassaDuch.)葉片的凈光合速率(Pn),緩解干旱造成的光抑制現象。本研究結果表明中度干旱脅迫下噴施ABA 200 μmol·L-1能夠提高多年生黑麥草的凈光合速率(Pn)和水分利用效率(WUE),氣孔導度(Gs)和胞間CO2濃度(Ci)下降,這與陳露露[39]和郭貴華[40]等研究一致。試驗還發現多年生黑麥草光合參數隨著干旱程度的加深和ABA濃度增加而逐漸上升。但不同強度或類型的干旱脅迫可能導致不同程度或方式的適應性反應[41],由于植物存在差異性,ABA也是一種生長抑制物質,關于干旱脅迫程度和外源ABA濃度的關系還需要進一步研究。在正常生長的植物上,噴施ABA效果與干旱脅迫下植物受到的影響有所不同。在沒有干旱脅迫下,噴施ABA 200 μmol·L-1,多年生黑麥草水分利用效率(WUE)提高,胞間CO2濃度(Ci)降低,這與李雪梅對小麥[42]研究一致。相反,噴施ABA 500 μmol·L-1,葉片光合作用受到了抑制。但是,劉小金等[43]對檀香幼苗的研究發現葉面噴施適量的ABA能夠提高檀香(SantalumalbumL.)幼苗的凈光合速率(Pn),氣孔導度(Gs)和蒸騰速率(Tr)上升,而水分利用效率(WUE)下降;隨著ABA濃度的升高氣孔導度(Gs)和蒸騰速率(Tr)提高,而水分利用效率(WUE)顯著降低。
葉綠素熒光參數能夠準確檢測植物的光合機構在逆境下的響應能力,從而反映植物的抗逆性[44]。裴斌等[45]發現干旱導致葉片初始熒光上升,而最大熒光、最大光化學效率和實際光化學效率逐漸降低。PSII原初光能轉化效率可以用葉綠素熒光參數Fv/Fm反映,干旱脅迫可導致植物的Fv/Fm和Fv/Fo降低[46]。Fo的增長越小,干旱脅迫對反應中心破壞程度越小,植物的抗旱性越強[47]。在干旱脅迫下,多年生黑麥草Fv/Fm和Fv/Fo呈下降趨勢,說明干旱對植物葉片造成了光抑制作用,影響光合電子的傳遞過程,與王飛[48]、代微然[49]研究結果一致,在逆境脅迫下或植物遭受到傷害時Fv/Fm會顯著降低[50]。
若Fo上升,表明PSII光合機構受到了損壞或者是植物本身一些防御性的激發能熱耗散過程加強的反映[51-53],多年生黑麥草在干旱脅迫下Fo變化顯著,表明干旱脅迫對PSII反應中心的破壞較大,且隨著脅迫的加劇,其光合功能的破壞程度增加,這進一步證明了胡化廣[54]和吳甘霖等[55]的研究結論。
另外,干旱脅迫使qP和ETR呈下降趨勢,非光化學猝滅系數(NPQ)是光合機構的一種自我保護機制,NPQ上升從而降低PSII活性,從而適應干旱環境,這與諶端玉[56]、史勝青[57]等研究結果一致。多年生黑麥草在干旱脅迫下,表觀電子傳遞速率(ETR)下降,可以有效緩解因PSII吸收太多的光能而產生的光抑制和光氧化[58]。在重度干旱脅迫下,噴施ABA 500 μmol·L-1對Fv/Fm和Fv/Fo沒有產生顯著影響,說明當干旱脅迫超過一定程度噴施ABA并不能改變對植物光合器官的傷害,產生光抑制現象。NPQ上升光合作用受到抑制,影響植物進行電子傳遞。
ABA對正常生長的多年生黑麥草的葉綠素含量產生抑制作用,但在重度干旱脅迫下噴施ABA能夠提高葉綠素含量并緩解干旱脅迫造成的影響,從而發揮促進作用。本研究中ABA濃度不同產生的影響不同,在大田進行ABA噴施時要掌握好噴施的濃度,以免造成不必要的損失,從而更加有效的緩解干旱對植物的傷害。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
