香根草對鎘毒害的生理響應
——采用傅里葉變換紅外光譜法(FTIR)
2019-11-07 06:55:18余順慧陳華華胡超生鄧洪平
生態學報 2019年19期
關鍵詞:植物
余順慧,張 靜,陳華華,張 波,胡超生,鄧洪平
1 重慶三峽學院,三峽庫區水環境演變與污染防治重慶高校市級重點實驗室, 萬州 404100 2 重慶市萬州區生態環境監測站, 萬州 404100 3 重慶市交通規劃勘察設計院,重慶 401121 4西南大學生命科學學院,三峽庫區生態環境教育部重點實驗室,重慶 400715
工業排污,農業耕作等生產活動造成的土壤重金屬污染問題日益嚴重。目前,我國重金屬污染土壤總面積已達2×107hm2,占我國耕地總面積的1/5[1]。其中,重金屬鎘(Cd)是毒性最強的元素之一,易溶于水、通過食物鏈,對人體健康構成巨大威脅。如何去除土壤中的重金屬Cd已成為人們關注的熱點。傳統的物理、化學修復方法存在成本高、破壞土壤理化性質、易造成二次污染等弊端。植物修復(Phytoremediation) 是利用植物來轉移、容納或轉化重金屬污染物使其對環境無害的技術,具有成本低,不破壞生態環境等優點。這已經成為土壤、水體重金屬污染研究領域的一項新興技術。
高水平的重金屬污染能引起植物細胞結構的傷害甚至致死。但在重金屬脅迫早期,當脅迫沒有產生生理癥狀時,在細胞和組織內的主要物質如蛋白質、糖含量、構象和分子內相互作用等方面都會發生變化,而這些變化一般通過測定可溶性糖、可溶性蛋白質含量及SOD、POD和CAT酶的活性等指標來進行研究。但測定這些生理指標的方法不但操作復雜、試劑花費較多,而且不能解析蛋白質等大分子物質結構上的變化[2]。傅立葉變換紅外光譜(FTIR)是一種基于化合物中官能團和極性鍵振動的結構分析技術[3],其特點是操作簡單、快速及靈敏度高。目前國內學者已經應用FTIR技術研究植物對重金屬的耐性機制[4- 6]。薛生國等采用FTIR法探討紫茉莉對Pb脅迫生理響應的影響,發現紫茉莉通過根系有機物含量的變化,將植物吸收的Pb 大部分積累在根部,阻止Pb向地上部分運輸,有效的保護了植物地上部分[7]。付川等應用FTIR技術研究紫花苜蓿對Cu脅迫生理響應,認為植物化學組分的變化有助于揭示超富集植物的Cu耐性機理[8]。但國內外研究多集中在生物量小,根系不夠發達的植物類群[9]。香根草(Vetiveriazizanioides) 為禾本科多年生草本植物,是三峽庫區的原生物種,具有生物量大、根系發達、抗逆性強等特點,并且對重金屬鉛、鎘具有較強的耐性和富集能力,可作為三峽庫區土壤重金屬污染修復的備選植物[10]。由此,探討不同Cd處理條件下其化學組成上的變化將有利于揭示其耐Cd機制。本研究擬利用FTIR技術,探討不同Cd處理條件下,香根草根、莖、葉的化學組分變化和毒害機理,為三峽庫區重金屬Cd污染防治和土壤植物修復提供理論依據。
1 材料及方法
1.1 試驗材料及方法
本研究以重慶三峽學院百安校區實驗區的香根草為試驗對象。2016年5 月隨機選取健壯的、生長較為一致的香根草分蘗苗。先用水洗凈,后移栽到含有Hoagland完全培養液的200 mL錐形瓶中進行水培純化7 d。當新芽長出后,將植株移入含有不同Cd2+濃度(0、1.0、3.0、5.0、7.0 mg/L)的營養液中進行繼續培養。Cd2+以3CdSO4·8H2O形式加入,每個濃度處理設3瓶。每3 d換一次培養液,24 h連續通氣,所有處理培養液的pH值維持在(6.5±0.1)。若瓶內液體量減少,則滴加含Cd2+營養液至初始刻度線;若瓶內液體變渾濁,則更換帶有Cd2+的營養液。處理10 d后收獲香根草植株。用自來水和去離子水洗凈,再分成兩部分,一部分取葉保存于液氮中備用,用于測定植株亞細胞組分。另一部分分別取根、莖、葉3部分于105 ℃烘箱殺青30 min,然后于80 ℃烘箱烘中干燥48 h,用鋼磨碎機研磨成粉末,過200目篩。用于測定植株根、莖、葉樣品的光譜信息。
1.2 傅里葉變換紅外光譜(FTIR)分析
來自于植物各部分的干粉用KBr混合(比例1.5∶100)。用傅里葉紅外光譜儀(日本島津制造IRPrestige-21,光譜范圍4000—400 cm-1,分辨率16 cm-1)測定香根草根、莖、葉樣品的光譜信息。
1.3 Cd2+亞細胞分布的測定
提取各細胞器和細胞質組分參照湯恵華等[11]方法。取嫩葉2.000 g,按1∶10的比例,加入預冷的勻漿液〔250 mmol/L蔗糖,50 mmol/L Tris-HCl(pH 7.4),1 mmol/L二硫赤蘚糖醇(C4H10O2S2)〕在4 ℃下研磨成勻漿。勻漿液先后在798 r/min離心10 min,1336 r/min離心15 min,13361 r/min離心20 min,先后得沉淀為細胞壁、葉綠體、線粒體組分,最后上清液為細胞質組分(含液泡及核糖和蛋白質等)。各組分供Cd2+含量測定。
1.4 微波消解樣品測定其重金屬Cd2+含量
精確稱取各樣品0.2000 g,加入酸液(濃HNO3∶濃H2SO4=4∶1),同時做空白組;將以上溶液在MARS240微波消解系統消解后,冷卻。過濾,濾液在25 mL容量瓶中定容,移入塑料瓶中保存,用AA- 6300原子吸收分光光度計測定樣品中Cd2+含量(mg/kg干重)[12]。
1.5 計算和數據分析
富集系數 (BF)= 植物地上或地下部分鎘含量/營養液中鎘含量
(1)
轉運系數(TF)= 植物地上部分鎘含量/根中鎘含量
(2)
實驗數據采用IBM SPSS 20進行統計分析和處理,并應用Origin 8.0 進行紅外光譜數據圖譜處理。
2 結果與討論
2.1 Cd2+ 脅迫對香根草重金屬分布的影響
與對照相比,隨著Cd2+濃度的增大,香根草地下部分、地上部分Cd2+的含量逐漸增加;Cd2+處理后香根草的地下部分Cd2+含量明顯大于地上部分(表1)。當用最高Cd2+濃度7 mg/L處理時,此時香根草出現受害癥狀,表現為幼苗矮化,根長、根體積減小等。按照李東旭等[13]提出的參考值,超富集植物的轉運系數和富集系數都大于1。本實驗香根草的轉運系數和富集系數都小于1,說明香根草不屬于超富集植物。但香根草生物量大、根系發達、抗逆性強。且具有較強的Cd耐性,可長時間適應低濃度Cd 脅迫,甚至是短時間的高濃度Cd脅迫[14]。香根草能吸收Cd 并儲存在體內,具有較強的穩定能力,在Cd 污染土壤的修復方面有一定應用潛力[1],香根草作為一種植物修復的候選物種。
表1 Cd2+脅迫對香根草植物體內的積累和分布情況
Table 1 Accumulation and distribution of cadmium inV.zizanioidestissues and translocation factor in cadmium-treated solutions for 10 days
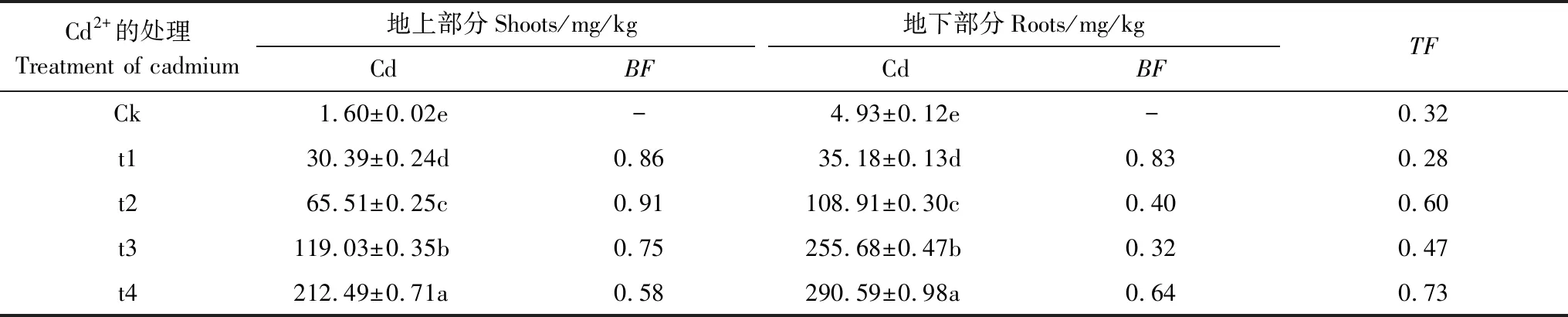
Cd2+的處理Treatment of cadmium地上部分Shoots/mg/kg地下部分Roots/mg/kgCdBFCdBFTFCk1.60±0.02e-4.93±0.12e-0.32t130.39±0.24d0.8635.18±0.13d0.830.28t265.51±0.25c0.91108.91±0.30c0.400.60t3119.03±0.35b0.75255.68±0.47b0.320.47t4212.49±0.71a0.58290.59±0.98a0.640.73
同列具有不同字母上標者為差異顯著(P<0.05); Ck: 對照 (0 mg/L); t1: 1 mg/L; t2: 3 mg/L; t3: 5 mg/L; t4: 7 mg/L; Cd: 鎘;BF:富集系數;TF: 轉運系數
2.2 Cd2+ 在香根草葉片內的亞細胞分布
Cd2+在香根草葉片內的亞細胞組分間分布不均勻,其中分配比例最高的是在細胞質,細胞壁次之,含量最少的是在葉綠體和線粒體(表2)。隨著Cd2+濃度的增高,葉片Cd2+總含量及各組分含量增加。當Cd2+濃度<3 mg/L時,Cd2+在細胞壁和細胞質中的分配比隨Cd2+處理濃度升高而升高,而當Cd2+濃度>3 mg/L時,Cd2+在細胞壁中的分配比例隨Cd2+處理濃度增高而降低,在細胞質中Cd2+的分配比例升高幅度也漸漸減少。可能是細胞質貯存Cd2+的能力有限。
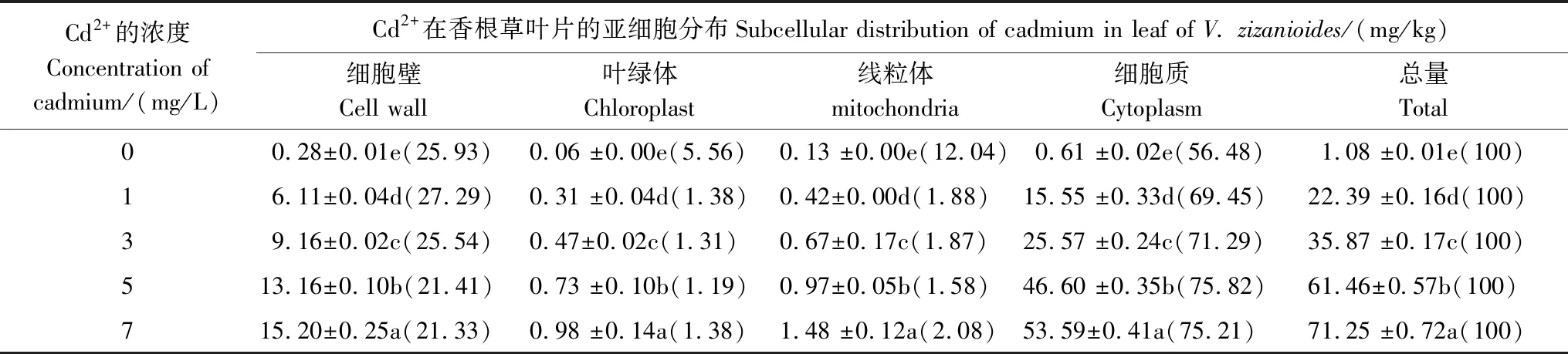
表2 Cd2+在香根草葉片的亞細胞分布
1)同列具有不同字母上標者為差異顯著(P<0.05);2)在括號里的數據為分配比例(%)
研究重金屬的亞細胞分布是為了確定該元素對細胞活動的影響[12]。本試驗中,細胞質是Cd2+主要貯存部位。細胞質包含液泡、細胞液、蛋白質等成分,大量貯存在香根草細胞質中的Cd2+是否被隔離在液泡中,以減少其毒性,提高香根草對Cd2+的耐受性,還有待進一步研究。細胞壁是重金屬進入細胞內部的第一道屏障,細胞壁中的糖、蛋白質及有機酸等可與金屬離子結合而貯存部分重金屬,減少重金屬對植物的毒害[12],這與后面FTIR研究中根、莖一致。Cd2+在葉綠體和線粒體含量最少,降低了Cd2+與有生命活性的原生質部分的接觸機會,使酶的活性不受影響,保證了光合作用和呼吸作用的正常進行,這可能是香根草對Cd2+具有一定耐受性的原因[15]。
2.3 不同濃度Cd2+處理對香根草根、莖和葉的FTIR分析

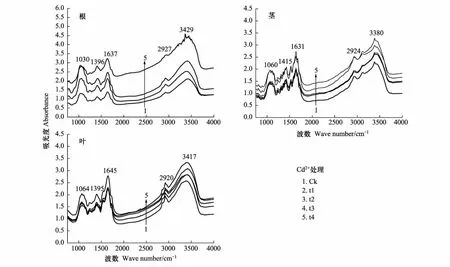
圖1 Cd2+處理下香根草根、莖、葉的紅外光譜圖Fig.1 Fourier Transform Infrared spectra of root, stem, leaf of V. zizanioides by cadmium treatment Ck: 對照 (0 mg/L); t1: 1 mg/L; t2: 3 mg/L; t3: 5 mg/L; t4: 7 mg/L
2.4 香根草特征峰吸光度變化的定量分析
對不同濃度重金屬處理前后的香根草幼苗的根、莖、葉進行了紅外光譜分析, 結果顯示各組分在重金屬處理前后的峰形基本保持不變。
由香根草的特征峰吸光度變化圖2可見,香根草幼苗根、莖、葉紅外光譜特征峰的吸收強度隨Cd2+濃度的升高而表現出不同的趨勢。根的特征峰吸光度變化為先降后升(見圖2)。在3380—3429 cm-1,2920—2927 cm-1,1631—1645 cm-1,1395—1415 cm-1,1030—1064 cm-1等處,香根草幼苗根在Cd2+濃度低于3.0 mg/L時,該峰的吸光度下降,可能根外表皮細胞壁的羥基吸附結合Cd2+形成穩定的化合物,使細胞表面的羥基減少[7]、植物根系不斷分泌有機酸螯合Cd2+,致使羧酸譜帶下降、蛋白質二級結構中,肽鍵間氫鍵的結合力隨著Cd2+濃度的升高而變弱、細胞壁通過降低果膠甲基化程度,使CEC(陽離子交換能力)提高,從而吸收更多的Cd2+,即通過細胞壁中積累Cd2+來增強抗逆性等。當Cd2+濃度大于3.0 mg/L時,峰值又呈上升趨勢,原因是高濃度的Cd2+可能破壞了根外表皮細胞壁的羥基吸附結合Cd2+的機制,導致根外表皮細胞壁的羥基無法結合Cd2+,隨著Cd2+對香根草毒害的進一步加重,其羧酸螯合力變弱,頻帶開始上升、此外不斷增加的Cd2+誘導富脯氨酸蛋白、病害相關蛋白和富甘氨酸蛋白等一些蛋白合成[7],Didierjean等[20]還認為這些重金屬脅迫誘導蛋白可能具有保護植物細胞免受重金屬毒害的作用,出現的顯著吸收峰可能與香根草根中氨基酸、多肽和蛋白質類物質含量升高等有關[7]。
香根草莖的特征峰吸光度變化為先升后降趨勢(圖2)。在低濃度時,所有香根草莖的特征峰吸光度隨著Cd2+濃度的升高,先升高(圖2),原因是低Cd2+下促進了有機物如纖維素、半纖維素、多糖等的分泌和運輸及氨基酸、多肽和蛋白質等物質的產生,通過滲透調節來增強抗逆性,而在低濃度Cd2+刺激下,對峰值變化的主要貢獻是膜脂過氧化作用,隨著Cd2+濃度的升高,膜過氧化程度加深,脂肪族酮類化合物(過氧化產物)在根部積累,引起峰值升高,當Cd2+濃度大于3.0 mg/L時,吸光度下降,表明隨著Cd2+毒的加劇,碳水化合物等物質的合成和運輸都降低、蛋白質二級結構中肽鍵間氫鍵的結合力受影響較大。
香根草葉的特征峰吸光度變化也是先升后降趨勢(圖2)。由圖2看出,在低Cd2+濃度時,所有香根草葉的特征峰吸光度隨著Cd2+濃度的增加而上升,先上升的原因是低Cd2+刺激下促進了碳水化合物及有機物的分泌和運輸,通過滲透作用來加強葉對Cd2+的抗逆性 、由根系輸送來的羧酸鹽較多,并以該形式積累在葉片和葉中蛋白質、氨基酸和多肽類物質含量增加。當Cd2+濃度大于3.0 mg/L時,吸光度降低,表明隨著Cd2+毒的加劇,Cd2+阻礙了這些碳水化合物、有機物及葉中蛋白質、氨基酸和多肽類物質的合成和運輸、根系羧酸螯合力變弱,其羧酸鹽的運輸能力也變弱等,曲線下降。
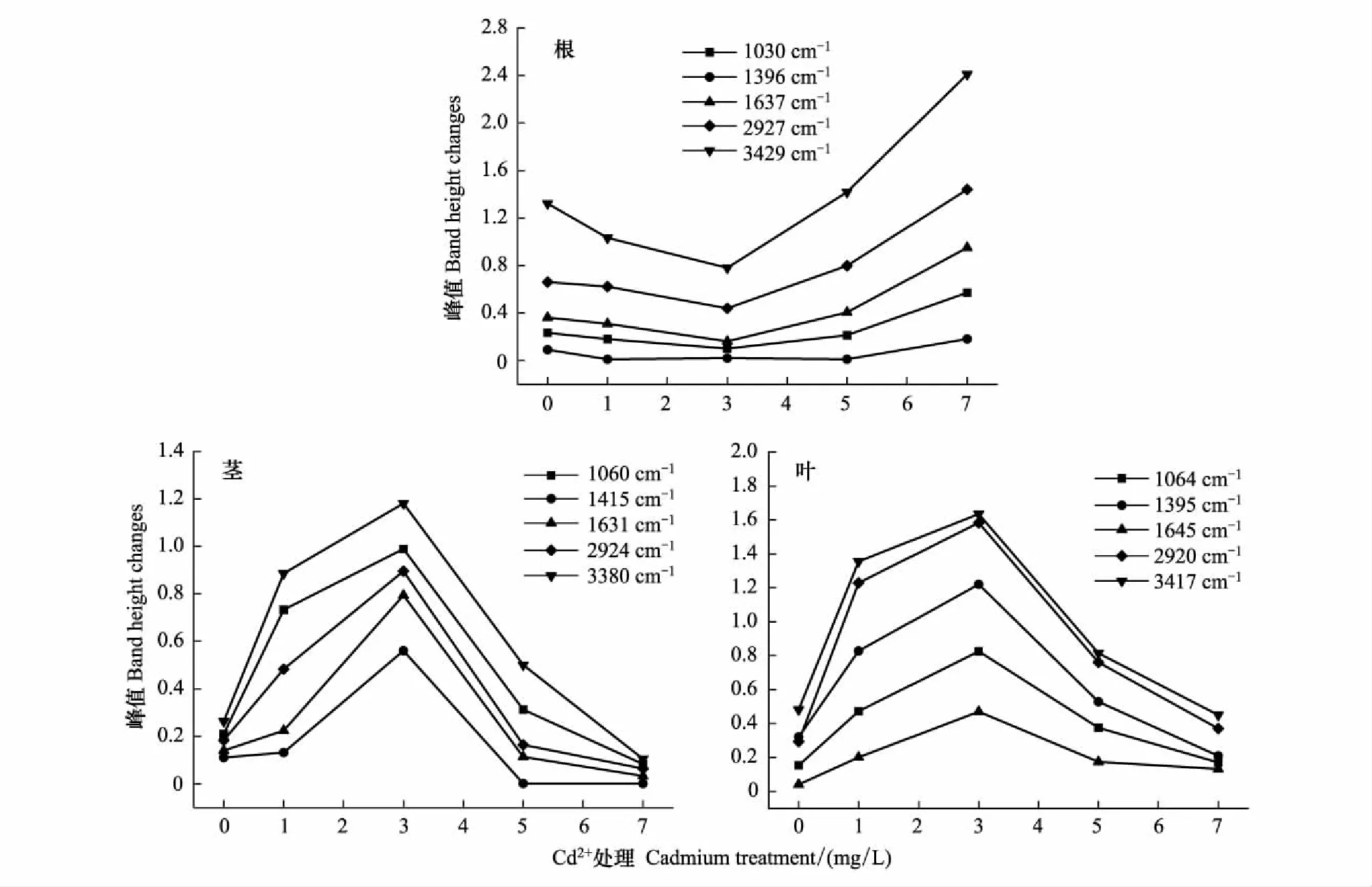
圖2 Cd2+處理下香根草根、莖、葉的特征峰吸光度變化圖Fig.2 The band height changes of root, stem and leaf in V. zizanioides by cadmium treatment 1030—1064 cm-1: 醇、酯基、醚基或酚類等化合物的C—O鍵的伸縮振動吸收峰;1395—1415 cm-1:甲基的吸收帶;1631—1645 cm-1: N—H酰胺的彎曲振動峰;2920—2927 cm-1:羧酸O—H與甲基飽和C—H鍵的伸縮振動重疊峰;3380—3429 cm-1:分子間氫鍵O—H自由羥基的伸縮振動峰
3 結論
(1)香根草根組織在2927 cm-1處峰高先下降后上升,表明在低Cd2+(Cd2+<3 mg/L)處理條件下香根草分泌的有機酸不斷螯合Cd2+,造成羧酸O—H的減少,但隨著Cd2+含量的升高,其羧酸螯合力變弱,有機酸含量漸漸升高。
(2)香根草莖組織在1631 cm-1峰高處先上升后下降,表明在低Cd2+(Cd2+<3 mg/L)處理條件下香根草產生氨基酸、多肽和蛋白質等物質,通過滲透調節來增強抗逆性,但隨著Cd2+含量的升高,蛋白質二級結構中肽鍵間氫鍵的結合力受影響較大
(3)香根草葉片富集的Cd2+主要分布于細胞質中,其次為細胞壁,而在線粒體和葉綠體中含量最低。
(4)雖然香根草對Cd2+的積累還未到達超富集植物的范圍,但從耐Cd2+能力、地上部分對Cd2+的富集量、高生物量、生長快等多方面綜合考慮,香根草在遭受Cd2+污染的土壤中植物修復技術領域,擁有潛在的利用價值。
致謝:東南大學王大勇教授對寫作給予幫助,特此致謝。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
