萱草葉枯病菌粗毒素液對(duì)分化芽生理生化代謝的影響
2019-10-25 02:28:26楊麗莉常建忠
山西農(nóng)業(yè)科學(xué) 2019年10期
關(guān)鍵詞:生長(zhǎng)
楊麗莉,楊 睿,常建忠,蔣 丹
(山西省農(nóng)業(yè)科學(xué)院旱地農(nóng)業(yè)研究中心,山西太原030031)
萱草(Hemerocallis hybrida)是百合科萱草屬多年生宿根植物,國(guó)內(nèi)本土大花萱草通常易感染銹病和炭疽病菌。國(guó)外引進(jìn)的大花萱草新品種[1]特別易感染一種新型病菌——萱草葉枯病(Kabatiella microsticta)[2],主要為害葉片,形成病斑,并不斷蔓延,尤其對(duì)紅運(yùn)萱草的危害達(dá)到70%以上。針對(duì)萱草葉枯病的致病菌國(guó)內(nèi)外學(xué)者已找到了病菌敏感的殺菌劑[2-4]。但是,園林綠化大面積噴施殺菌劑勢(shì)必會(huì)對(duì)環(huán)境造成污染,同時(shí)加大了園林的管理難度。因此,最有效的方法是對(duì)其進(jìn)行抗葉枯病性狀改良,提高其抗病性。
研究葉枯病菌毒素液對(duì)深入了解病原菌致病機(jī)制、利用毒素就寄主抗病性進(jìn)行快速檢測(cè)、篩選抗病材料是十分重要的[5]。本試驗(yàn)就萱草葉枯病菌毒素液對(duì)紅運(yùn)萱草愈傷組織分化芽生長(zhǎng)的影響和防御酶(SOD、POD、CAT)活性變化進(jìn)行了研究,旨在為探索葉枯病發(fā)生機(jī)制和利用病菌毒素液進(jìn)行抗病突變體篩選提供依據(jù)。
1 材料和方法
1.1 材料
1.1.1 供試萱草苗 從國(guó)外引進(jìn)分株2~3 年生易感染葉枯病的品種紅運(yùn),在山西省農(nóng)業(yè)科學(xué)院東陽(yáng)試驗(yàn)示范基地種植。
1.1.2 供試菌株 萱草葉枯病菌(Kabatiella microsticta Bubak(syn.Aureobasidium microstictum(Bubak)W.B.Cook)),由吉林農(nóng)業(yè)大學(xué)植物病理教研室分離鑒定并保存。
1.1.3 保護(hù)酶測(cè)定試劑盒 采用南京建成生物工程研究所生產(chǎn)的試劑盒。SOD 試劑盒(測(cè)總),羥胺法(貨號(hào)為A001-1);POD 試劑盒(植物,貨號(hào)為A084-3);CAT 可見(jiàn)光試劑盒(貨號(hào)為A007-1)。
1.2 方法
1.2.1 葉枯病菌毒素培養(yǎng) 受體材料為大花萱草紅運(yùn)的愈傷組織分化芽。紅運(yùn)的組織培養(yǎng)為專利技術(shù)[6]。
1.2.2 葉枯病菌粗毒素液的制備和毒力檢測(cè) 其參照文獻(xiàn)[7]進(jìn)行。
1.2.3 葉枯病菌粗毒素液培養(yǎng)試驗(yàn) 受體為大花萱草紅運(yùn)愈傷組織繼代培養(yǎng)分化出的1~2 cm 的分化芽。繼代培養(yǎng)基中分別添加濃度20%,40%,60%的病菌毒素液,分別標(biāo)記為1,2,3 號(hào),同時(shí)以不添加毒素液為對(duì)照(CK),高壓滅菌后備用。選取大小基本一致的萱草分化芽,剪去上部葉片,每瓶8 株,每處理3 瓶,3 次重復(fù)。(25±1)℃,16 h 光/8 h暗,光照強(qiáng)度2 800 lx。觀察生長(zhǎng)狀況。
1.3 測(cè)定項(xiàng)目及方法
1.3.1 生長(zhǎng)量指標(biāo)的測(cè)定 分別在分化芽的第0,8,16,24,32 天稱其鮮質(zhì)量,繪制生長(zhǎng)量曲線。調(diào)查成活數(shù),計(jì)算存活率。

1.3.2 保護(hù)酶活性的測(cè)定 采用南京建成生物工程研究所試劑盒法測(cè)定保護(hù)酶活性。在處理前和處理后第2,7,14,21 天分別進(jìn)行保護(hù)酶活性的測(cè)定。保護(hù)酶提取液制備和測(cè)定均按試劑盒標(biāo)準(zhǔn)進(jìn)行。以雙蒸水為對(duì)照,3 次重復(fù)。分別計(jì)算總SOD、POD 和CAT 酶活力。
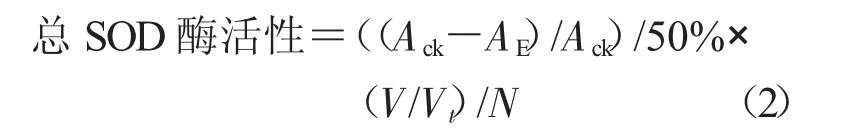
式中,總SOD 酶活性為每克組織在1 mL 反應(yīng)液中SOD 抑制率達(dá)50%時(shí)所對(duì)應(yīng)的SOD 量為一個(gè)SOD 活力單位(U)(U/g);Ack為對(duì)照OD 值;AE為樣品OD 值;V 為反應(yīng)液總體積(mL);Vt為取樣量(mL);N 為勻漿液濃度(組織鮮質(zhì)量(g)/勻漿介質(zhì)體積(mL),按1%計(jì))。
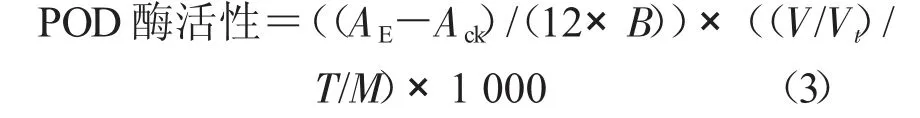
式中,POD 酶活性為37 ℃下,每毫克組織蛋白每分鐘催化1 μg 底物的酶量定義為一個(gè)酶活力單位(U/mg);Ack為對(duì)照OD 值;AE為樣品OD 值;B 為比色光徑(1 cm);V 為反應(yīng)液總體積(mL);Vt為取樣量(mL);T 為 反應(yīng)時(shí)間30 s;M 為勻漿蛋白濃度(mg/mL),以1.0 mg/mL 計(jì)算。
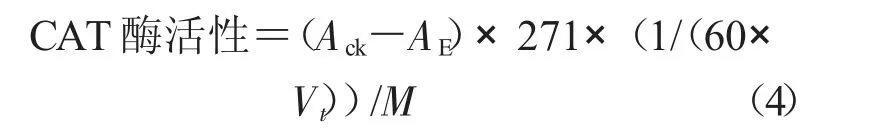
式中,CAT 酶活性為每毫克組織蛋白每秒鐘分解1 μmol H2O2的量為一個(gè)活力單位(U/mg);271 為斜率的倒數(shù);Ack為對(duì)照OD 值;AE為樣品OD 值;Vt為取樣量(mL);M 為勻漿蛋白濃度,以1.0 mg/mL計(jì)算。
1.4 數(shù)據(jù)處理與分析
數(shù)據(jù)處理采用Excel 2007 進(jìn)行,單因素方差分析采用SPSS 軟件進(jìn)行,采用LSD 法進(jìn)行平均值間多重比較。
2 結(jié)果與分析
2.1 葉枯病菌粗毒素液對(duì)分化芽生長(zhǎng)的影響
用添加不同濃度葉枯病菌粗毒素液的固體培養(yǎng)基對(duì)愈傷組織分化芽進(jìn)行培養(yǎng),結(jié)果發(fā)現(xiàn),病菌毒素液對(duì)分化芽均有不同程度的毒害作用,主要表現(xiàn)在葉片出現(xiàn)水漬型黃化、生長(zhǎng)緩慢和致死效應(yīng),并隨著培養(yǎng)時(shí)間的延長(zhǎng)毒害作用加劇。前3 d 各處理的毒害作用表現(xiàn)均不明顯;3 號(hào)樣品在7 d 左右時(shí)葉尖部開(kāi)始發(fā)黃,并向下蔓延;2 號(hào)和1 號(hào)則出現(xiàn)在10~15 d;隨著培養(yǎng)時(shí)間的延長(zhǎng),3 號(hào)整株失綠,生長(zhǎng)停滯,逐漸死亡;2 號(hào)和1 號(hào)的生長(zhǎng)狀態(tài)與3 號(hào)相似,但是毒害作用較輕,受傷害程度大小依次為3 號(hào)>2 號(hào)>1 號(hào)(圖1)。

從表1 可以看出,隨著毒素液濃度的增加抑制作用增強(qiáng)。毒素液濃度在20%(1 號(hào))時(shí),分化芽生長(zhǎng)勢(shì)比較旺盛;達(dá)到40%(2 號(hào))時(shí)生長(zhǎng)緩慢;達(dá)到60%(3 號(hào))時(shí),前16 d 時(shí)生長(zhǎng)速度雖然不及1 號(hào)和2 號(hào),但是還有緩慢生長(zhǎng),之后生長(zhǎng)趨于停滯狀態(tài)。從不同處理增長(zhǎng)量的方差分析結(jié)果可以看出,前8 d,1 號(hào)與CK 間沒(méi)有顯著差異,2 號(hào)和3 號(hào)與CK和1 號(hào)間在0.05 水平上差異顯著;到第32 天時(shí),1、2、3 號(hào)和CK 之間在0.05 水平差異均顯著。說(shuō)明病菌毒素液對(duì)分化芽生長(zhǎng)的影響是非常巨大的,濃度達(dá)到60%時(shí),分化芽生長(zhǎng)極其緩慢,甚至停止生長(zhǎng)。
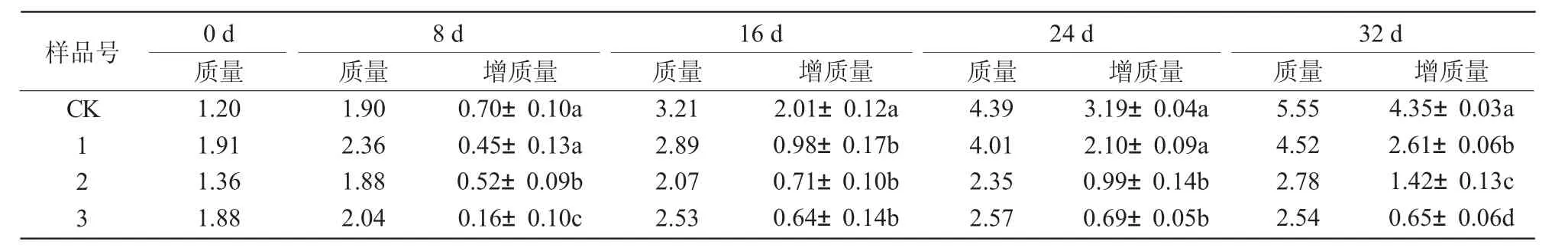
表1 葉枯病菌粗毒素液對(duì)分化芽生長(zhǎng)的影響 g
2.2 葉枯病菌粗毒素液對(duì)分化芽的致死作用
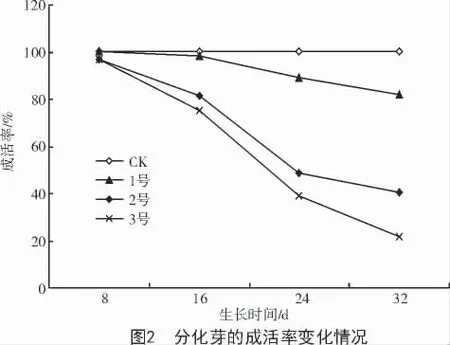
葉枯病菌粗毒素液對(duì)分化芽的生長(zhǎng)產(chǎn)生很大的抑制和傷害作用,進(jìn)一步影響到了成活率。從圖2 可以看出,毒素液對(duì)成活率的影響在前16 d 尚不明顯,16 d 后十分明顯。同一濃度下,隨著培養(yǎng)時(shí)間的延長(zhǎng),成活率有不同程度的下降。菌液濃度增加,分化芽成活率顯著下降,其中,菌液濃度在40%(2 號(hào))、生長(zhǎng)24 d 時(shí),成活率下降到了50%;32 d時(shí),降到40%左右。菌液濃度在60%(3 號(hào))時(shí),從16 d 起,成活率下降幅度明顯高于1,2 號(hào),在32 d時(shí)成活率只有20%左右。說(shuō)明20%濃度(1 號(hào))的菌液對(duì)分化芽成活率的影響不明顯,40%濃度(2 號(hào))時(shí)的死亡率達(dá)到40%~50%,60%濃度(3 號(hào))有強(qiáng)致死效應(yīng)。結(jié)合分化芽的生長(zhǎng)狀況分析得出,在培養(yǎng)基中添加病菌毒素液濃度為40%是一個(gè)篩選的合理劑量,培養(yǎng)時(shí)間為24 d 較適宜。
2.3 葉枯病菌粗毒素液對(duì)防御酶活性的影響
2.3.1 對(duì)SOD 酶活性的影響 SOD 催化O2-·歧化反應(yīng)生成O2和H2O2,其活性被認(rèn)為是抗逆境的重要指標(biāo)[8-9]。培養(yǎng)基中添加葉枯病菌粗毒素液對(duì)SOD酶活性均有不同程度的影響,導(dǎo)致分化芽SOD 酶活性呈現(xiàn)先升高后下降的趨勢(shì),升高的幅度與毒素液濃度呈正相關(guān)(圖3)。SOD 酶活性在第2 天達(dá)到最高值;7 d 內(nèi),3 號(hào)維持在最高水平,1,2 號(hào)緩慢下降,14 d 時(shí)下降到CK 水平;之后持續(xù)下降,在21 d時(shí),SOD 酶活性達(dá)到最低,1,2,3 號(hào)均低于CK,其中,3 號(hào)最低,其次依次為2 號(hào)和1 號(hào)。說(shuō)明分化芽接種到含病菌毒素液的培養(yǎng)基上,在毒素逆境條件作用下,分化芽細(xì)胞迅速啟動(dòng)SOD 酶防御系統(tǒng),酶活性迅速升高,清除O2-·自由基,并持續(xù)在較高水平。毒素液濃度越高,對(duì)細(xì)胞的毒害作用越強(qiáng),酶活性升高的幅度越大。隨著逆境時(shí)間的延長(zhǎng),細(xì)胞受毒害作用持續(xù)增強(qiáng),SOD 酶系統(tǒng)不足以清除植株中積累的大量超氧陰離子自由基,細(xì)胞逐漸衰老死亡,SOD 酶活性持續(xù)下降。在3 個(gè)處理中,1 號(hào)和2 號(hào)比3 號(hào)受傷害小。

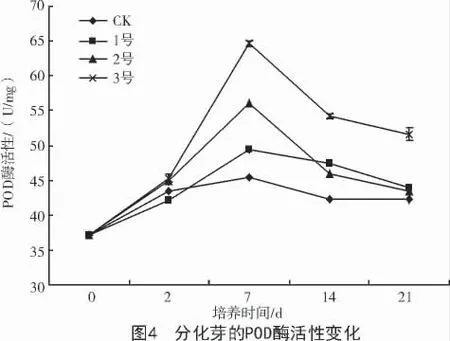
2.3.2 對(duì)POD 酶活性的影響 POD 是植物體內(nèi)普遍存在的氧化還原酶,它不僅參與木質(zhì)素、酚類物質(zhì)及植保素的合成,還作為整個(gè)代謝途徑的調(diào)節(jié)子,是細(xì)胞內(nèi)重要的內(nèi)源活性氧的清除劑,是公認(rèn)的與植物抗性相關(guān)的酶[9-10]。培養(yǎng)基中添加葉枯病菌粗毒素液分化芽POD 酶活性先升高后下降(圖4)。培養(yǎng)24 h,POD 酶活性略有升高,培養(yǎng)到第7 天病癥出現(xiàn)時(shí),酶活性才迅速升高,升高的幅度與添加的毒素液濃度呈正相關(guān),與接種大豆花葉病毒后的反應(yīng)相同[11]。隨著毒素液培養(yǎng)時(shí)間的延長(zhǎng),酶活性下降,下降的幅度與毒素液濃度呈負(fù)相關(guān)。在21 d時(shí),3 號(hào)酶活性最高,與1,2 號(hào)和CK 間顯著差異。說(shuō)明毒素濃度越高對(duì)植株的刺激作用越強(qiáng),細(xì)胞以提高POD 酶活性來(lái)應(yīng)對(duì)逆境環(huán)境,可能是細(xì)胞的應(yīng)激反應(yīng)和防御機(jī)制[12]。
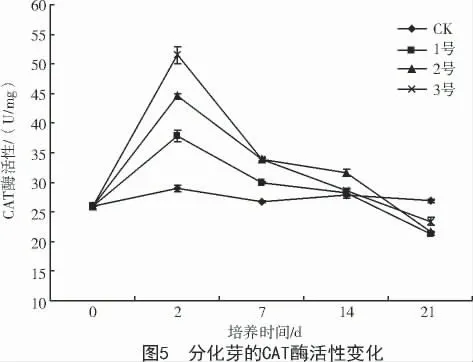
2.3.3 對(duì)CAT 酶活性的影響 CAT 存在于需氧的生物體內(nèi),在消除過(guò)氧化物酶體中的H2O2時(shí)起到重要作用[13]。培養(yǎng)基中添加葉枯病菌粗毒素液分化芽CAT 酶活性先升高后下降,升高的幅度與毒素液濃度呈正相關(guān)(圖5)。CAT 酶活性在第2 天達(dá)到峰值,各處理之間、各處理與CK 之間均呈顯著差異。之后酶活性下降,在21 d 時(shí)接近CK 水平。
3 結(jié)論與討論
本研究結(jié)果表明,葉枯病菌毒素液對(duì)大花萱草分化苗生長(zhǎng)具有一定的毒害作用,使得植株生長(zhǎng)量、存活率和保護(hù)酶活性等方面存在著明顯的劑量差異。病菌毒素液劑量低,毒害作用弱;劑量高,毒害作用強(qiáng)。這一結(jié)果對(duì)于室內(nèi)進(jìn)行抗病資源的篩選與鑒定具有十分重要的參考價(jià)值[14],20%~40%的劑量范圍是適合的篩選劑量。結(jié)合誘變育種進(jìn)行抗病突變體選擇壓篩選,獲得抗病突變體材料,對(duì)大花萱草抗葉枯病育種具有十分重要的意義。
參與植物體內(nèi)多種生理代謝過(guò)程的SOD、POD和CAT 以及PAL 和PPO 等酶系與植物抗性密切相關(guān)[15]。其中,SOD 酶是植物與病原物識(shí)別過(guò)程中產(chǎn)生初始抗性信息的一個(gè)關(guān)鍵酶,其主要功能是通過(guò)歧化反應(yīng)清除超氧陰離子自由基,因此,SOD 常作為抗性機(jī)制起作用[16];POD 酶屬于氧化酶系統(tǒng),是細(xì)胞內(nèi)重要的內(nèi)源活性氧的清除劑。有研究發(fā)現(xiàn),在病原菌染后,植物體內(nèi)細(xì)胞保護(hù)酶活性發(fā)生了相應(yīng)變化[11,17]。MONTALBINI 等[18]研究發(fā)現(xiàn),接種煙草花葉病毒(Tobacco mosaic virus,TMV)后,煙草葉片細(xì)胞SOD 酶活性明顯加強(qiáng)。朱友林等[19]在對(duì)玉米大斑病菌侵染玉米的研究中發(fā)現(xiàn),抗病品種POD 基因等防衛(wèi)基因較感病品種更快速被誘導(dǎo)表達(dá)。吳岳軒等[20]研究發(fā)現(xiàn),雜交稻感染白葉枯病菌后,葉片中POD 活性升高。本試驗(yàn)中,萱草分化芽在經(jīng)不同濃度的粗毒素液培養(yǎng)后,3 種酶的活性都有不同程度的變化,呈現(xiàn)不同程度的先升高后降低,活性升高的程度與毒素液的濃度呈正相關(guān),并顯示顯著差異,與接種病菌后的反應(yīng)是一致的。低濃度毒素液對(duì)分化芽的傷害作用較輕,濃度增加,傷害作用加劇。酶活性峰值出現(xiàn)的時(shí)間依防御酶的種類不同而不同。SOD 和CAT 酶活性峰值出現(xiàn)在第2 天,POD酶活性峰值出現(xiàn)在第7 天。因此,分化芽通過(guò)各級(jí)信號(hào)傳導(dǎo)激活防御酶系統(tǒng),啟動(dòng)與抗病性有關(guān)的基因得到表達(dá),從而應(yīng)對(duì)病菌毒素液的傷害。這一結(jié)果為利用病菌毒素液作為選擇壓篩選抗病突變體提供了理論依據(jù)。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
