T細胞免疫球蛋白和黏蛋白結構域蛋白3抑制銀屑病患者CD14+單核細胞分泌白細胞介素12
2019-10-17 00:39:46秦俊霞秦小衛趙鵬
中華皮膚科雜志 2019年9期
秦俊霞 秦小衛 趙鵬
山西省人民醫院皮膚科,太原 030012
銀屑病的發病與多基因遺傳背景下的細胞免疫異常密切相關,T 細胞、粒細胞、單核細胞、角質形成細胞、內皮細胞、神經細胞等以及這些細胞之間的復雜相互作用誘導銀屑病的發生發展[1]。T細胞免疫球蛋白和黏蛋白結構域蛋白3(T cell immunoglobulin and mucin domain-containing protein 3,TIM-3)是一類T細胞表面抑制性分子,在病毒感染、腫瘤等疾病過程中發揮介導免疫耐受的作用[2-3]。TIM-3 不但可調控T 細胞功能,還在多種抗原提呈細胞中高表達,調控單核巨噬細胞活化,參與機體固有免疫應答[4-5]。TIM-3 在銀屑病患者 Th1 和Th17 細胞中的表達降低,誘導并促進炎癥應答[6],但TIM-3 對銀屑病患者單核細胞的調控作用尚未見相關報道。本研究利用體外細胞培養系統,研究TIM-3對銀屑病患者CD14+單核細胞分泌細胞因子功能的調控作用,以期初步闡釋TIM-3對固有免疫的調控在銀屑病發病中的作用。
對象和方法
一、對象
2017年11月至2018年3月于山西省人民醫院皮膚科就診的尋常型銀屑病患者47例,入組標準:年齡≥18 歲且符合《中國銀屑病治療專家共識(2014版)》診斷標準[7]。排除標準:合并惡性腫瘤、合并自身免疫性疾病、合并慢性病毒感染或者長期接受免疫抑制劑治療。其中,男21 例,女26 例,年齡18 ~ 57(41.8 ± 13.9)歲,銀屑病面積和嚴重程度指數(psoriasis area and severity index,PASI)評分4 ~ 29(8.12± 3.37)分。19例年齡和性別相匹配的健康體檢者作為對照組,其中男8例,女11例,年齡18 ~ 65(39.7 ± 10.9)歲。本研究方案已通過山西省人民醫院倫理委員會審核[(2017)省醫倫審字19號],所有入組志愿者均簽署知情同意書。
二、方法
1.外周血單個核細胞(peripheral blood mononuclear cell,PBMC)的分離:清晨空腹留取外周血20 ml,EDTA 抗凝。使用Ficoll 淋巴細胞分離液(美國Sigma 公司),采用密度梯度離心法分離PBMC,凍存于液氮中備用。
2.流式細胞儀檢測CD14+細胞TIM-3的表達和IL-12的分泌:每例受試者取106個PBMC,設立2個復孔,每個復孔中加入5 × 105個PBMC,加入含10%胎牛血清的RPMI 1640培養液于37 ℃、5%CO2條件下培養,其中1 個復孔加入Toll 樣受體4(Tolllike receptor 4,TLR4)配體脂多糖(lipopolysaccharide,LPS;美國 Invivogen 公司,終濃度為 5 mg/L)和TLR7/8 配體 R848(美國 Invivogen 公司,終濃度為5 mg/L)刺激培養12 h,在刺激培養的后6 h加入布雷非德菌素A(美國BD公司,終濃度為10 mg/L)抑制蛋白轉運,另1個復孔不加刺激因子。培養后收集細胞,轉入FACS 管中,加入抗CD14-APC(美國eBioscience 公司)和抗 TIM-3-FITC(美國eBioscience公司)進行細胞表面染色,加入抗IL-12-PE(美國eBioscience 公司)進行細胞內染色。使用FACS AriaⅡ流式細胞儀檢測,CellQuest Pro軟件獲取細胞,FlowJo Version 8.7.2軟件進行數據分析。
3.角質形成細胞的分離和培養:取10 例尋常型銀屑病患者皮損標本置于預冷的0.2%胰酶中冷消化12 h,然后37 ℃熱消化30 min,待真表皮分離時轉入裝有含10%胎牛血清的RMPI 1640 培養液無菌平皿中沖洗,分離表皮,放入離心管中反復吹打,過100目尼龍篩,獲取細胞,然后以105/cm2的細胞密度接種于培養瓶中,于37 ℃、5%CO2條件下培養12 h,待細胞貼壁后,強力沖洗去除未貼壁細胞,然后消化貼壁細胞,收集并計數。
4.CD14+單核細胞分選:復蘇凍存的PBMC,于4 ℃、300×g離心10 min,收集細胞,加入分離緩沖液重懸細胞,并加入CD14 Microbeads(德國美天旎公司),4 ℃避光孵育15 min。將使用分離緩沖液預處理的分離柱置于分離架上,將上述細胞懸液加入分離柱中,CD14+細胞吸附于分離柱內,從分離架上取下分離柱,加入分離緩沖液,收集CD14+單核細胞并計數。
5.CD14+單核細胞與角質形成細胞共培養系統的建立:取純化的CD14+單核細胞,加入抗TIM-3抗體(美國R&D 公司,終濃度為10 mg/L)刺激培養12 h,對照組加入對照抗體IgG(美國R&D 公司,終濃度為10 mg/L)培養12 h,洗滌細胞后,取5×105個CD14+單核細胞,與同一患者分離的105個角質形成細胞直接混合共培養,向培養液中加入LPS 和R848刺激培養12 h,在刺激培養的后6 h加入布雷非德菌素A 抑制蛋白轉運,收集CD14+單核細胞進行TIM-3和IL-12的流式細胞檢測。
6.Western 印跡檢測細胞因子信號抑制物1(suppressor of cytokine signaling-1,SOCS-1)表達和抗信號傳導及轉錄激活因子1(signaling transducers and activators of transcription-1,STAT-1)磷酸化:取抗TIM-3刺激后的CD14+單核細胞,加入50 μl 細胞裂解液,冰上裂解15 min 后于95 ℃孵育10 min,收集上清液,加入等量的2×SDS-PAGE 緩沖液,取 30 μl 標本加入 SDS-PAGE 電泳濃縮膠孔中電泳至分離膠后,電轉移至PVDF膜。用封閉緩沖液封閉轉膜后的PVDF膜后,加入抗SOCS-1抗體(1∶1 000稀釋)、抗STAT-1抗體(1∶1 000稀釋)、抗磷酸化STAT-1抗體(1∶1 000稀釋)或抗GADPH抗體(1∶2 000稀釋),4 ℃孵育過夜。洗滌后加入抗兔IgG-HRP(1∶2 000 稀釋),室溫孵育2 h。所有抗體均購自美國Abcam 公司。洗滌后利用化學發光法顯影。使用ImageJ軟件對蛋白條帶進行灰度檢測。
7.統計學方法:所有數據均采用SPSS21.0軟件處理,先進行正態性檢測,符合正態分布,兩組間數據比較采用t檢驗,相關性分析采用Pearson 檢驗。P<0.05為差異有統計學意義。
結 果
一、PBMC中CD14+單核細胞表面TIM-3的表達
在無TLR 配體刺激狀態下,銀屑病組CD14+TIM-3+細胞比例(12.20% ± 2.83%)高于對照組(9.91% ± 1.77%),t= 3.27,P< 0.01。TLR 配體刺激后,與刺激前比較,CD14+TIM-3+細胞比例在銀屑病組(6.80% ± 1.11%)和對照組(4.85% ± 1.37%)均顯著降低(t=9.86 和12.17,均P< 0.01),但銀屑病組仍高于對照組(t=6.06,P< 0.01)。見圖1。
二、PBMC中CD14+單核細胞分泌IL-12的水平
在無TLR 配體刺激的狀態下,銀屑病組CD14+IL-12+細胞比例(0.80%±0.13%)和對照組(0.82%±0.19%)均較低,兩組比較,t= 0.453,P= 0.652。TLR 配體刺激后,與刺激前比較,CD14+IL-12+細胞比例在銀屑病組(13.49% ± 2.80%)和對照組(28.97% ± 8.97%)均增加(t=13.68 和t=30.99,均P< 0.01),但銀屑病組低于對照組(t= 10.71,P<0.01)。見圖2。
三、CD14+單核細胞TIM-3表達與IL-12分泌呈負相關
LPS+R848刺激后兩組CD14+單核細胞TIM-3表達與IL-12 的分泌均呈顯著負相關,對照組r=-0.47,P=0.039;銀屑病組r=-0.37,P=0.010。見圖3。
四、封閉TIM-3通路對銀屑病患者CD14+單核細胞分泌IL-12的影響
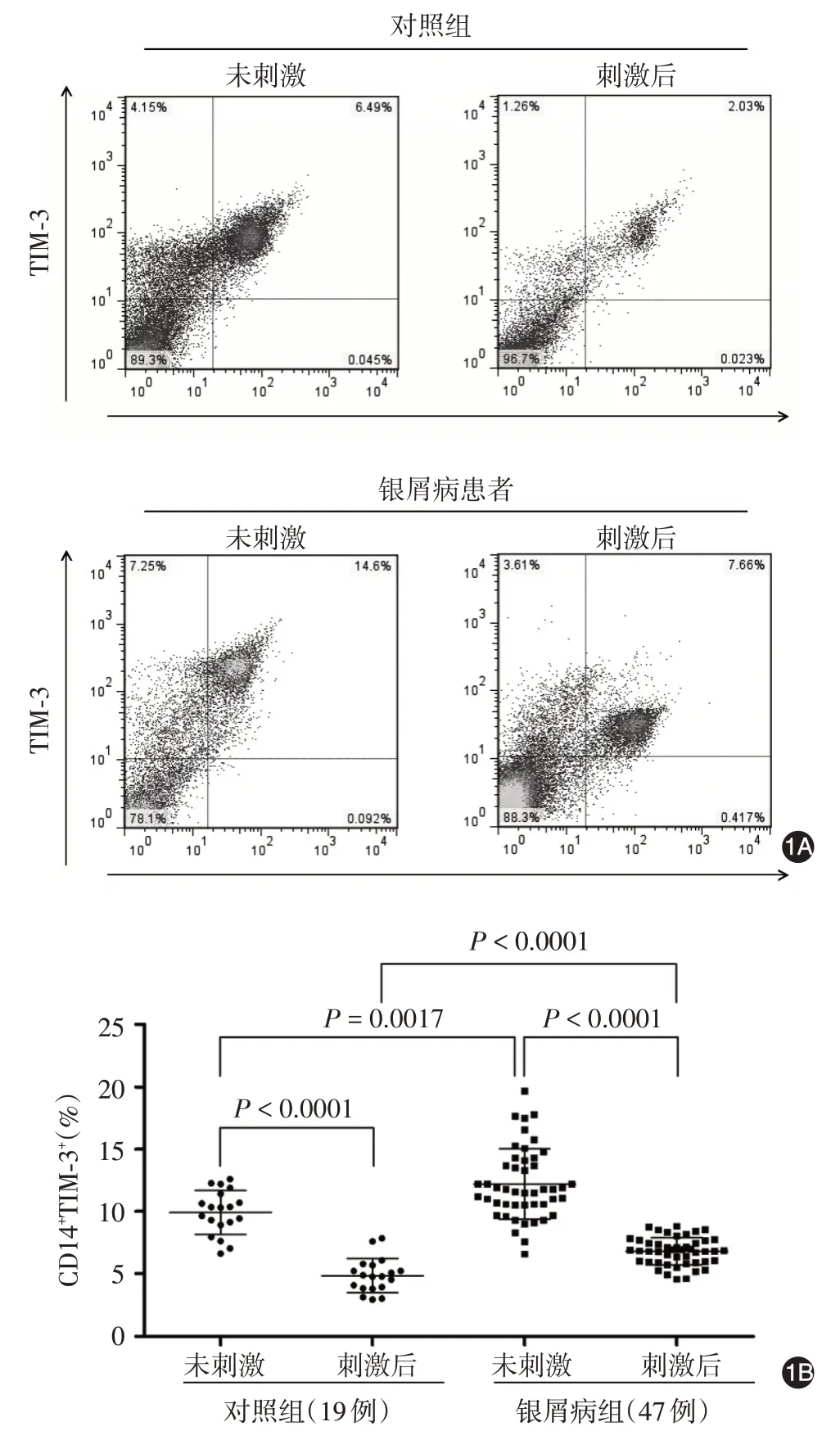
圖1 對照組和銀屑病組外周血單個核細胞中CD14+TIM-3+比例 1A:兩組刺激前后CD14+單核細胞表達TIM-3 的流式散點圖;1B:兩組CD14+TIM-3+細胞比例比較。TIM-3:T 細胞免疫球蛋白和黏蛋白結構域蛋白3

圖2 對照組和銀屑病組外周血單個核細胞中CD14+單核細胞分泌白細胞介素12(IL-12)水平比較 2A:兩組刺激前后CD14+單核細胞表達分泌IL-12 的流式散點圖;2B:兩組CD14+IL-12+細胞比例比較
使用抗TIM-3 抗體封閉TIM-3 通路可降低CD14+單核細胞 SOCS-1 的表達,IgG 對照組為0.54±0.12,抗TIM-3抗體組為0.08±0.02,t=7.14,P<0.01;STAT-1磷酸化水平則顯著升高,IgG對照組為0.34±0.08,抗TIM-3抗體組為0.89±0.10,t=8.96,P< 0.01。見圖4A。

圖3 對照組和銀屑病組CD14+單核細胞T細胞免疫球蛋白和黏蛋白結構域蛋白3(TIM-3)表達與白細胞介素12(IL-12)分泌的相關性分析 3A:對照組;3B:銀屑病組

圖4 抑制TIM-3對10例銀屑病患者CD14+單核細胞下游信號分子和IL-12分泌的影響 4A:SOCS-1、磷酸化STAT-1、總STAT-1的典型Western印跡檢測;4B:CD14+單核細胞單獨培養及與角質形成細胞共培養系統中的IL-12+細胞比例。TIM-3:T細胞免疫球蛋白和黏蛋白結構域蛋白3;IL-12:白細胞介素12;SOCS-1:細胞因子信號抑制物1;STAT-1:信號傳導及轉錄激活因子1
取10 例銀屑病患者CD14+單核細胞,加入抗TIM-3 抗體或對照抗體IgG 培養,單獨培養或建立與角質形成細胞的共培養系統。在CD14+單核細胞單獨培養系統中,CD14+IL-12+細胞比例在抗TIM-3 抗體處理組(18.63% ± 4.36%)和IgG 處理組(15.33% ± 5.91%)間差異無統計學意義(t= 1.42,P=0.17,圖4B);在CD14+單核細胞與自體角質形成細胞的共培養系統中,抗TIM-3 處理組CD14+IL-12+細胞比例(35.02%±10.91%)則顯著高于IgG處理組(23.03% ± 8.55%),t= 2.73,P= 0.014,圖4B。共培養系統中,抗TIM-3 抗體組與對照組CD14+IL-12+細胞比例均顯著高于單獨培養系統組(t=4.41和2.34,P=0.0003和0.031,圖4B)。
討 論
TIM-3 作為一種T 細胞表面抑制性因子,主要抑制CD4+和CD8+T 細胞,降低促炎細胞因子的分泌,發揮免疫抑制功能[8]。在銀屑病患者中,活化Th1細胞和Th17細胞表面TIM-3表達顯著降低,后者使Th1 細胞和Th17 細胞逃避了由TIM-3 介導的負性免疫調控,促進銀屑病的發生發展[9]。而新近的研究亦發現,TIM-3還可調控單核巨噬細胞的功能活性。在胃癌[10]、自發性腦出血[11]、瘧疾[12]等疾病過程中,CD14+單核細胞中TIM-3 的表達均顯著升高,且與疾病進程密切相關。在慢性乙型肝炎患者中,單核細胞的TIM-3高表達可增加促炎細胞因子的分泌和Th17 細胞的應答,促進機體炎癥應答[13]。而在慢性丙型肝炎患者中,高表達TIM-3的單核細胞可通過抑制IL-12的分泌發揮負性調控免疫應答的功能[5,14]。
本研究發現,銀屑病患者CD14+單核細胞表面TIM-3的表達顯著高于對照組。但是,應用TLR配體活化單核細胞后,無論在對照組還是銀屑病組,CD14+單核細胞表面TIM-3 的表達均降低,但銀屑病組中CD14+TIM-3+的比例仍高于對照組,這與在慢性丙型肝炎中的發現相一致[14]。更為重要的是,TLR配體可誘導CD14+單核細胞分泌IL-12,而銀屑病組單核細胞分泌IL-12 的水平較對照組顯著降低,且與TIM-3的表達呈負相關。這提示在銀屑病中,無論在靜息還是活化狀態下,CD14+單核細胞中高表達的TIM-3都可能發揮負性免疫調控作用,通過降低IL-12 的分泌發揮抑制單核細胞功能的作用。
本研究發現,銀屑病角質形成細胞可誘導CD14+單核細胞表達IL-12,而封閉TIM-3通路可促進角質形成細胞誘導的CD14+單核細胞分泌IL-12水平進一步升高,但其中的分子機制尚未闡明。TIM-3 通路在不同疾病狀態下存在不同甚至完全相反的調控效應。在慢性丙型肝炎患者中,TIM-3可與CD14+單核細胞中的SOCS-1相互作用,通過抑制STAT-1 的磷酸化發揮負性免疫調控作用,促進感染慢性化[14]。相反,在腫瘤小鼠模型中,TIM-3可通過抑制STAT-1/miR-155 軸,上調SOCS-1 的表達,增強單核巨噬細胞M2 方向的極化[15]。本研究發現,使用抗TIM-3 抗體封閉銀屑病患者CD14+單核細胞中的TIM-3 通路促進STAT-1 的磷酸化,而SOCS-1 表達下降,這與在慢性丙型肝炎中的研究結果[14]相一致。基于倫理原因,本研究未納入正常人的角質形成細胞,因此,TIM-3 對正常人角質形成細胞介導的細胞因子分泌的影響尚不清楚。但本研究結果仍提示TIM-3可直接調控CD14+單核細胞的功能,參與機體固有免疫應答。
總之,在銀屑病患者皮損中,角質形成細胞可誘導銀屑病CD14+單核細胞TIM-3 表達升高,從而負性調控CD14+單核細胞介導的固有免疫應答。
利益沖突所有作者均聲明不存在利益沖突
