玉米根際土壤中大環(huán)內(nèi)酯類抗性基因的分布特征
2019-10-08 08:10:46陳運杰郭欣妍
生態(tài)與農(nóng)村環(huán)境學報 2019年9期
陳運杰,郭欣妍,楊 藝,王 娜①,楊 燁
(1.南京中醫(yī)藥大學醫(yī)學與生命科學學院,江蘇 南京 210023;2.生態(tài)環(huán)境部南京環(huán)境科學研究所,江蘇 南京 210042)
抗生素作為畜禽感染性疾病治療藥物及動物生長促進添加劑,在養(yǎng)殖業(yè)中發(fā)揮著非常重要的作用[1]。然而,長期大量依賴抗生素的養(yǎng)殖方式,可誘導動物腸道菌群產(chǎn)生大量攜帶抗生素抗性基因(antibiotic resistance genes,ARGs)的耐藥菌(antibiotics resistance bacteria,ARBs),且耐藥菌會隨排泄物最終進入環(huán)境[2-3];同時,抗生素藥物母體和代謝產(chǎn)物隨著糞便等排泄物直接進入土壤、水等環(huán)境介質(zhì)中,又增加了環(huán)境微生物的進化選擇壓力,將誘導微生物產(chǎn)生耐藥性[4]。
最近一些研究認為,植物根際是耐藥基因轉(zhuǎn)移的“熱區(qū)”[5]。植物根際指受植物根系的影響,在物理、化學和生物學特性上不同于周圍土體的根表面微域土區(qū),是土壤水分、養(yǎng)分和有益或有害微生物進入植物根系的門戶。在根際范圍內(nèi),微生物種類豐富,能量和物質(zhì)代謝活躍,根際生長和營養(yǎng)成分滲出會影響細胞密度、分布以及土壤細菌的代謝能力,一些抗生素的抗性效應(yīng)在此區(qū)域內(nèi)會進一步得到加強。REICHEL等[6]研究了施用含磺胺嘧啶(SDZ)和二氟沙星(DIF)的糞肥后玉米根際土壤與非根際土壤的微生物結(jié)構(gòu)多樣性,對63 d中宇宙試驗的土壤樣品磷脂脂肪酸(PLFA)進行分析發(fā)現(xiàn),施用含SDZ和DIF的糞肥增加了土壤中革蘭陰性菌和真菌數(shù)量,但是根際土壤微生物菌群的變化與非根際土壤相比并不十分明顯;通過分析PCR-DGGE條帶發(fā)現(xiàn),條件致病菌假單胞菌屬(Pseudomonas)在根際土壤中的增加更為顯著。這表明植物根際環(huán)境可能促進土壤微生物群落產(chǎn)生耐藥性,也可能促使糞肥中抗性基因在微生物間水平轉(zhuǎn)移。因此,植物根際可能是影響ARGs增殖與傳播行為的重要環(huán)境。但迄今為止,相關(guān)文獻報道較少,研究角度比較單一,且所研究的抗生素品種主要集中于磺胺類抗生素,僅局限于施用含抗生素的糞肥后植物根際微生物多樣性變化[6],以及植物根際部位磺胺類抗生素及其抗性基因(sul1、sul2和sul3)的表觀分布規(guī)律[7-8]。
世界衛(wèi)生組織(World Health Organization,WHO)報告指出,人畜共用抗生素的使用會大大增加動物與人體內(nèi)抗性細菌交叉感染的風險,對人體健康造成極大危害[9]。大環(huán)內(nèi)酯類抗生素(包括紅霉素、螺旋霉素、乙酰螺旋霉素、交沙霉素和羅紅霉素等)不僅在臨床上作為青霉素過敏患者的替代藥物而發(fā)揮著重要作用,對于養(yǎng)殖業(yè)也是廣泛使用的一類獸藥品種。目前,關(guān)于養(yǎng)殖業(yè)抗性基因的報道以四環(huán)素和磺胺類為主,但有關(guān)養(yǎng)殖行業(yè)使用頻率同樣較高的大環(huán)內(nèi)酯類抗生素所導致的ARGs(主要為erm基因)傳播擴散問題對人體健康的隱患更為嚴重。這是由于大環(huán)內(nèi)酯類ARGs是一類較為穩(wěn)定的抗性基因,在土壤中殘留豐度相對更高。JOY等[10]在豬糞中檢出金霉素、泰樂菌素及其對應(yīng)的抗性基因tet和erm,在40 d的堆肥過程中,雖然泰樂菌素降解90%,但其對應(yīng)的ermB基因卻只降解40%。KNAPP等[11]對荷蘭和蘇格蘭不同地區(qū)農(nóng)田土壤中抗性基因進行定量分析,結(jié)果顯示大環(huán)內(nèi)酯類ARGs(ermB、ermC、ermE和ermF)豐度隨著時間的推移而呈遞增趨勢。SU等[12]研究發(fā)現(xiàn),廣東東江流域水底沉積物中ermB、ermC和ermF這3種大環(huán)內(nèi)酯類ARGs也存在較高豐度(105~1011copies·g-1)。可見大環(huán)內(nèi)酯類抗性基因的環(huán)境潛在風險不容忽視。
筆者以污染隱患較大的大環(huán)內(nèi)酯類抗性基因(erms)為研究對象,采用陸生微宇宙模擬試驗方法,在人工控制條件下模擬目標污染物進入土壤中的分布行為。利用實時熒光定量PCR(real-time fluorescence quantitative PCR,qPCR)技術(shù)探明植物根際對erms在土壤中演變與歸趨的影響規(guī)律,以及其在土壤剖面的垂直分布規(guī)律,為闡明erms在農(nóng)田環(huán)境中傳播擴散機制提供科學依據(jù),也為建立抗性基因的生態(tài)環(huán)境安全評價和預警體系提供理論基礎(chǔ)。
1 材料與方法
1.1 微宇宙試驗方法的建立
1.1.1糞肥的收集
供試動物糞肥采集于江蘇省農(nóng)業(yè)科學院六合動物科學基地。選取1頭育肥豬,首先在不飼喂任何抗生素藥物情況下培育2個月,之后添加抗生素藥物飼喂,方法為每天將1 g硫氰酸紅霉素溶于8 L水中,令其自由飲用,連續(xù)飲用4 d,采集7 d內(nèi)產(chǎn)生的豬糞便。糞便經(jīng)短期堆肥腐化后收集并于-20 ℃條件下冷凍保存?zhèn)溆谩?/p>
1.1.2土柱單元設(shè)置
陸生微宇宙系統(tǒng)的土芯取自生態(tài)環(huán)境部南京環(huán)境科學研究所實驗用地,土壤為水稻土,其基本理化性質(zhì):pH為6.23,w(有機碳)為49.8 g·kg-1,離子交換量為18.0 cmol·kg-1,w(黏粒)為19.4%,w(粉粒)為75.8%,w(砂粒)為4.8%。土柱桶材質(zhì)為聚乙烯,內(nèi)徑為17 cm,高度為60 cm(包括0.15 m厚種植層土和0.45 m厚深層土)。填裝土柱前,用電子混勻機將糞肥和水稻土(以干重計)按照1∶25質(zhì)量比充分混合,用水稻土填裝深土層(0.45~0.6 m)后,再用糞肥-土壤混合物填裝種植層(0~0.15 m)。土柱填裝后先對土柱進行預淋溶,使土柱持水,之后按正常種植方式種植玉米并正常管理,每日適量補水保持土柱質(zhì)量含水率為8.5%~15.2%。微宇宙系統(tǒng)的溫度恒定設(shè)為21 ℃,采用鹵鎢燈(400 W)控制光照周期為t(光)∶t(暗)=16 h∶8 h[13]。以種植玉米的土柱作為試驗組,以不種植玉米的土柱作為對照組(CK),其余處理方式與種植玉米的試驗組完全相同。
1.2 樣本采集
玉米種子購于南京潤祥種業(yè)有限公司(品名:江南花糯,由江蘇省農(nóng)業(yè)科學院糧食作物研究所研制),每柱播種3粒種子,待預生長1周后,每柱僅保留1株生長良好的幼苗作為試驗用玉米株,并以此作為玉米生長的第0天(D0)。之后分別于生長7、14、28、42、56和63 d(D7、D14、D28、D42、D56和D63)時采集土樣,主要分為2個部分:(1)對試驗組土柱中根際(rhizosphere,RH)和非根際(bulk soil,BK)土樣進行采集。參照文獻[7],采用抖動植物根際方式采集貼近根系0~4 mm的根際土壤約5 g,而非根際土壤則使用圓形取樣器(高為9.2 cm,內(nèi)徑為1.2 cm)采集同一土柱中表層(0~0.1 m)土壤獲得。(2)在每個采樣時段利用特制的推土裝置將試驗組和對照組(CK)土芯整體推出后,同時取0~0.2、>0.2~0.4和>0.4~0.6 m這3段各自混為1個樣品。所有樣品均采集3個平行樣,混合后置于無菌保鮮袋中,于4 ℃條件下無菌保存。
1.3 土壤總DNA(gDNA)的提取
土壤gDNA提取參照PowerSoil?DNA Isolation Kit試劑盒說明書操作,DNA洗脫液于-80 ℃條件下保存待用。采用w=1.0%的瓊脂糖凝膠電泳和Nandrop1000對提取的gDNA質(zhì)量和濃度進行檢測。
1.4 實時熒光定量PCR(qPCR)
1.4.1PCR擴增抗性基因特異性片段
所涉及的目標基因主要包括4種大環(huán)內(nèi)酯類抗性基因(ermB、ermC、ermF和ermX)和16S rDNA(作為分析時的內(nèi)參基因),引物設(shè)計參照文獻[14],具體信息見表1。以土壤gDNA為模板,PCR反應(yīng)過程:94 ℃預變性300 s,然后進行30個循環(huán),每個循環(huán)包括94 ℃變性30 s,適當退火溫度(Tm)下退火30 s,72 ℃延伸45 s,最后72 ℃終延伸360 s結(jié)束反應(yīng),反應(yīng)體系為12.5 μL 的2×ES Taq MasterMix,1 μL 上游引物(10 μmol),1 μL下游引物(10 μmol),1 μL模板DNA(<500 ng),9.5 μL的RNase-free水,反應(yīng)體系總體積為25 μL。
表1 qPCR引物序列
Table 1 Primer sequence for qPCR

基因名稱引物名稱退火溫度/℃產(chǎn)物大小/bpermBFW: ACCTTGGATATTCACCGAAC58119RV: CTTTGGCGTGTTTCATTGCTermCFW: ACTGCCATTGAAATAGACCA56292RV: TAGCAAACCCGTATTCCACermFFW: AAGAAATTGCCCGTTCGTT55274RV: ACTTGAACATTTCGGGCATCermXFW: AAATTTTCTCACCGACCACA62219RV: AGTACAGCGTCAGTCCA16SrDNAFW: CAATGGACGAAAGTCTGACG60146RV: ACGTAGTTAGCCGTGGCTTT
1.4.2目的基因的純化與擴增
將含目的基因片段的PCR產(chǎn)物進行w=1.5%的瓊脂糖凝膠電泳后,利用PCR純化試劑盒(EasyPure Qucik Gel Extraction Kit)對目的基因條帶進行純化回收。將目的基因轉(zhuǎn)染至Trans-T3感受態(tài)細胞進行克隆培養(yǎng)。最后,利用Axy Prep Plasmid Miniprep Kit提取質(zhì)粒DNA,再由蘇州金唯智生物科技有限公司測序以檢測目的基因片段是否插入。
1.4.3基因拷貝數(shù)計算
利用Nanorop1000對所提取質(zhì)粒純度和DNA含量進行檢測,若A260/A280介于1.8~2.0之間且A260/A230>2.0,則認為是純DNA,否則需進一步純化。參照文獻[15],將質(zhì)粒濃度換算為1 μL質(zhì)粒溶液所攜帶的絕對模板拷貝數(shù)(y,copies·μL-1)公式為
y=[x/(a+b)×660]× 10-9× 6.02 × 1023。
(1)
式(1)中,x為質(zhì)粒濃度,ng·μL-1;a為載體長度,bp;b為目的基因長度,bp。
1.4.4標準曲線的建立
將已知初始拷貝數(shù)的標準品質(zhì)粒依次進行10倍梯度稀釋,稀釋濃度分別為10-2、10-3、10-4、10-5、10-6、10-7、10-8和10-9,再進行qPCR檢測,反應(yīng)結(jié)果經(jīng)熒光定量軟件分析得到對應(yīng)循環(huán)數(shù)(Ct,Y),以其作為縱坐標,以拷貝數(shù)的lg值(X)作為橫坐標,繪制標準曲線。4種大環(huán)內(nèi)酯類抗性基因及16S rDNA標準曲線見表2。
在qPCR中,隨著循環(huán)數(shù)的增加,熒光強度逐漸增強,在經(jīng)過一段時間的指數(shù)擴增后,擴增曲線趨于穩(wěn)定,出現(xiàn)“平臺效應(yīng)”,即隨著模板擴增循環(huán)數(shù)的增加,由于其熒光強度和循環(huán)數(shù)不同,從而得到不同循環(huán)數(shù)與熒光強度之間的關(guān)系圖,即擴增曲線。以ermB為例,其擴增曲線、標準曲線、溶解曲線和熔融峰見圖1(a)~(d)。
表2 大環(huán)內(nèi)酯類抗性基因及16S rDNA標準曲線
Table 2 Standard curve equation of macrolides antibiotic resistance genes and 16S rDNA
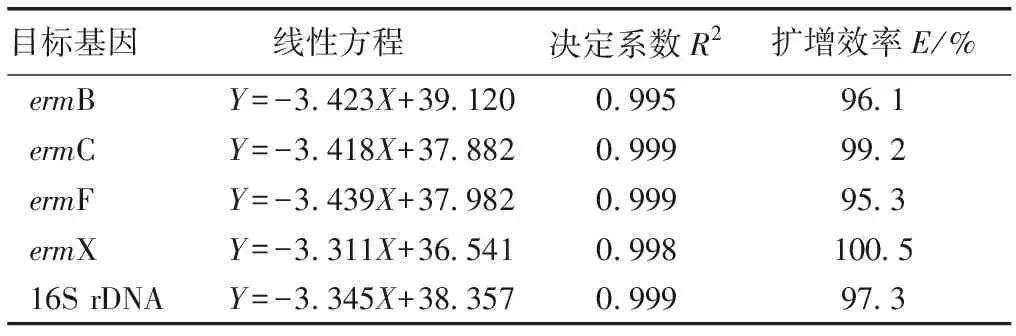
目標基因 線性方程 決定系數(shù)R2擴增效率E/%ermBY=-3.423X+39.1200.99596.1ermCY=-3.418X+37.8820.99999.2ermFY=-3.439X+37.9820.99995.3ermXY=-3.311X+36.5410.998100.516S rDNAY=-3.345X+38.3570.99997.3
X為目標基因拷貝數(shù)的lg值,Y為循環(huán)數(shù)。
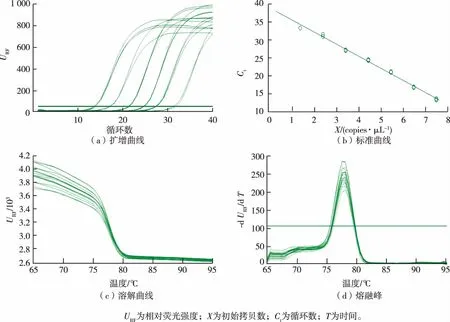
圖1 ermB基因的擴增曲線、標準曲線、溶解曲線和熔融峰
1.5 抗性基因(ARGs)的定量分析
以16S rDNA為內(nèi)參基因,使用qPCR技術(shù)對ermB、ermC、ermF和ermX 4種大環(huán)內(nèi)酯類抗性基因進行定量分析,反應(yīng)體系為5 μL 的2×UltraSYBR Mixture,0.2 μL上游引物(10 μmol),0.2 μL下游引物(10 μmol),1 μL模板DNA(10~100 ng),3.6 μL 的RNase-free 水,反應(yīng)體系總體積為10 μL。
qPCR反應(yīng)于CFX96 TouchTMReal-Time PCR Detextion System(Bio-Rad,USA)儀器上進行,采用三步法進行反應(yīng):95 ℃預變性10 min,然后進行40個循環(huán),每個循環(huán)包括95 ℃變性15 s,適當退火溫度(表1)下退火30 s,72 ℃延伸30 s。溶解曲線程序按照儀器默認值設(shè)置,從60 ℃上升到95 ℃,其間每隔0.5 ℃采集1次熒光以生成溶解曲線,根據(jù)溶解曲線變化檢測擴增結(jié)果的特異性。
1.6 數(shù)據(jù)統(tǒng)計
所有試驗數(shù)據(jù)均重復測定3次,以平均值±標準差形式表示,采用SPSS 22.0軟件進行數(shù)據(jù)處理與分析,以P<0.05表示組間差異顯著。大環(huán)內(nèi)酯類抗性基因(erms)相對豐度為erms濃度與16S rDNA濃度的比值,即erms∶16S rDNA的計算值。采用OriginLab 8.0軟件制圖。
2 結(jié)果與分析
2.1 根際(RH)與非根際(BK)土壤中大環(huán)內(nèi)酯類抗性基因的分布
根據(jù)標準曲線計算得到RH和BK土壤中大環(huán)內(nèi)酯類抗性基因(erms)相對豐度。如圖2所示,4種大環(huán)內(nèi)酯類抗性基因均可檢出,相對豐度范圍大致為10-4~10-1。在檢測的2種土壤中,ermF和ermX相對豐度較高,為10-3~10-1;其次為ermB,為10-3~10-2;而ermC相對豐度最低,為10-4~10-3。如表3所示,BK土壤中ermC、ermF和ermX在7 d時的相對豐度均顯著高于RH土壤(P<0.05),而63 d時,除ermF基因外,RH和BK土壤中抗性基因相對豐度無顯著差異(P>0.05)。
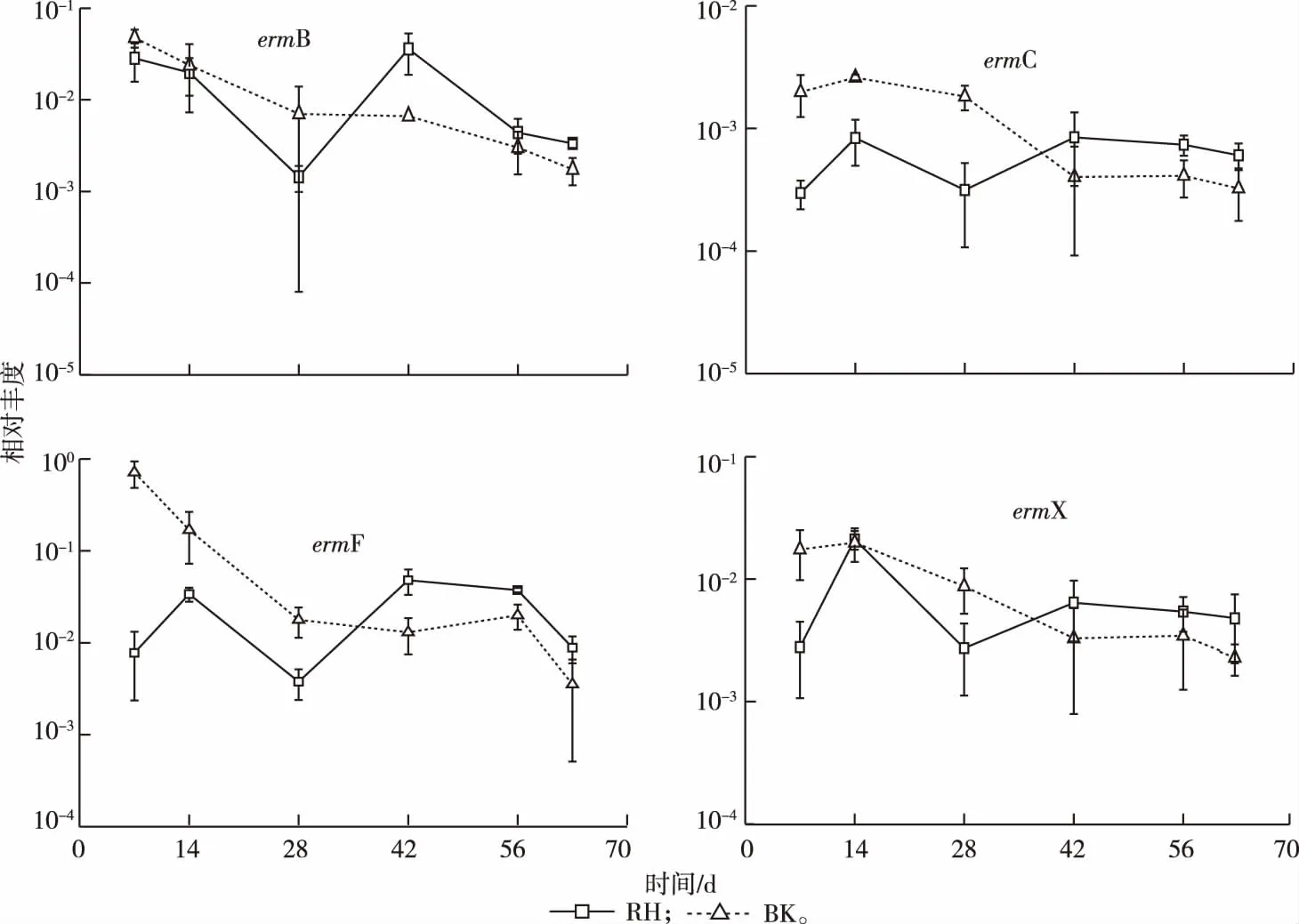
圖2 根際(RH)和非根際(BK)土壤中大環(huán)內(nèi)酯類抗性基因相對豐度
圖2顯示,在63 d的試驗周期內(nèi),隨著玉米生長時間的推移,BK土壤中erms相對豐度總體呈下降趨勢,而RH土壤中4種抗性基因相對豐度變化均呈現(xiàn)波動狀態(tài),部分抗性基因在63 d時出現(xiàn)增殖。具體表現(xiàn)為7 d時,BK土壤中ermB、ermC、ermF和ermX相對豐度分別為4.72×10-2、1.98×10-3、7.13×10-1和1.75×10-2,而63 d時則分別為1.74×10-3、3.24×10-4、3.53×10-3和2.28×10-3,其增幅分別為-96.3%、-83.6%、-99.5%和-87.0%。類似地,RH土壤中4種抗性基因相對豐度增幅則分別為-88.3%、103.0%、-88.6%和71.5%。
28 d之前,土壤中抗性基因相對豐度表現(xiàn)為BK > RH,但28 d時,RH土壤中erms相對豐度開始呈現(xiàn)上升趨勢,并在42 d后表現(xiàn)為RH > BK,且除ermB基因(42 d時)和ermF基因(42、63 d)外,其余差異性并不顯著(表3)。
2.2 剖面土壤中大環(huán)內(nèi)酯類抗性基因垂直分布
種植玉米(MA)和未種植玉米(CK)剖面土壤中抗性基因相對豐度見圖3。在MA和CK土壤中,4種erms均能被檢測到,且相對豐度隨著玉米生長周期的增加呈下降趨勢。與RH和BK土壤中抗性基因相對豐度的檢測結(jié)果相似,MA和CK剖面土壤中ermF和ermX相對豐度較高,ermB和ermC相對豐度較低。MA土壤中ermB、ermC、ermF和ermX相對豐度分別為10-6~10-2、10-6~10-3、10-4~10-2和10-4~10-3,較CK土壤的10-7~10-3、10-6~10-4、10-5~10-2和10-5~10-3高1~2個數(shù)量級。總體而言,MA各土壤層中erms相對豐度高于CK土壤。
表3 根際和非根際土壤中抗性基因顯著性差異P值分析
Table 3 Variance analysis of ARGs relative abundance in rhizosphere and bulk soil

抗性基因不同時間(d)P值71428425663ermB0.0670.9830.1300.0190.1280.368ermC0.0350.0730.0300.1580.0780.137ermF0.0280.0220.2860.0220.1920.027ermX0.0470.4610.3950.0820.0620.051
就0~0.2 m土層而言,無論是MA還是CK土壤,4種erms均可檢出,其中ermF相對豐度最高,在10-1~10-2之間,ermC相對豐度最低,在10-4~10-3之間。就>0.2~0.4 m土層而言,MA土壤中除了ermB直到42 d時才被檢測到以外,其余3種抗性基因在不同采樣時間均可被檢測到,且相對豐度呈現(xiàn)先增長后下降趨勢;而就CK土壤而言,除了ermB在56 d之后才被檢測到以外,其余3種抗性基因在不同采樣時間均可被檢測到;在同一采樣時間,MA土壤中erms相對豐度高于CK土壤。就>0.4~0.6 m土層而言,28 d之前,MA土壤中4種erms均未檢出,其中ermC和ermF在28 d之后才檢出,ermB 在42 d之后才檢出,ermX則在56 d之后檢出;而對于CK土壤,除了ermC較早被檢測到(28 d時)以外,ermB和ermF直到56和42 d之后才檢出,ermX始終未檢出。
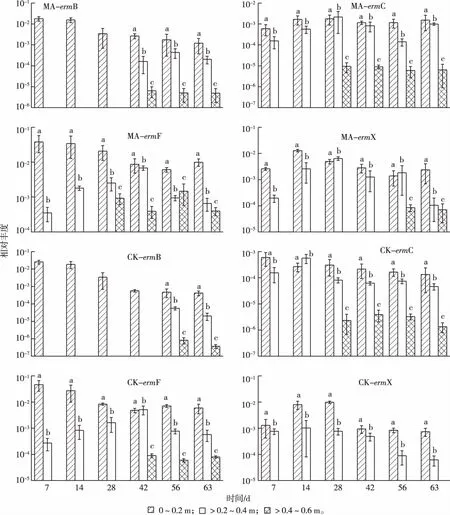
MA和CK分別指種植玉米和未種植玉米土壤不同深度土層中抗性基因相對豐度。就同一幅圖而言,同一組直方柱上方
通過對剖面土壤中抗性基因垂直分布的研究發(fā)現(xiàn),不同抗性基因在土層中的分布也有顯著差異(圖3),例如,MA土壤0~0.2 m土層中ermB基因(42 d時)相對豐度比>0.4~0.6 m土層高1 000倍,而ermC相對豐度則高100倍。隨著剖面土壤深度的增加,抗性基因相對豐度逐漸下降,各土層抗性基因相對豐度由大到小依次為0~0.2、>0.2~0.4和>0.4~0.6 m。
3 討論
筆者研究中,在63 d的采樣周期內(nèi),非根際(BK)土壤中ermC、ermF和ermX相對豐度在7 d時均顯著高于RH土壤(P< 0.05),這可能與非根際土壤主要取自含有糞肥的耕作層(0~0.1 m)有關(guān)。POPOWSKA等[16]從動物糞便堆肥樣本和施用糞肥的農(nóng)田及有機果園土壤樣本中分離得到紅霉素耐藥菌,采用定性PCR技術(shù)檢測到耐藥菌攜帶有ermC、ermV、ermX、msrA、oleB和vg基因,可見,大環(huán)內(nèi)酯類抗性基因通過糞便施肥會廣泛進入環(huán)境并進一步傳播擴散。隨著時間的推移,BK土壤中erms相對豐度總體呈下降趨勢,63 d時抗性基因相對豐度比7 d時增加-99.5%~-83.6%,這可能是因為在一段較長時間后,由于土壤中抗生素逐漸降解,在缺少外源壓力的情況下,相應(yīng)的抗性基因表現(xiàn)為相對穩(wěn)定或持續(xù)性緩慢下降[17]。而在RH土壤中抗性基因相對豐度增幅則為-88.6%~103.0%,erms相對豐度總體變化趨勢較BK更穩(wěn)定,63 d時4種抗性基因相對豐度均超過BK土壤,暗示根際對erms豐度的維持具有一定作用,使得根際周圍的土壤更有可能成為抗性基因的“蓄積池”。
在28 d之前,BK土壤中erms相對豐度均高于RH土壤,但28 d之后RH開始呈現(xiàn)上升趨勢,在42 d之后出現(xiàn)逆轉(zhuǎn),表現(xiàn)為RH土壤中erms相對豐度高于BK土壤。RH土壤中erms的這種逆轉(zhuǎn)情況可能與玉米生長過程中根際分泌物不同,進而引起根際周圍攜帶有抗性基因的微生物群落變化有關(guān)[18]。根據(jù)玉米的生長周期,28 d之后,玉米正處于生長的關(guān)鍵期——穗期,此時根系繼續(xù)擴展,受其分泌物的持續(xù)性影響,根際周圍土壤養(yǎng)分更為豐富,微生物多樣性和豐富度顯著高于非根際[19]。研究顯示,erm基因容易被質(zhì)粒或轉(zhuǎn)座子等水平基因移動原件(HGT)捕獲,可以非常容易地在不同宿主細菌之間轉(zhuǎn)移[12]。BRANDT等[20]研究發(fā)現(xiàn),在添加人工根際滲出液作為碳源后,土壤中磺胺嘧啶(SDZ)的細菌群落抗性和相應(yīng)的磺胺類抗性基因大大增加,這可能是因為 SDZ 耐藥菌得到增殖。JECHALKE等[21]研究了植物根際對SDZ抗性轉(zhuǎn)移的影響作用,發(fā)現(xiàn)玉米和青草的根際區(qū)域抗性質(zhì)粒轉(zhuǎn)移率明顯增加,同時結(jié)果顯示植物的不同品種及不同發(fā)育周期會對土壤微生物活性的促進作用以及抗生素抗性基因的轉(zhuǎn)移作用產(chǎn)生影響。
通過對剖面土壤中抗性基因垂直分布的研究發(fā)現(xiàn),4種erms在不同深度土層中的分布不一,且種植玉米(MA)土壤中抗性基因相對豐度顯著高于未種植玉米(CK)土壤。隨著縱向距離的加深,抗性基因檢出頻率和豐度也相應(yīng)呈遞減趨勢。JOY等[10]研究發(fā)現(xiàn),糞便耐藥菌攜帶的sul1、sul2和ermF 基因容易隨著雨水而滲透土層,徑流至周邊的地表水或逐漸滲透至地下水,說明抗生素抗性基因具有能夠沿著土壤界面向下遷移的能力,繼而可能會對地下水造成污染,具有潛在的環(huán)境風險。
隨著土壤深度的增加,土壤養(yǎng)分和含氧量逐漸下降,從而使得土壤中微生物多樣性呈現(xiàn)差異性分層,進而導致攜帶抗性基因微生物豐度隨著土壤深度增加而減少[22]。RYSZ等[23]研究發(fā)現(xiàn),大腸埃希菌(Escherichiacoli)在厭氧環(huán)境下易丟失抗性基因,含氧量高低將會影響微生物對抗性基因的保持能力。因此,土壤含氧量變化會對抗性基因豐度產(chǎn)生影響。此外,抗性基因在土壤中的傳播還與可移動基因元件存在密切關(guān)系[12],因而呈現(xiàn)不同抗性基因在土壤垂直分層中的分布存在顯著差異(P< 0.05)。MA土壤中4種erms相對豐度范圍為 10-6~10-2,較CK土壤(10-7~10-2)高1~2個數(shù)量級,而在同一采樣時間,CK深層(>0.2~0.4和>0.4~0.6 m)土壤中抗性基因的檢出頻率和豐度顯著低于MA。研究結(jié)果提示植物根際對土壤中抗性基因的增殖及向下遷移具有一定作用。
4 結(jié)論
在玉米根際和非根際土壤中,4種大環(huán)內(nèi)酯類抗性基因(erms)均被檢出,且ermF相對豐度最高,ermC相對豐度最低。隨著玉米生長時間的推移(7~63 d),BK土壤中erms相對豐度呈逐漸下降趨勢,而RH土壤中erms相對豐度雖呈現(xiàn)一定波動,但最終趨于穩(wěn)定,并高于BK土壤,提示根際對于土壤中抗性基因豐度的促進和維持具有明顯作用。
通過對剖面土壤中抗性基因垂直分布的研究發(fā)現(xiàn),不同抗性基因在土壤中的縱向分布差異顯著,不同土層中4種erms相對豐度由大到小依次為0~0.2、>0.2~0.4和>0.4~0.6 m。種植玉米(MA)土壤中抗性基因相對豐度顯著高于未種植玉米(CK)土壤,隨著縱向距離的加深,抗性基因的檢出頻率和豐度也相應(yīng)呈遞減趨勢。這表明植物根際的存在促進了土壤中抗性基因豐度的增加以及在剖面土壤中的縱向遷移。
總之,植物根際的存在對抗性基因在土壤中的增殖、擴散及縱向遷移具有一定推動作用,這種作用可能主要與根際分泌物對土壤中細菌群落的影響有關(guān),因此進一步分析玉米生長過程中根際與非根際土壤中細菌群落的組成變化,將為揭示植物根際對土壤中抗性基因的影響提供一定理論基礎(chǔ)。
