氮、磷、硫對鉻超富集植物李氏禾吸收CrⅥ的影響
2019-09-23 06:10:53伍嬋翠劉杰梁英
江蘇農業科學 2019年14期
伍嬋翠 劉杰 梁英


摘要:通過水培試驗,以用25% Hoaglands營養液培養的植株作為對照,研究無機營養元素氮、磷、硫對鉻超富集植物李氏禾吸收六價鉻[Cr(Ⅵ)]的影響。結果表明,李氏禾根系對Cr(Ⅵ)的吸收符合系Michaelis-Menten方程,其米氏常數(Km)為91.84 μmol/L。與全素培養條件相比,在缺氮、缺硫條件下,李氏禾的根系吸收位點與Cr(Ⅵ)的親和力增加,根系分泌物能增加土壤中難溶性鉻的活性,從而有利于對Cr(Ⅵ)的吸收;而在缺磷培養條件下,根系分泌物不利于土壤中難溶性鉻的溶出,缺磷條件對李氏禾吸收Cr(Ⅵ)存在非競爭性抑制作用。由研究結果可知,適當施用磷肥有利于提高李氏禾對Cr(Ⅵ)污染土壤的植物修復效率。
關鍵詞:超富集植物;李氏禾;氮;磷;硫;植物修復效率
中圖分類號: X173 ?文獻標志碼: A ?文章編號:1002-1302(2019)14-0316-05
受人類活動的影響,土壤中的六價鉻[Cr(Ⅵ)]污染已經成為亟待解決的環境問題之一。李氏禾(Leersia hexandra Swartz)是我國境內發現并報道的首例鉻超富集植物[1],因其強大的吸收和富集Cr(Ⅵ)的能力,在Cr(Ⅵ)污染土壤的修復方面具有良好的應用前景。然而,重金屬污染土壤的植物修復效率受到很多外界因素的影響,土壤中的非金屬營養元素正是影響因素之一。此外,氮(N)、磷(P)、硫(S)對土壤重金屬形態轉化及植物吸收累積的影響也成為環境領域的研究熱點之一。
已有一些研究者分析了不同營養元素對植物吸收重金屬的影響。樓玉蘭的研究表明,硝態氮的施用會促進OH-或HCO3-的釋放,使根際pH值上升,從而降低土壤中重金屬的活性,減少了植物對重金屬的吸收[2]。郭光光等的研究表明,施用磷肥提高了蓖麻地下部對銅(Cu)的吸收和積累[3]。López-Bucio等研究發現,磷抑制了擬南芥對鉻的吸收[4]。Shi等研究發現,在銅污染土壤中施加硫肥有利于水稻對銅的吸收累積[5]。Zhong等研究發現,在砷(As)污染土壤中施用硫肥可減少油菜籽粒中的砷含量[6]。Marchiol等利用高粱(Sorghum bicolor)、向日葵(Helianthus annus)在各種金屬污染土壤中進行植物提取試驗,結果表明,施肥的植物葉片中富集的As、Cu和鋅(Zn)要高于不施肥的植物[7]。由此可見,不同的營養元素對植物吸收土壤中重金屬的影響存在一定差異,會因營養元素、植物和重金屬種類的不同而有不同的影響。目前,關于氮、磷、硫影響鉻超富集植物吸收Cr(Ⅵ)的研究未見報道。因此,研究氮、磷、硫對鉻超富集植物李氏禾吸收 Cr(Ⅵ) 的影響,對于提高Cr(Ⅵ)污染土壤的植物修復效率、進一步完善植物修復技術都有重要意義。
1 材料與方法
1.1 供試材料
試驗于2016年3月至2017年1月在桂林電子科技大學花江校區生命與環境科學學院環境工程實驗室中進行。
本試驗所用李氏禾幼苗采自桂林市靈川縣靈田鄉花江邊的無污染農田。選取生長一致的李氏禾幼苗,用去離子水輕柔洗凈后,放置于人工氣候箱(LRH-550-GSI,珠江牌)中,用25% Hoaglands(霍格蘭氏)營養液進行水培培養,每3 d更換1次營養液。培養條件如下:光—暗周期為12 h—12 h,白天溫度控制在25 ℃,夜晚溫度設置為20 ℃,相對濕度控制為75%,光照度為7 000 lx。每天需要加入超純水以補充因水分蒸發而減少的培養液體積,每3 d更換1次營養液,預培養 15 d。
供試土壤取自用含鉻廢水處理的模擬人工濕地試驗系統。待土壤自然風干、用瑪瑙研缽研磨并過100目尼龍篩后備用。參照HJ 491—2009《土壤 總鉻的測定 火焰原子吸收分光光度法》[8],采用微波消解儀(ETHOS A,Milestone,意大利)對土樣進行消解,用原子吸收分光光度計(AA-6300,日本島津)采用火焰法測定消解液中的鉻濃度(mg/L)。供試土壤的總鉻含量為1.80 mg/g。
1.2 試驗方法
1.2.1 N、P、S與Cr(Ⅵ)的競爭吸收試驗 試驗前,將植物分成7組:(1)對照(用25% Hoaglands溶液培養);(2)用NO3-完全缺失的25% Hoaglands溶液培養;(3)用含有 5 mmol/L NO3-的25% Hoaglands溶液培養;(4)用H2PO4-完全缺失的25% Hoaglands溶液培養;(5)用含有5 mmol/L H2PO4-的25% Hoaglands溶液培養;(6)用SO42-完全缺失的25% Hoaglands溶液培養;(7)用含有5 mmol/L SO42-的25% Hoaglands溶液培養。將培養介質的pH值均調至5.5。48 h后,將K2Cr2O7溶液加入各組植物中,使其Cr(Ⅵ)濃度分別為20、40、60、80、100 μmol/L。每個處理設3個重復,于處理后4 h收獲植物。
將收獲后的李氏禾根系先用去離子水沖洗3次,再置于超聲波清洗儀中,用10 mmol/L乙二胺四乙酸(EDTA)溶液清洗10 min,以排除植物根系表面吸附的Cr6+離子對試驗結果的影響[9],最后用超純水清洗3次。輕柔地吸干李氏禾根部表面的水分,并用陶瓷刀將根部切下,先于105 ℃殺青 30 min,再于80 ℃烘至恒質量。用陶瓷剪將李氏禾根系樣品剪至長度為1~2 mm,得到植物干樣。
使用微波消解儀(ETHOS A,Milestone,意大利),采用美國環保局(United States Environmental Protection Agency,簡稱USEPA)的方法3050B[10]對處理好的植物干樣進行微波消解。在每個批次的試驗中,菠菜標準物質(GBW10015)也用同樣的方法進行消解,作為質控,僅當檢測濃度在標定值的95%~105%范圍內時,分析結果方可接受。
1.2.2 根系分泌物對難溶性鉻的活化試驗 試驗前,將植物分成4組:(1)對照(用25% Hoaglands溶液培養);(2)用NO3-完全缺失的25% Hoaglands溶液培養;(3)用H2PO4-完全缺失的25% Hoaglands溶液培養;(4)用SO42-完全缺失的25% Hoaglands溶液培養。在試驗第10天,光照2 h后將李氏禾取出,先用蒸餾水沖洗根部3遍,在新制備的超純水中加入幾滴0.05%百里酚溶液[11],浸泡李氏禾根部5 min,再用超純水清洗根部3次。將李氏禾放入300 mL 0.5 mmol/L CaCl2溶液中[12],每杯加入3滴0.05%百里酚溶液,放回培養箱中光照收集4 h。4 h后,將收集液用0.45 μm濾膜抽濾后于40 ℃、90 r/min旋轉蒸發濃縮,在比色管中定容至25 mL。
稱取0.200 g三氧化二鉻[分析純(AR)]、0.200 g鉻酸鉛(AR)、2.000 g土壤樣品,分別倒入裝有根系分泌物的各個比色管中。在室溫下連續振蕩(振速為150 r/min)3 h后取上清液。用0.45 μm針式過濾器過濾后待測。每組設3個平行樣。
1.3 樣品的檢測與統計分析
用原子吸收分光光度計(AA-6300,日本島津公司)采用火焰法對樣品中的鉻濃度進行測定。標準系列使用液采用鉻標準溶液[GBW(E)080257]配制而成。試驗數據以3個平行樣品測定值的算術平均值±標準偏差表示,數據用單因素方差分析法(ANOVA)進行統計分析,用最小顯著差數法(LSD)評價數據間的差異水平(P<0.05表示差異顯著)。
2 結果與分析
2.1 NO3-與Cr(Ⅵ)的競爭吸收
在5 mmol/L NO3-和NO3-缺失的條件下,考察氮對李氏禾吸收Cr(Ⅵ)的影響。由圖1可見,李氏禾對Cr(Ⅵ)的吸收速率隨著Cr(Ⅵ)濃度的增加而提高。在本試驗的Cr(Ⅵ)濃度范圍內,缺氮培養條件下李氏禾的鉻吸收速率均高于對照;在5 mmol/L NO3-的培養條件下,李氏禾的鉻吸收速率均低于對照。當營養液中的Cr(Ⅵ)濃度為20 μmol/L時,缺氮培養的李氏禾對鉻的吸收速率比對照增加了18.0%;而在 5 mmol/L NO3-培養條件下, 李氏禾對鉻的吸收速率比對照減少了25.4%。說明NO3-抑制了李氏禾根系對Cr(Ⅵ)的吸收,而氮元素的缺失則明顯促進了李氏禾根系對鉻的吸收。
為了進一步研究NO3-與李氏禾根系吸收Cr(Ⅵ)之間的關系,將缺氮、對照和5 mmol/L NO3-培養條件下李氏禾根系對Cr(Ⅵ)的吸收速率用Michaelis-Menten方程進行擬合(圖2),根據雙倒數法求得Michaelis-Menten方程的各參數,列于表1。由圖2、表1可以看出,在5 mmol/L NO3-的培養條件下,李氏禾根系對Cr(Ⅵ)的最大吸收速率(vmax)比對照降低了16.2%,但米氏常數(Km)較對照明顯提高,說明在此條件下,NO3-與Cr(Ⅵ)競爭李氏禾根部相同的吸收位點。而在缺氮條件下,李氏禾根系對Cr(Ⅵ)吸收的米氏常數較對照略小,表明在此條件下,李氏禾根系吸收位點與Cr(Ⅵ)的親和力略微提高。
2.2 H2PO4-與Cr(Ⅵ)的競爭吸收
為了研究磷與李氏禾吸收Cr(Ⅵ)之間的關系,本試驗在5 mmol/L H2PO4-和H2PO4-缺失的條件下考察李氏禾根系中的鉻含量。由圖3可見,鉻吸收率隨Cr(Ⅵ)濃度增加而增加;在本試驗范圍內的任何Cr(Ⅵ)濃度處理中,5 mmol/L H2PO4-培養條件下的鉻吸收速率均高于對照,且在缺磷培養條件下的鉻吸收速率均低于對照。當營養液中Cr(Ⅵ)濃度為100 μmol/L時,5 mmol/L H2PO4-培養條件下的鉻吸收速率比對照提高了23.9%,而缺磷培養條件下的鉻吸收速率比對照減少了14.4%。由上述分析得出,H2PO4-有利于李氏禾對Cr(Ⅵ)的吸收。
將5 mmol/L H2PO4-、對照和缺磷培養條件下李氏禾根系對Cr(Ⅵ)的吸收用Michaelis-Menten方程進行擬合(圖4)。由表2可以看出,用5 mmol/L H2PO4-培養的李氏禾根系對Cr(Ⅵ)的最大吸收速率(vmax)較對照提高了17.5%,但米氏常數(Km)較對照明顯降低了17.0%,表明在此條件下,李氏禾根系吸收位點與Cr(Ⅵ)的親和力增加。缺磷條件培養的李氏禾根系對Cr(Ⅵ)的最大吸收速率(vmax)較對照有所降低,其米氏常數(Km)與對照接近,表明缺磷對李氏禾吸收Cr(Ⅵ)存在非競爭性抑制作用。
2.3 SO42-與Cr(Ⅵ)的競爭吸收
在5 mmol/L SO42-和SO42-缺失的條件下,考察硫對李氏禾吸收Cr(Ⅵ)的影響。由圖5可見,鉻吸收素速率隨著Cr(Ⅵ)濃度增加而提高。在本試驗的Cr(Ⅵ)濃度范圍內,缺硫條件培養的李氏禾對鉻的吸收速率均高于對照,5 mmol/L SO42-條件下培養的李氏禾對鉻的吸收速率均低于對照。當營養液中Cr(Ⅵ)濃度為20 μmol/L時,缺硫條件培養的李氏禾對Cr(Ⅵ)的吸收速率較對照提高了29.8%,而5 mmol/L SO42-條件下培養的李氏禾對鉻的吸收速率比對照減少了18.8%。
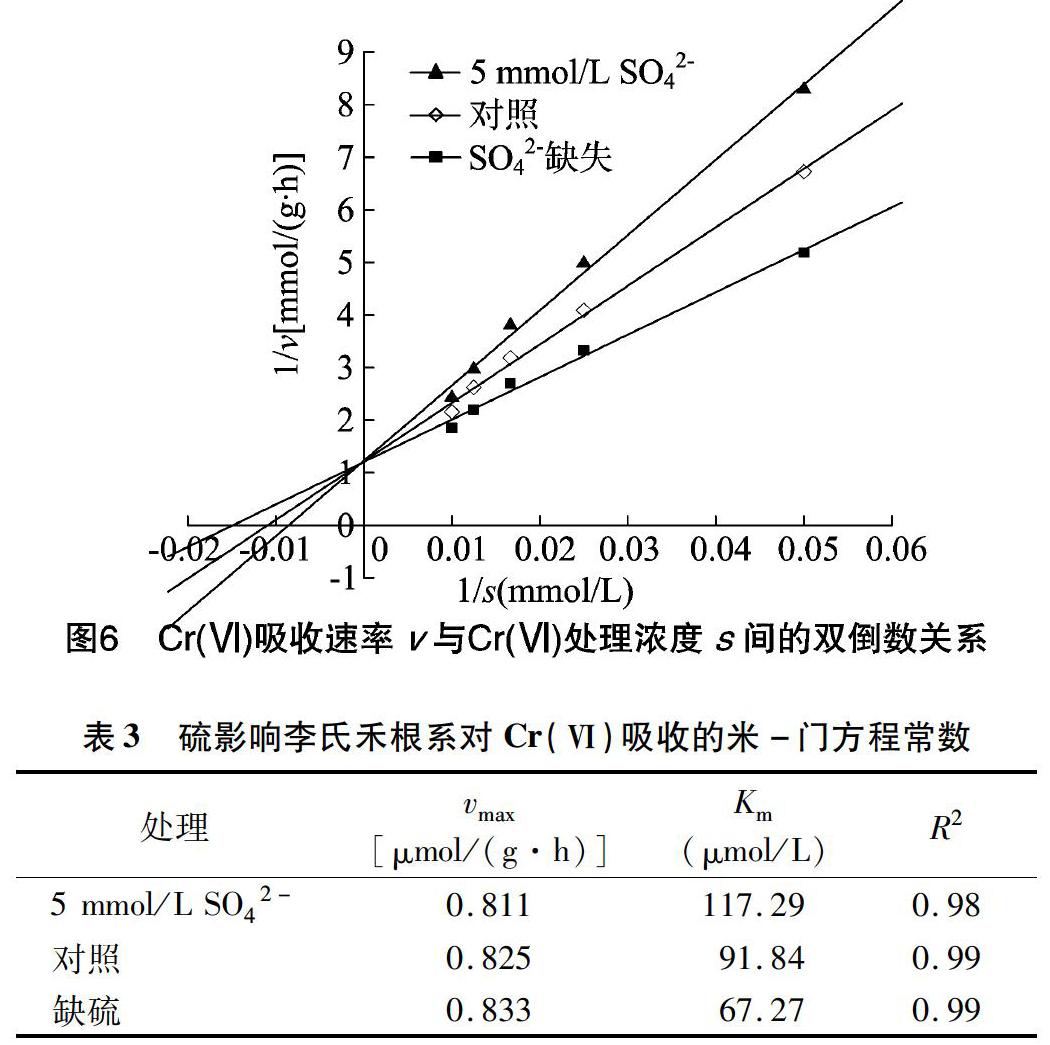
將缺硫、5 mmol/L SO42-和對照培養條件下李氏禾根系對Cr(Ⅵ)的吸收用Michaelis-Menten方程進行擬合(圖6)。由表3可以看出,5 mmol/L SO42-條件培養的李氏禾對 Cr(Ⅵ) 的最大吸收速率(vmax)與對照相近,但米氏常數(Km)較對照提高了27.7%。這表明添加SO42-對李氏禾吸收 Cr(Ⅵ) 存在競爭性抑制。在缺硫培養條件下,李氏禾根系對Cr(Ⅵ)吸收的米氏常數較對照減少了26.8%,表明在此條件下, 李氏禾的根系吸收位點與Cr(Ⅵ)的親和力增加, 即缺硫促進了李氏禾根細胞對Cr(Ⅵ)的吸收。
2.4 缺N/S/P培養對根系分泌物活化難溶性鉻的影響
為了進一步驗證李氏禾吸收Cr(Ⅵ)與非金屬營養元素氮、硫和磷之間的競爭關系,取缺氮、缺硫和缺磷培養條件下的李氏禾根系分泌物,對難溶性鉻進行活化試驗,并與全素培養條件下的李氏禾根系分泌物對難溶性鉻的活化效果進行對比。由圖7可以看出,與全素培養的李氏禾根系分泌物相比,缺氮、缺硫條件培養的根系分泌物對Cr2O3的活化量雖略微增加,但并無顯著差異(P<0.05),而缺磷條件培養的根系分泌物對Cr2O3的活化量顯著降低,較全素條件培養的根系分泌物活化量減少了29.8%。缺氮和缺硫條件培養的根系分泌物對PbCrO4的活化量與全素條件培養的根系分泌物相比的顯增加,而缺磷條件培養的根系分泌物對PbCrO4的活化量僅為全素條件培養的根系分泌物活化量的43.4%。在不同培養條件下,根系分泌物對于鉻污染土壤中鉻的活化量也存在顯著差異。缺氮和缺硫條件培養的根系分泌物比全素條件培養的根系分泌物活化效果明顯增加,而缺磷條件培養的根系分泌物校全素條件培養的根系分泌物的活化效果顯著降低。
3 討論與結論
土壤中存在的各種螯合劑及活性功能團可與重金屬競爭吸附位點,從而影響重金屬的生物有效性[13]。本試驗結果表明,NO3-與Cr(Ⅵ)競爭李氏禾根部相同的吸收位點,缺氮有利于李氏禾對Cr(Ⅵ)的吸收。據報道,森林生態系統中的植物對Pb的吸收會因為NO3-的存在而受到明顯抑制,從而減少Pb對植物的毒害作用[14]。還有研究表明,氮可以影響玉米根系對鉻的吸收,且兩者呈現負相關,即當氮含量增加時,玉米根系中的鉻含量不斷下降[15],這說明氮的存在可能會抑制玉米根系對鉻的吸收。以上結論都與本試驗的結果一致。當植物吸收NO3-時,會促使根系分泌OH-或HCO3-,使根際環境pH值升高,造成根際土壤堿化,從而降低土壤中的重金屬溶解度,減少根系對重金屬的吸收[2]。這有可能是NO3-抑制李氏禾吸收Cr(Ⅵ)的原因。
由于磷自身形態的復雜性,使得關于磷影響植物對土壤中重金屬吸收的研究存在爭議。關于磷影響植物對重金屬吸收的報道也有不同結論。本試驗結果表明,H2PO4-有利于李氏禾對Cr(Ⅵ)的吸收。這與一些學者的研究結論一致。如Singh的試驗結果表明,在含鎘土壤中加入過量的磷,可以促進油菜和燕麥對鎘的吸收[16]。黃化剛等通過盆栽試驗,發現高濃度的磷有助于東南景天對鎘、鋅的積累[17]。陳同斌等的研究發現,添加大量磷(含量為400 mg/kg以上)會使超富集植物蜈蚣草地上部、地下部的含砷量明顯升高,磷過量對蜈蚣草富集砷具有明顯的促進作用[18]。其原因有可能是磷的加入降低了土壤pH值,從而提高了土壤重金屬的有效性[19]。此外,也有研究認為,磷對植物吸收重金屬存在抑制作用。劉世亮等研究發現,磷抑制了油麥菜對鎘的吸收[20],這與本研究結論相反。由此可見,添加磷對植物吸收重金屬的影響與植物品種、重金屬種類以及磷化合物組成的不同有密切關系。
硫不僅是植物必需的營養元素,也對植物重金屬吸收調控具有重要影響。本試驗結果表明,缺硫促進了李氏禾根細胞對Cr(Ⅵ)的吸收,添加SO42-對李氏禾吸收Cr(Ⅵ)存在競爭性抑制。這與前人的研究結論一致。如Hu等研究了SO42-對水稻吸收砷的影響,發現隨著硫施加量的增加,水稻地上部分的砷累積量減少[21]。Fan等研究發現,施加過量硫能顯著降低水稻糙米中鎘的累積量[22]。Shewry等認為,硫酸根離子對大麥幼苗吸收Cr(Ⅵ)起到抑制作用[23]。其原因有可能是因為SO42-常被還原成S2-,并與土壤中其他金屬離子如Hg2+、Cu2+、Cd2+等形成金屬硫化物沉淀,從而降低金屬的移動性[24]。
根分泌現象是植物根系為適應環境變化而作出的反應[25],植物的根分泌活動會受到一些因素(如本身基因型、生長環境等)的影響,其活動過程十分復雜。上述試驗結果充分證明,李氏禾在不同營養條件(缺氮、缺硫或缺磷)下,其根系分泌物具有一定的特異性,其對特定物質的活化能力也有所不同。McGrath等通過根袋試驗證明,超富集植物天藍遏藍菜(Thlaspi caerulescens)能夠將土壤中難溶態的鋅轉化為可溶態鋅,從而有利于植物對鋅的吸收[26]。本試驗表明,在缺氮條件下,李氏禾根系分泌物能有效活化土壤中的鉻,使其活性提高,這與樓玉蘭等認為的施用硝態氮會促使根系分泌OH-或HCO3-,從降低土壤重金屬活性的研究結論[2]相符。在缺氮、缺硫培養條件下,李氏禾根系分泌物通過活化土壤中的難溶性鉻,促進植物對鉻的吸收,與本研究中缺氮、缺硫條件有利于李氏禾根系對Cr(Ⅵ)吸收的試驗結果一致,與全素培養條件相比,缺磷培養條件下的根系分泌物不利于土壤中難溶性鉻的溶出,因此不利于李氏禾根系對鉻的吸收,與本研究中缺磷對李氏禾吸收Cr(Ⅵ)存在非競爭性抑制作用的結論一致,也與趙晶等認為的磷的加入降低了土壤pH值,提高了土壤重金屬有效性的研究結論[19]相符。
綜上所述,缺氮、缺硫有利于李氏禾對Cr(Ⅵ)的吸收,缺磷不利于李氏禾對Cr(Ⅵ)的吸收,適當添加H2PO4-有利于李氏禾對Cr(Ⅵ)的吸收。在后續工作中有必要進一步研究添加不同濃度的磷對李氏禾吸收Cr(Ⅵ)的影響,以確定H2PO4-的最佳施用量。試驗結果對促進鉻超富集植物李氏禾對污染土壤中Cr(Ⅵ)的吸收,從而進一步提高鉻污染土壤的植物修復效率和完善植物修復技術有著重要的理論和現實意義。
參考文獻:
[1]Zhang X H,Liu J,Huang H T,et al. Chromium accumulation by the hyperaccumulator plant Leersia hexandra Swartz[J]. Chemosphere,2007,67(6):1138-1143.
[2]樓玉蘭,章永松,林咸永. 氮肥形態對污泥農用土壤中重金屬活性及玉米對其吸收的影響[J]. 浙江大學學報(農業與生命科學版),2005,31(4):392-398.
[3]郭光光. 氮磷施用對蓖麻吸收,轉運銅的影響及機制[D]. 武漢:華中農業大學,2017:52.
[4]López-Bucio J S,Dubrovsky J G,Raya-González J,et al. Arabidopsis thaliana mitogen-activated protein kinase 6 is involved in seed formation and modulation of primary and lateral root development[J]. Journal of Experimental Botany,2014,65(1):169-183.
[5]Shi J Y,Lin H R,Yuan X F,et al. Enhancement of copper availability and microbial community changes in rice rhizospheres affected by sulfur[J]. Molecules,2011,16(2):1409-1417.
[6]Zhong L,Hu C,Tan Q,et al. Effects of sulfur application on sulfur and arsenic absorption by rapeseed in arsenic-contaminated soil[J]. Plant Soil and Environment,2011,57(9):429-434.
[7]Marchiol L,Fellet G,Perosa D,et al. Removal of trace metals by Sorghum bicolor and Helianthus annus in a site polluted by industrial wastes:a field experience[J]. Plant Physiology and Biochemistry,2007,45(5):379-387.
[8]中華人民共和國國家環境保護標準. 土壤 總鉻的測定 火焰原子吸收分光光度法:HJ 491—2009[S]. 北京:中國環境科學出版社,2009.
[9]Leita L,Contin M,Maggioni A. Distribution of cadmium and induced Cd-bingding proteins in roots,stems and leaves of Phaseolus vulgaris[J]. Plant Science,1991,77(2):139-147.
[10]de Oliveira L M,Ma L Q,Santos J A G,et al. Effects of arsenate,chromate,and sulfate on arsenic and chromium uptake and translocation by arsenic hyperaccumulator Pteris vittata L.[J]. Environmental Pollution,2014,184:187-192.
[11]張玉鳳,馮 固,李曉林. 叢枝菌根真菌對三葉草根系分泌的有機酸組分和含量的影響[J]. 生態學報,2003,23(1):30-37.
[12]Pinto A P,Simoes I,Mota A M. Cadmium impact on root exudates of sorghum and maize plants:a speciation study[J]. Journal of Plant Nutrition,2008,31(10):1746-1755.
[13]Karlsson T,Skyllberg U. Complexation of zinc in organic soils EXAFS evidence for sulfur associations[J]. Environmental Science & Technology,2007,41(1):119-124.
[14]Burzyński M. The uptake and accumulation of phosphorous and nitrates and the activity of nitrate reductase in cucumber seedlings treated with PbCl2 or CdCl2[J]. Acta Societatis Botanicorum Poloniae,1988,57(3):349-359.
[15]李國軍. 大量營養元素對玉米苗期鉻吸收及生理特性的影響研究[D]. 太原:山西大學,2010:15.
[16]Singh B R. Cadmium and fluoride uptake by oats and rape from phosphorus fertilizers in two different soils[J]. Norwegian J Agric Sci,1990,4:239-250.
[17]黃化剛,李廷強,朱治強,等. 可溶性磷肥對重金屬復合污染土壤東南景天提取鋅/鎘及其養分積累的影響[J]. 植物營養與肥料學報,2012,18(2):382-389.
[18]陳同斌,范稚蓮,雷 梅,等. 磷對超富集植物蜈蚣草吸收砷的影響及其科學意義[J]. 科學通報,2002,47(15):1156-1159.
[19]趙 晶,馮文強,秦魚生,等. 不同氮磷鉀肥對土壤pH和鎘有效性的影響[J]. 土壤學報,2010,47(5):953-961.
[20]劉世亮,劉忠珍,介曉磊,等. 施磷肥對Cd污染土壤中油麥菜生長及吸收重金屬的影響[J]. 河南農業大學學報,2005,39(1):30-34.
[21]Hu Z Y,Zhu Y G,Li M,et al. Sulfur (S)-induced enhancement of iron plaque formation in the rhizosphere reduces arsenic accumulation in rice (Oryza sativa L.) seedlings[J]. Environmental Pollution,2007,147(2):387-393.
[22]Fan J L,Hu Z Y,Ziadi N,et al. Excessive sulfur supply reduces cadmium accumulation in brown rice (Oryza sativa L.)[J]. Environmental Pollution,2010,158(2):409-415.
[23]Shewry P R,Peterson P J. The uptake and transport of chromium by barley seedlings (Hordeum vulgare L.)[J]. Journal of Experimental Botany,1974,25(87):785-797.
[24]孫麗娟,段德超,彭 程,等. 硫對土壤重金屬形態轉化及植物有效性的影響研究進展[J]. 應用生態學報,2014,25(7):2141-2148.
[25]涂書新,孫錦荷,郭智芬,等. 植物根系分泌物與根際營養關系評述[J]. 土壤與環境,2000,9(1):64-67.
[26]McGrath S P,Shen Z G,Zhao F J. Heavy metal uptake and chemical changes in the rhizosphere of Thlaspi caerulescens and Thlaspi ochraleucm grown in contaminated soils[J]. Plant and Soil,1997,188(1):153-159.
