長江中下游麥區小麥地方種質千粒重全基因組關聯分析
2019-09-17 09:22:00程宇坤姚方杰蔣云峰江千濤蒲至恩魏育明陳國躍
四川農業大學學報 2019年4期
關鍵詞:關聯
程宇坤,李 健,姚方杰,蔣云峰,江千濤,李 偉,蒲至恩,鄧 梅,魏育明,陳國躍*
(1.四川農業大學小麥研究所,成都 611130;2.四川農業大學農學院,成都 611130)
小麥是世界第一大糧食作物,在全世界范圍內有三分之一的人口將小麥作為主食。在中國是僅次于水稻的第二大糧食作物,具有非常重要的基礎性地位[1]。提高小麥產量是小麥育種的最重要的目標之一。小麥的產量構成因子由單位面積穗數、穗粒數及千粒重組成,改良任何一個產量構成因子均能提高小麥產量。在產量三因子中,更多的遺傳研究集中在千粒重。小麥千粒重是典型的多基因控制的數量性狀,同時也受環境的影響。迄今為止,國內外已有大量學者利用家系連鎖作圖(family-based linkage mapping)對小麥千粒重進行了遺傳研究,并對控制小麥千粒重的QTL 區段進行了定位和遺傳解析。該方法通過雙親雜交,建立遺傳作圖群體,繪制高密度分子遺傳連鎖圖譜, 遺傳群體表型性狀,基于連鎖分析, 將控制千粒重的QTL(quantitative trait locus)定位在遺傳連鎖圖譜上[2-3]。鄭有良等[4](1993)對多小穗品系“10 阿”進行研究發現,粒重低受 5A、1B、2B、6B、2D 和 7D 等染色體上的隱性基因所控制;從其減小效應的程度來看,1B 和2B 染色體上的基因表現為強效;5A、6B、2D 和 7D 染色體上的基因表現為弱效。P.Ramya 等[5]利用RIL 群體對控制小麥千粒重進行QTL 定位分析, 定位到10 個控制千粒重的 QTL, 分布在 1A、1D、2B、2D、4B、5B和6B 染色體上。M.Golabadi 等[6]利用單標記分析方法在干旱脅迫和正常灌溉條件下進行千粒重QTL檢測,其中Xcfa2114-6A 能夠解釋千粒重表型變異率的20%;另外,在3B、7B 染色體上分別檢測到3個標記與千粒重緊密連鎖。近年來,多個研究團隊利用不同的遺傳群體均發現,在SSR 標記Xgwm332附近存在控制小麥千粒重的QTL,且均表現為主效QTL[7-12]。Li C.L.等[13]利用寧 7840×Clark 構建的 RIL群體分析在該標記附近發現了一個與小麥千粒重相關的QTL 區段QC-7AL,該區段位于小麥7AL 染色體上的IWA7406-IWA6535 之間,區間長度為6.5 cM,在不同環境中解釋千粒重的表型變異為10.9%~14.88%。孫宇慧等[14]進一步對該QTL 進行了精細定位和遺傳分析, 結果表明,QC-7AL 主要位于小麥7AL 染色體的 IWA7406-IWA5913 標記區間, 長度為3.1 cM,對應的物理距離約為5.63 Mb,其中包含了473 個SNP 標記和81 個基因。利用家系連鎖作圖方法對小麥千粒重進行遺傳解析往往受群體雙親遺傳差異的制約,只能在雙親的背景中分析遺傳差異,研究結果有一定的局限性。近年來,隨著小麥全基因組測序研究的完成及高通量分子標記的開發,另一種用于解析植物復雜性狀基因型的重要方法——自然群體關聯作圖(association mapping)應運而生。關聯作圖亦稱關聯分析,以自然群體為研究對象,以長期重組后保留下來的基因(位點)間連鎖不平衡(1inkage disequilibrium,LD)為基礎,對自然群體中目標性狀的遺傳變異與基因多樣性進行關聯,無須構建遺傳作圖群體,可直接鑒定出與表型變異密切相關且具有特定功能的基因位點或標記位點[15]。該方法可以同時檢測自然群體中的多個等位基因,具有低耗費、高精度、強實用性等優點,目前已廣泛應用于水稻、小麥、玉米、油菜等作物復雜性狀的遺傳解析中[16-18]。近年來,小麥的關聯作圖主要圍繞抗病(條銹病、葉銹病、白粉病等)、產量(株高、穗粒數、可育小穗數、千粒重)、品質(蛋白質含量、濕面筋含量等)相關性狀進行。R.R.Mir 等[19]利用關聯分析發現了11 個與小麥粒重相關聯的SSR、AFLP和 SAMPL 新標記。Zhang D.L.等[20]發現 Xgwm311-A500、Xgwml31-A110 和 Xcfe273-A306 等 23 個增加小麥籽粒數的優異等位變異。武玉國等[21]選用108對多態性高的SSR 標記對黃淮麥區175 份小麥進行關聯分析,得到23 個與株高、產量相關性狀顯著關聯的標記。此外,張國華等[22]以128 份黃淮麥區小麥品種(系)為材料,通過關聯分析表明獲得38 個在2 個或以上環境或均值下與產量相關性狀顯著關聯的位點;并進一步發掘了一批與產量相關性狀的優異等位變異。但迄今為止,尚未有關于長江中下游麥區小麥地方種質千粒重全基因組關聯分析的報道。
本研究團隊在前期研究中利用SSR 分子標記和DArT-seq 芯片技術, 對長江中下游麥區的小麥地方品種進行全基因組分子掃描,發現長江中下游麥區的小麥地方品種在分子水平上具有豐富的遺傳多樣性[23]。因此,本研究擬通過對188 份長江中下游麥區的小麥地方品種在多環境下的千粒重進行鑒定,結合覆蓋小麥全基因組的7 099 個DArT-seq標記開展關聯分析,以期發掘控制長江中下游麥區的小麥地方品種千粒重的顯著性位點或區段,為小麥產量分子輔助育種提供分子依據。
1 材料和方法
1.1 供試材料
供試的188 份長江中下游麥區小麥地方品種來自 9 個省區,包括:江蘇省(61 份)、浙江省(58份)、湖北省(34 份)、安徽省(8 份)、陜西省(7 份)、江西省(5 份)、湖南省(5 份)、上海市(4 份)、河南省(4 份)(表 1)。以上地方種質均由國家種質資源庫(北京)提供,四川農業大學小麥研究所保存。
1.2 表型鑒定
供試材料分別于2017年和2018年分別種植于四川農業大學崇州試驗基地(簡稱2017CZ、2018CZ)和綿陽農業科學院小麥試驗基地(2017MY)。按照完全隨機區組設計,每份材料3 行區、行長2 m、行距0.3 m。單粒點播,株距10 cm,每行20 粒種子,生長過程中按常規田間管理。5 月中旬成熟后,每份材料混收、脫粒、去除雜質后曬干至含水量達到14%以下。每份材料種子隨機數粒1 000 粒進行稱量,3次重復,取其平均值為其千粒重(g)。
表1 供試小麥地方種質Table 1 Wheat landraces used in this study

(續表1)
1.3 基因型鑒定
2018年,于小麥三葉期,每份材料代表性植株8 株,每株取1 片新鮮嫩葉,采用改良的CTAB 法[24]提取基因組DNA。DNA 提取后質控其純度和濃度,并將其稀釋至80 ng/μL 備用。將制備好的188 份長江中下游麥區小麥地方品種基因組DNA 送到澳大利亞 DArT-seq 公司(Diversity Arrays Technology,http://www.diversityarrays.com/) 進行全基因芯片掃描,獲得原始芯片測序數據;進而對原始數據進行質控:①剔除一致性<95%的等位基因標記;②剔除標記數據缺失率在10%以上的數據。
1.4 數據分析
通過利用Excel 對千粒重表型數據進行描述性統計;采用SPSS 16.0 軟件對表型數據進行方差及相關性分析[25];基于公式 H2=VG/(VG+VE)計算千粒重的廣義遺傳力(broad-sense heritability),其中VG 代表遺傳方差,VE 代表環境方差[26]。在研究團隊前期群體結構、 親緣關系和連鎖不平衡分析基礎上,基于3 個環境下小麥千粒重及其最佳線性無偏估計值(best linear unbiased estimates,BLUE),結合DArT 分子標記,采用一般線性模型(Q+K)(general linear model,GLM,Q+K)進行全基因組關聯分析[27-28]。獲得的顯著關聯標記位點(-log10(P-value)>3.0)進一步發掘其優異等位變異,解析各等位變異的表型遺傳效應[29]。
以LD 距離作為候選基因的預測區間, 在該距離內的關聯位點為同一個數量性狀位點(QTL)[30-31];QTL 的命名方法按照QTL+性狀+sicau+染色體。其中,QTL 以 Q 表示, 性狀以英文縮寫表示,sicau 代表四川農業大學,染色體以小麥染色體的名稱表示,位于同一染色體的相同性狀的QTL 在性狀后面用1、2 等區分。采用 Excel、R 軟件(R version 3.0.3)[32]及ggplot2 作圖。
2 結果與分析
2.1 長江中下游麥區地方種質千粒重表型特征分析
188 份長江中下游麥區地方種質千粒重在3 個環境下均表現出豐富的表型變異,其變異幅度分別為22.69~49.95 g、17.71~42.55 g、17.92~50.93 g(表 2)。進一步分析發現,來自上海的丈四紅、江蘇的鎮3-96 和浙江的松花蒲3 份地方種質在不同環境中均表現出較高的千粒重,分別為43.20、41.79 和40.67 g。基于千粒重BLUE 值發現, 共有87 份小麥地方種質千粒重>33.0 g,主要來自于江蘇(31 份)、浙江(24 份)和湖北(18 份),其余 14 份來自其他省區(圖1)。方差分析顯示千粒重在基因型間存在顯著或極顯著差異, 表明千粒重既受基因型的控制,也易受環境影響。3 個環境下千粒重的廣義遺傳力(H2)均較高,分別達到 0.90、0.86 和 0.92,揭示長江中下游麥區地方種質千粒重主要受遺傳控制。千粒重的峰度和偏度均接近0,說明千粒重基本符合正態分布,揭示千粒重是多基因控制的數量性狀(表2)。供試材料的千粒重在3 個環境中相關性均達到極顯著水平(P<0.001)。其中,2017 綿陽和 2018年崇州相關程度最高,其相關系數達到0.707 2;2017年綿陽和2017年崇州相關程度最低, 其相關系數為0.676 1(表3)。
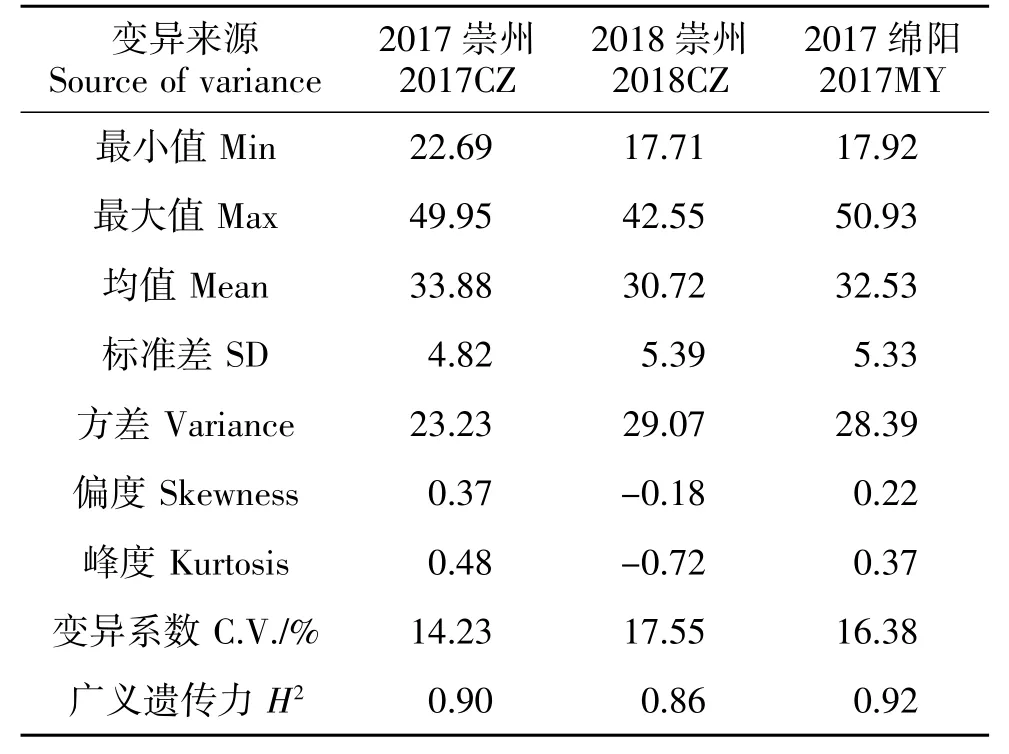
表2 188 份材料在3 個環境下的千粒重表型變異Table 2 Variation of thousand kernel weight in 188 wheat accessions detected in three environments
2.2 小麥千粒重全基因組關聯分析
本研究團隊在前期研究中, 利用DArT 芯片技術對供試材料進行全基因組分子掃描,共獲得9730個DArT 分子標記。進一步對獲得的初始標記進行質控,最終獲得7 099 個高質量的有效DArT 標記。同時,利用這些有效標記,對188 份長江中下游麥區小麥地方種質進行群體結構分析,將供試材料劃分為3 個亞群[23]。本研究在此基礎上,結合千粒重表型,基于一般線性模型(Q+K)對來自長江中下游麥區的188 份小麥地方種質進行全基因組關聯分析。
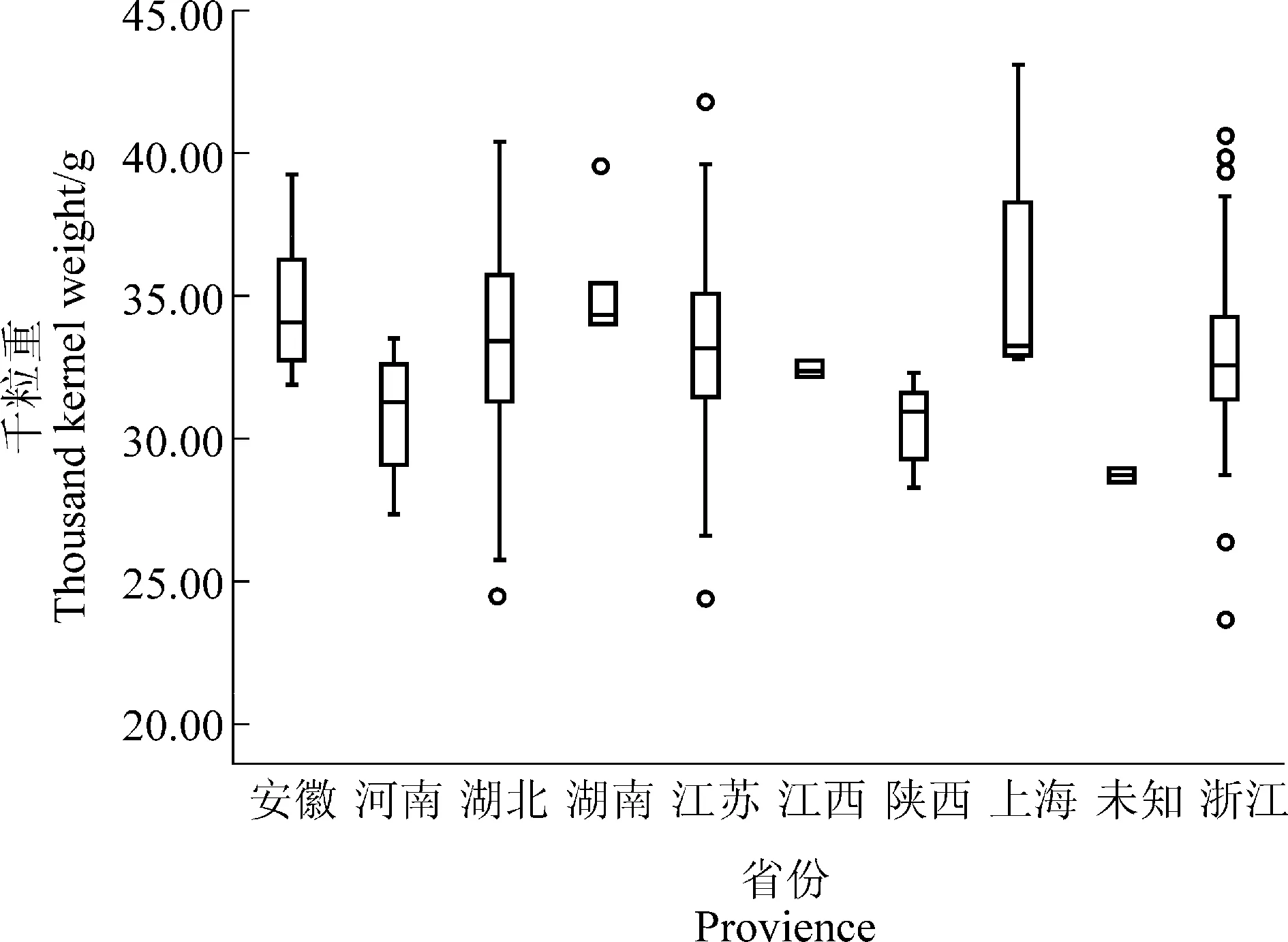
圖1 不同省份千粒重BLUE 值比較Figure 1 Comparison of thousand kernel weight BLUE value in different provinces
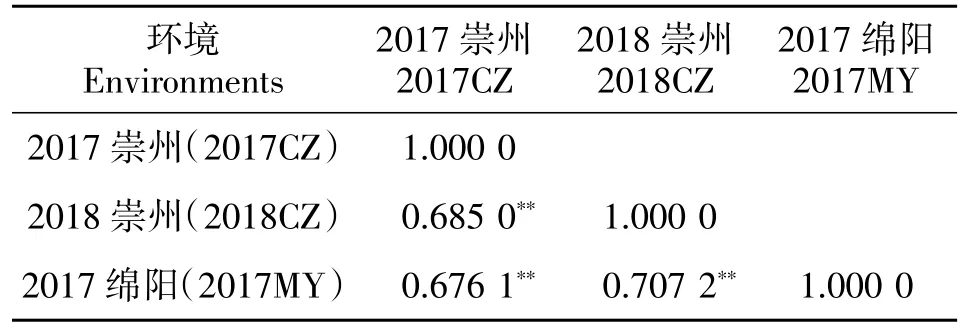
表3 不同環境下千粒重相關性分析Table 3 Thousand kernel weight correlation coefficients in different environments
通過關聯分析發現, 在2017年崇州共檢測到121 個與千粒重關聯的位點, 分布在除3D、4D、6B和6D 以外的17 條染色體上;2017年綿陽檢測到與千粒重相關的位點48 個,分布在11 條染色體上(1B、1D、2A、2B、2D、3B、4A、4B、5B、6B 和 6D);2018年崇州檢測到與千粒重相關的位點46 個,分布在1A、1B、1D、2A、2B、2D、3B、4B、5B、6B 和 6D 染色體上;與BLUE 值關聯檢測到與千粒重相關的位點6 個,分布在 1D、3B、4A 和 4B 染色體上。綜合分析 3 個環境及BLUE 值關聯位點, 共鑒定出8 個DArT 標記在2 個或2 個以上的環境中與小麥千粒重顯著相關(表4)。其中,標記 1219741 在 3 個環境下和BLUE 值中被重復檢測到,其貢獻率(R2)為10.94%。4 個位點:3533475、1252691、7348209、3939196、1095647 和 1233875,則分別在 2017年崇州、綿陽和2018年崇州3 個環境下均被重復檢測到,R2范圍從9.68%到12.65%。而1377569 則在2017年崇州和2018年崇州以及BLUE 值下被重復檢測到,其 R2為 9.19%。
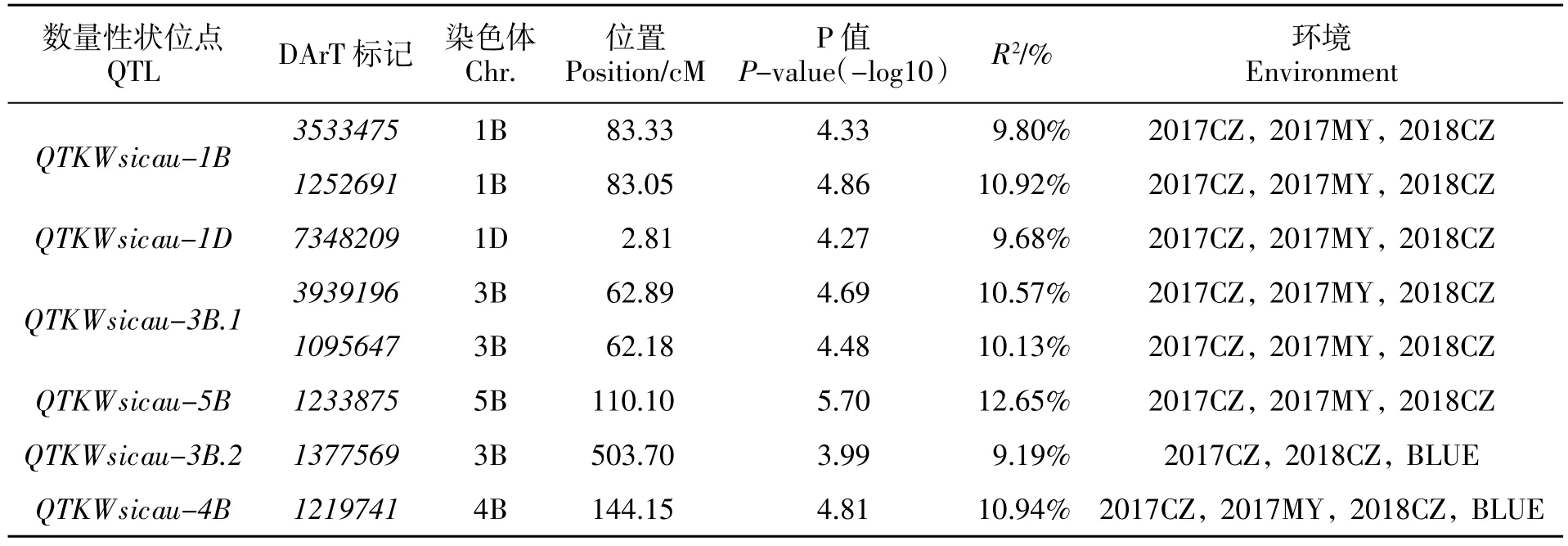
表4 與小麥千粒重顯著關聯位點Table 4 Significant loci associated with thousand kernel weight in 188 wheat accessions
2.3 控制小麥千粒重顯著關聯位點遺傳效應分析
結合表型變異,對上述獲得的8 個與小麥千粒重顯著關聯位點進行遺傳效應分析,結果發現,5 個顯著關聯位點對小麥千粒重具有正向遺傳效應。其中,1219741 可使千粒重增加1.8 g, 其在群體中出現的頻率占40%;位點3939196 可使千粒重增加0.6 g, 在群體中出現頻率為 29%;而 7348209 和3533475 均可使千粒重增加0.3 和0.1 g, 在群體中所占比例為27%和77%;1377569,千粒重增加0.2 g,所占群體比例最高為91%。另外3 個低頻率顯著關聯位點 1252691、1095647 和 1233875, 則對小麥千粒重具有負向遺傳效應,分別使千粒重降低1.6、0.3和2.2 g, 在群體中所占比例分別為24%、35%和31%(圖 2)。
3 討論與結論
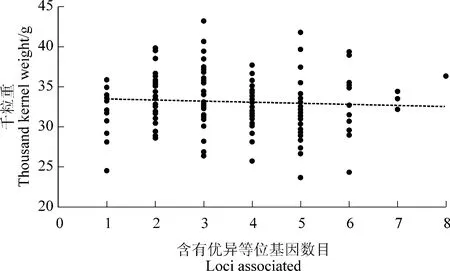
圖2 顯著關聯位點在供試材料中的分布及其與千粒重之間的線性回歸Figure 2 Distribution of the eight loci associated with thousand kernel weight contained in 188 accessions and linear regression relationships between loci associated and thousand kernel weight
千粒重是小麥產量的主要構成因素之一,該性狀受數量性狀基因控制,因此其表型選擇往往費力費時。利用分子標記輔助選擇將是一個有效的方法。在過去的十多年間,利用雙親群體已定位了較多的控制小麥千粒重QTL, 分布在小麥的1A、1D、2A、2B、2D、3A、3B、3D、4B、5A、5B、5D、6A、6D、7A和7D 染色體上[33-43]。本研究利用DArT 芯片測序技術對188 份長江中下游小麥地方種質千粒重進行了關聯分析,共鑒定出8 個與小麥千粒重穩定關聯的位點、 共涉及6 個染色體區段, 分布在1B、3B、4B、5B 和 1D 染色體上(表4)。其中,在 1B 染色體上鑒定出一個控制小麥千粒重位點QTKWsicau-1B,其解釋的表型變異率為9.80%~10.92%。蒲艷艷等[44]在1B 染色體上也發現一個控制小麥千粒重的區段QTgw.1-1B,通過標記共線性比較分析發現,該區段與本研究獲得的與小麥千粒重關聯位點QTKWsicau-1B 距離較近, 很可能是同一控制小麥千粒重的位點(區段)。廖祥政等[45]以人工合成小麥Am3 為供體親本, 普通小麥萊州953 為輪回親本,通過連續回交和自交,構建一個含85 個株系的F2∶3群體,并利用該群體,檢測到3 個千粒重QTL。其中一個QTL(QGw.caas-4B)位于被定位在染色體4BL上標記 Xgwm513~Xwmc657 區間內。張學勇等[17]也在該區間內檢測到提高千粒重1.91 g 的QTL。本研究在4B 染色體上也鑒定到另一個與小麥千粒重顯著關聯的位點QTKWsicau-4B, 其可解釋的表型變異率為10.94%。另外,張桂芝和高明剛[46-47]分別在附近區域上發現與穗粒數相關的QGns-4B 以及與千粒重相關的QTgw-4B.2。參照和對比D.Somers 等[48]小麥遺傳連鎖圖譜和DArT 分子標記高密度連鎖圖譜, 盡管本研究鑒定的QTL 與前人研究的QTL 在相近區域內, 但其定位位置并不重疊。因此推測,QTKWsicau-4B 是一個不同于已鑒定的控制小麥千粒重QTL。有意思的是,在位于1D 染色體上,我們鑒定了另一個控制小麥千粒重的QTL(QTKWsicau-1D)。在這個區段內,蒲艷艷和高明剛等[44,47]人發現該區段與小麥產量相關性狀有關,并分別鑒定出控制小麥每平米穂數相關位點QSn.1-1D 和千粒重的QTgw-1D。前人對該區段進行研究顯示,該區段與小麥條銹病抗性也有關聯,并分別定位了QYr.caas-1DS_Naxos、QYrst.orr-1DS_Stephens、QYr.sun-1D_CPI133 和QYrdr.wgp-1DS_IWA226 等控制條銹病抗性的QTL[49-53]。在本研究中,在3B 染色體上共鑒定出2 個控制小麥千粒重關聯位點:QTKWsicau-3B.1 和QTKWsicau-3B.2,在這兩個位點附近,也發現其與條銹病抗性基因相關[42-43]。這一現象揭示這些QTL 區段可能不僅僅控制著單一目標性狀,它們既控制著產量相關性狀,同時還攜帶著抗病基因或位點。
猜你喜歡
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:30
當代陜西(2021年17期)2021-11-06 03:21:36
原道(2020年2期)2020-12-21 05:47:06
當代陜西(2019年15期)2019-09-02 01:52:00
中國非營利評論(2018年2期)2018-06-18 10:48:50
學苑創造·A版(2018年11期)2018-02-01 06:29:20
自動化學報(2017年1期)2017-03-11 17:31:17
讀者(2017年5期)2017-02-15 18:04:18
西藏科技(2016年5期)2016-09-26 12:16:39
振動工程學報(2015年1期)2015-03-01 01:15:42
