作物種子的營養成分與其萌發特性關系的探討
2019-09-11 02:41:38王子勝
種子 2019年8期
單 瑩 王子勝
(1.遼寧省經濟作物研究所, 遼寧 遼陽 111000; 2.沈陽農業大學農學院, 沈陽 110161)
1876年,Nobbe提出種子生長力即種子活力的概念,從而創立了種子學[1]。科學家圍繞種子的發生和發育、結構和成分[2-3]、生理生化特性[4-5]、對環境的應激反應[6-9]等開展了一系列的研究。近年來,有關種子的信號調控系統[10-12]和分子生物學研究[13-15]也在逐步深入,嘗試著從更深層次揭開種子的秘密。
種子發育和成熟的過程也是貯藏物質逐漸累積增加的過程[13],種子內的貯藏物質主要是蛋白質、脂肪、糖類、其他含氮物質[16]。研究表明,糖類是種子的主要貯藏物質,是種子生命活動的主要呼吸基質[17];脂類可以限制種子的透水性[18],對細胞膜起到保護作用;蛋白質幫助構成細胞結構,并為種子的正常代謝和發育提供氮素來源[19]。種子內的貯藏物質在不同種類、不同成熟度、不同貯藏年限的種子之間,甚至在同一作物的不同品種之間、同一品種不同產地之間都存在一定的差異。陳新軍等研究認為,不同植物種子所含成分的差異,對其活力有一定的影響[20]。陳素傳等通過對栓皮櫟的研究認為,不同種源間栓皮櫟種子的長度、寬度和種子千粒質量以及可溶性糖、淀粉和蛋白質含量差異極顯著[21]。張瑩等[22]對山白樹種子、烏璟等[23]對白木香種子、鄔忠康等[24]對不同品種荷蘭豆種子、呂曉梅等[25]對不同產地的長柱紅山茶種子、相微微等[26]對不同地區大明綠豆種子的營養成分進行了分析比較,也得到了相似的結論。
種子萌發是一個連續的過程,包括吸水膨脹,代謝恢復,胚根突破外圍包被組織,即完成萌發[10]。種子萌發過程中,會發生一系列生理生化變化,其中包括有毒有害物質的積累、酶活性的變化、激素水平的變化等等。有關種子萌發期間酶活性和激素水平變化的研究較多,并取得了豐富的成果[27-28]。關于,種子萌發過程中的生理生化變化與種子的營養成分含量之間,相關性研究記載不多,相關研究較多以牧草、園藝作物或者樹木的種子為模板,針對大田作物種子開展的研究有限。因此,本試驗選擇具有代表性的8種大田農作物種子,檢測其貯藏物質含量和萌發期間生理生化變化等指標,研究分析了大田作物種子萌發的規律,以期為農業生產過程中種子生產、保存和應用提供參考。
1 材料與方法
1.1 試驗材料
試驗選取了棉花、紅小豆、水稻、玉米、黃豆、黑豆、花生7種作物種子為材料。
1.2 方 法
1.2.1種子基本指標的測定
每份樣品取適量,檢測百粒重,并利用培養箱進行發芽試驗。
1) 種子百粒重。
選擇大小均勻、無破損的種子(棉花種子為脫絨后光籽),測量百粒重,每樣品重復3次,取平均值。
2) 種子發芽試驗。
每樣品取100粒種子,使用發芽盒,溫度28 ℃、濕度40%,利用培養箱進行發芽試驗。發芽前利用95% 乙醇溶液對種子、發芽盒和培養箱進行表面消毒,在第4天測定發芽率。
1.2.2種子營養成分含量的測定
利用索氏提取法測定種子的粗脂肪含量,用蒽酮顯色法測定種子的可溶性糖和淀粉含量。操作方法參考張治安、陳展宇編寫的《植物生理學實驗技術》[29]。
1.2.3種子萌發期間酶活性的測定
每樣品種子稱取1 g,經全面消毒后,使用發芽盒在溫度28 ℃、濕度40%的條件下,利用培養箱進行發芽。分別在發芽1、4、7 d,取發芽種子測定丙二醛(MDA)含量、過氧化物酶(POD)活性、過氧化氫酶(CAT)活性和超氧化物歧化酶(SOD)活性。所有指標的測定均重復3次,最終結果取平均值。
采用TBA比色法測定種子MDA含量,采用愈創木酚法測定種子POD活性,采用紫外吸收法測定種子CAT和SOD活性。操作方法參考李玲等編寫的《植物生理學模塊實驗技術》[30]。
1.2.4數據處理
利用Excel軟件處理數據和作圖,利用DPS軟件分析。
2 結果與分析
2.1 種子主要營養成分含量
由不同種類種子營養成分含量差異(表1)可見,各樣本間養分含量差別明顯。其中粗脂肪含量較低(0.51%~4.62%)的小豆、玉米和水稻種子中,淀粉含量明顯較高(38%~59%),即淀粉類種子;而其它樣本的粗脂肪含量高(16%~45%),淀粉含量明顯較低(3.1%~5.0%),即脂肪類種子(油料作物種子)。除棉花和花生的可溶性糖含量較低外,其它作物種子的可溶性糖含量基本接近,與其它2種成分之間并沒有表現出相關的趨勢。
表1 不同種類種子營養成分含量
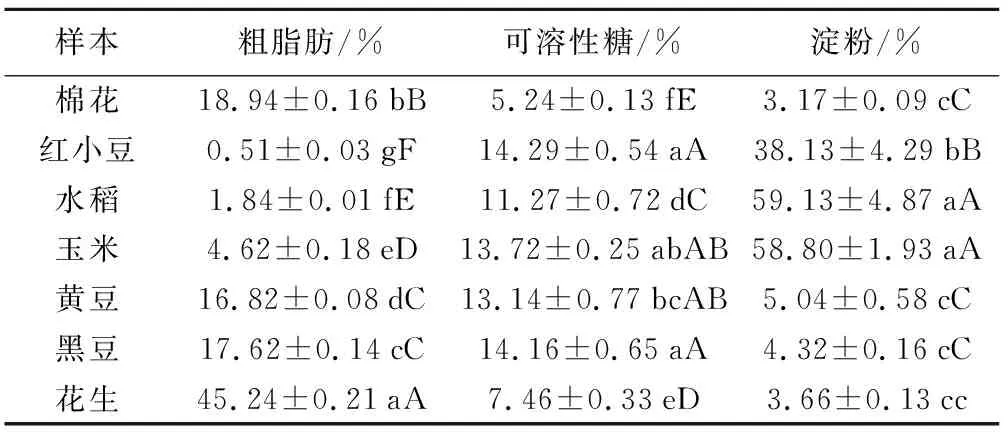
樣本 粗脂肪/%可溶性糖/%淀粉/%棉花18.94±0.16bB 5.24±0.13fE3.17±0.09cC紅小豆0.51±0.03gF14.29±0.54aA38.13±4.29bB水稻1.84±0.01fE11.27±0.72dC59.13±4.87aA玉米 4.62±0.18eD13.72±0.25abAB58.80±1.93aA黃豆16.82±0.08dC13.14±0.77bcAB5.04±0.58cC黑豆17.62±0.14cC14.16±0.65aA4.32±0.16cC花生45.24±0.21aA7.46±0.33eD3.66±0.13cc
注:表中大寫字母表示在1%水平上差異顯著,小寫字母表示在5%水平上差異顯著。下同。
2.2 種子百粒重和發芽率及其與養分含量的相關分析
2.2.1種子百粒重和發芽情況
由種子百粒重和發芽試驗的結果(表2)可見,樣本間的百粒重和4 d發芽率差別明顯。通過計算,種子的百粒重和4 d發芽率之間的相關系數為0.722 6,為中度正相關;淀粉類種子和脂肪類種子的發芽率之間也沒有表現出明顯的差異。
表2 樣本百粒重和4 d發芽率

樣本百粒重/g4d發芽率/% 棉花9.89±0.06fE8.62±1.24dC小豆21.77±0.50dD70.67±3.21abA水稻2.40±0.06gF42.33±14.57cB玉米27.90±0.69bcBC70.00±8.00abA黃豆27.03±1.42cC57.67±4.51bcAB黑豆29.47±0.91bB55.67±8.74bcAB花生47.17±1.81aA76.67±2.08aA
2.2.2養分含量與種子百粒重和發芽率的相關分析
以種子的營養成分含量為自變量,以種子百粒重和4 d發芽率為因變量,進行相關分析和逐步回歸分析,結果(表3)表明,樣本的百粒重主要受到粗脂肪和可溶性糖含量的影響,并與粗脂肪含量呈中度正相關;3種成分含量都會影響到4 d發芽率,其與粗脂肪含量呈較弱的低度相關。由此可見,粗脂肪含量對種子百粒重和發芽率的影響較大。
表3 養分含量與百粒重和發芽率的相關和逐步回歸分析
2.3 種子發芽期間MDA濃度變化及其與養分含量的相關分析
2.3.1MDA濃度變化情況
MDA濃度變化(圖1)表明,發芽期間,棉花種子的MDA濃度明顯高于其他作物,其次是花生、黑豆、黃豆,濃度最低的是水稻種子。總體來說,,脂肪類種子的MDA濃度要高于淀粉類種子,隨著發芽時間的延長,淀粉類種子MDA濃度呈升高趨勢,而脂肪類種子MDA濃度則呈先升再降趨勢。
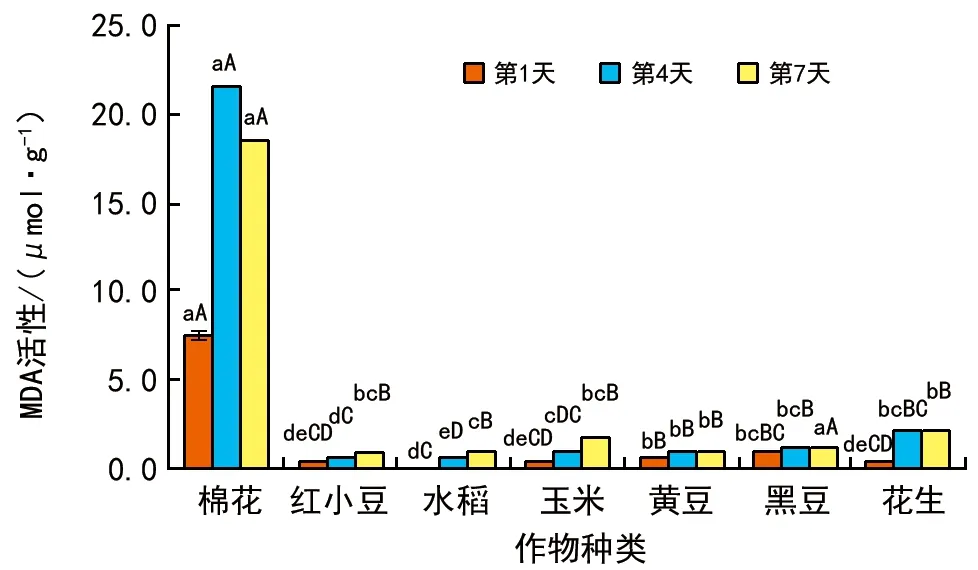
圖1 種子發芽期間MDA濃度變化情況
2.3.2養分含量與MDA濃度的相關分析
以種子的營養成分含量為自變量,以種子發芽第1天、第4天和第7天的MDA濃度為因變量,進行相關分析和逐步回歸分析,結果(表4)表明,種子發芽期間,MDA濃度主要受到可溶性糖含量的影響,并與其呈顯著負相關;其次是淀粉含量;粗脂肪含量對MDA濃度的影響較小。
表4 養分含量與MDA濃度變化的相關和逐步回歸分析

發芽時間/d粗脂肪/%X1可溶性糖/%X2淀粉/%X31 0.1192-0.7028-0.3911Y=7.45-0.52×X240.1803-0.7765-0.3792Y=22.76-1.65×X27 0.1408-0.7701-0.3160Y=19.47-1.397×X2
2.4 種子發芽期間POD活性變化及其與養分含量的相關分析
2.4.1POD活性變化
POD活性的變化(圖2)表明,種子發芽期間,淀粉類種子的POD活性呈逐漸降低趨勢,其中玉米和水稻種子發芽1 d時的POD活性明顯高于其他作物,其下降也較為明顯。脂肪類種子POD活性變化趨勢相對較復雜,但黑豆和花生POD活性一直保持在較高水平。從現有數據看,不同樣本間、不同的發芽時間之間,都沒有表現出明顯的規律。
2.4.2養分含量與POD含量的相關分析
以種子的營養成分含量為自變量,以種子發芽第1天、第4天和第7天 POD活性為因變量,進行相關分析和逐步回歸分析,結果(表5)表明,發芽1 d時, POD活性主要受到淀粉含量的影響,呈顯著正相關,與粗脂肪含量呈顯著負相關,與可溶性糖含量呈低度正相關。發芽4 d和7 d時表現出相同趨勢:主要受到粗脂肪含量的影響,并與其呈顯著到極顯著正相關,與可溶性糖和淀粉含量呈顯著負相關,且其相關性逐步減弱。

圖2 種子發芽期間POD活性變化情況
表5 養分含量與POD活性變化的相關和逐步回歸分析

發芽時間/d粗脂肪/%X1可溶性糖/%X2淀粉/%X31 -0.54410.39410.7562Y=0.0497+0.007×X34 0.7845-0.5723-0.3181Y=0.0180+0.00538×X170.8314-0.3811-0.4192Y=-0.025+0.0072×X1
2.5 種子發芽期間CAT活性變化及其與養分含量的相關分析
2.5.1CAT活性變化情況
CAT活性變化(圖3)表明,棉花種子發芽1 d和4 d 時CAT活性較高,而花生種子在發芽7 d時CAT活性較高。棉花和花生種子的CAT活性在整個試驗期間都保持在相對較高水平,而水稻和小豆種子的CAT活性則相對較低。總體來說,除棉花外,其它樣本發芽期間CAT活性呈逐漸上升的趨勢。
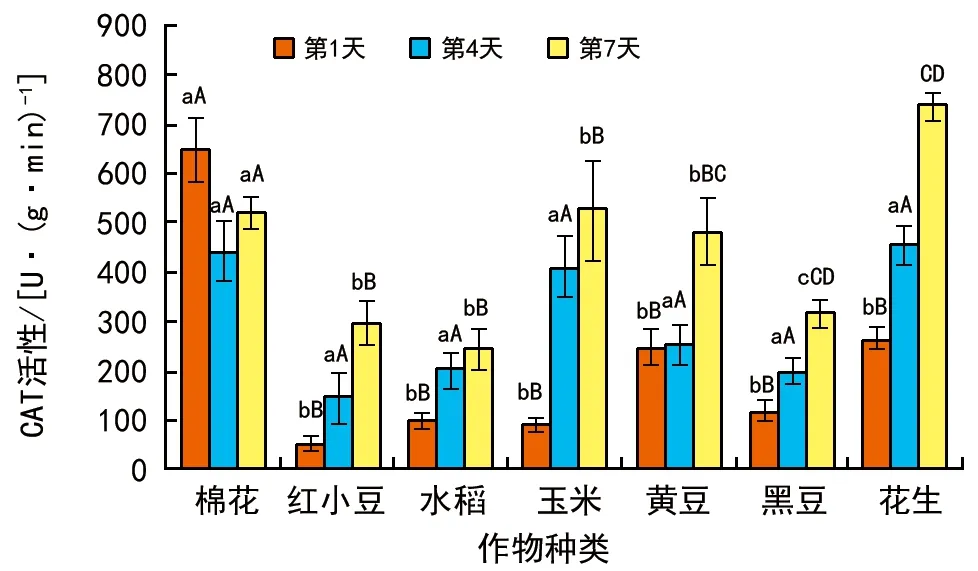
圖3 種子發芽期間CAT活性變化情況
2.5.2養分含量與CAT活性的相關分析
以種子的營養成分含量為自變量,以種子發芽第1、4、7天的CAT活性為因變量,進行相關分析和逐步回歸分析,結果(表6)表明,種子發芽期間,CAT活性與粗脂肪含量呈顯著正相關,與可溶性糖含量呈顯著或極顯著負相關,與淀粉含量呈顯著到減弱的負相關關系。逐步分析的結果也表明,種子發芽期間,CAT活性主要受到可溶性糖含量的影響。
表6 養分含量與CAT濃度變化的相關和逐步回歸分析

發芽時間/d粗脂肪/%X1可溶性糖/%X2淀粉/%X310.4098-0.8559-0.5332Y=770.9243-49.0648×X24 0.3933-0. 4801-0.0263Y=493.5094-7.7616×X1-3.4011×X270.5695-0.7847-0.4058Y=305.5121+8.7686×X2
2.6 種子發芽期間SOD活性變化及其與養分含量的相關分析
2.6.1SOD活性變化情況
SOD的活性變化(圖4)表明,棉花種子的SOD活性在發芽1 d時較高,而花生種子SOD活性在發芽4 d和7 d時較高。比較之下,棉花和花生的SOD活性相對較高,而小豆和水稻SOD活性較低。總體來看,種子發芽期間SOD活性隨發芽時間的延長而逐漸降低,并且脂肪類種子SOD活性高于淀粉類種子。
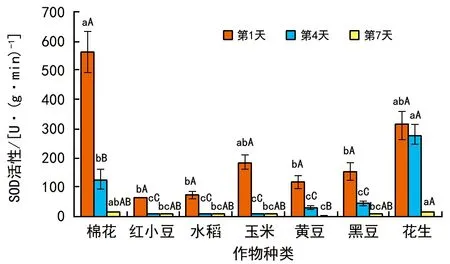
圖4 種子發芽期間SOD活性變化情況
2.6.2養分含量與SOD活性的相關分析
以種子的營養成分含量為自變量,以種子發芽第1、4、7天的SOD活性為因變量,進行相關分析和逐步回歸分析,結果(表7)表明,發芽期間SOD活性與3種成分的含量都有較強的相關關系,其與粗脂肪含量為正相關,與可溶性糖和淀粉含量呈負相關。逐步回歸分析表明,種子發芽期間,SOD活性主要受到可溶性糖含量的影響,隨可溶性糖含量的升高而降低。
表7 養分含量與SOD活性變化的相關和逐步回歸分析

發芽時間/d粗脂肪/%X1可溶性糖/%X2淀粉/%X310.5174-0.8356-0.5317Y=691.0671-41.0613×X240.9095-0.7531-0.5135Y=104.8730+4.7174×X1-9.8172×X270.6259-0.8343-0.2612Y=20.6640-1.1515×X2
3 結論與討論
3.1 不同種子養分含量表現不同的特性
試驗中各樣本的養分含量差異明顯。3種營養成分中,粗脂肪含量與淀粉含量呈負相關,可溶性糖含量與其它2種成分之間沒有表現出相關關系。脂肪含量較高的種子被稱為脂肪類種子或油料作物種子(脂肪含量≥15%),是天然油脂的主要來源,本試驗中包括花生、棉花和黃豆;而淀粉含量較高的種子被稱為淀粉類種子(淀粉含量≥35%),一般作為人畜的淀粉類食物來源,本試驗中包括水稻、玉米和小豆。值得注意的是,黑豆作為雜豆類的一種,一直都被認為是淀粉類種子,試驗檢測的脂肪含量高達17.62%,基本與黃豆持平,而淀粉含量僅有4.32%,因此應將其作為油料作物,成為植物油脂的一個新來源。同時,黑豆作為一種比較受歡迎的保健食品,建議對其油脂的組成成分進行進一步分析,充分開發其藥食同源的新用途。
3.2 種子百粒重和4 d發芽率受到營養成分含量的影響
試驗中樣本間的百粒重和第4天發芽率區別明顯,并且呈顯著正相關關系,即種子越大,發芽率越高;2個指標在淀粉類種子和脂肪類種子之間沒有表現出區別。2個指標更多受到粗脂肪含量的影響,可溶性糖含量和淀粉含量對其影響有限。然而,水稻種子由于種皮較堅厚,發芽較慢,第4天發芽率僅為42%,棉花種子的4 d發芽率僅為8%,其原因有待進一步探討。
3.3 種子萌發期間MDA主要來自于脂類物質的分解和轉化
本試驗中,脂肪類種子MDA濃度要高于淀粉類種子。隨著發芽時間的延長,淀粉類種子MDA濃度呈升高的趨勢,而脂肪類種子MDA濃度則呈先升再降趨勢。總體來看,MDA濃度隨粗脂肪含量的增加而升高,隨可溶性糖和淀粉含量的增加而降低。
可溶性糖是種子發芽到轉入光合作用之前的主要呼吸底物[6],可溶性糖含量高,則種子的生命活動相對旺盛,有害物質積累隨之降低。本試驗中,MDA含量與可溶性糖含量呈負相關,這一點與前人的研究結果相符合。趙娟等研究認為,種子中脂類物質限制種子的透水性,對細胞膜起到保護作用;但是脂類容易變質,從而引起種子的劣變[18]。本試驗中脂肪類種子MDA濃度要高于淀粉類種子,結合筆者的相關試驗(數據整理中),可以推測,脂類是種子發芽過程中MDA的重要來源之一。
3.4 種子發芽期間酶活性的變化特點
發芽過程中,淀粉類種子的POD活性呈逐漸下降趨勢,脂肪類種子POD活性變化較為復雜,該指標與粗脂肪含量呈正相關關系。CAT活性呈逐漸上升趨勢,SOD活性呈逐漸降低趨勢,二者主要隨可溶性糖含量的增加而降低。脂肪類種子的SOD活性高于淀粉類種子,CAT活性在兩類種子中沒有表現出明顯趨勢。
本試驗中,粗脂肪含量與POD活性、CAT活性和SOD活性均表現較強的正相關關系,由此可以推測,發芽過程中的氧自由基主要來自于脂類物質的分解和轉化,氧自由基的增加反而提高了酶的活性。樣本中, POD活性較高的花生和黑豆,其發芽和生長情況良好;同為脂肪類種子的棉花其POD活性明顯低于花生,發芽率也低于花生;可以認為,POD對種子起到了直接的保護作用。同時,棉花和花生的CAT和SOD活性一直較高,但花生的發芽率較高,而棉花的發芽率卻偏低,說明CAT和SOD對種子的保護作用并沒有直接體現在種子活力上。關于其作用機制,有待進一步探討。
3.5 種子發芽期間生理指標的測定應在吸水完成后進行
本試驗中,MDA、POD 、CAT、SOD 4個指標在發芽第1天時,起伏較大,在發芽第4天和第7天時,各指標都表現出一定的趨勢。由此認為,種子在發芽第1天時,還處在吸脹階段,基本的生理和代謝功能還沒有恢復。發芽第4天和第7天時,種子的活性已經被充分調動起來,代謝過程基本恢復,此時的檢測結果反映的才是種子本身的活力水平,也更值得信賴。
本試驗中種子發芽率與酶活性水平的關系,由于部分樣本出現腐爛現象,發芽率數據不具有代表性,因此沒有進行比較。關于這方面的工作,計劃在新一輪的試驗中,通過不同溫濕度條件下,種子的發芽率和酶活性水平之間的關系研究來進一步確定。
3.6 種皮對種子起到明顯的保護作用
本試驗中,為防止雜菌污染,對發芽盒、發芽箱及種子進行了表面消毒,但種子發芽率一直偏低,第4天開始出現腐爛現象,第7天時腐爛現象已經很嚴重,同時黃豆樣品也出現較嚴重的腐爛現象。樣品中順利發芽且發育良好,基本沒有腐爛的是水稻和花生。
Loic研究認為,種皮是種子抵御外界不良環境的第一層保護機制,種皮中的黃酮類物質不僅能為種子提供一個抵御病害的天然屏障,還能清除氧自由基[31]。結合本試驗的實際,認為棉花種子在脫絨過程中,種皮受到損傷,對種子的保護能力下降。徐敏等[9]曾經利用棉花脫絨和未脫絨的種子進行發芽試驗,也發現脫絨棉籽并未如預期在發芽方面占有優勢,推測應該是同樣的原因。水稻的種皮較厚,保護性較強,因而水稻發芽雖然稍慢,但發芽率較高,發芽后長勢良好且沒有腐爛現象。黃豆的種皮相對較薄,保護能力有限,因此黃豆也出現了較嚴重的腐爛現象。但是花生同樣作為脂肪類種子且種皮較薄,卻能夠順利發芽并保持良好的生長狀態。可見,種皮的薄與厚并不代表其保護能力,種皮對種子的保護機制值得進一步探討。
種子只有正常萌發和生長才能真正體現其價值,綜合本試驗研究結論,淀粉類種子在萌發方面具有一定優勢,生產中可以適當降低播種量,脂肪類種子對于萌發期間的環境條件要求較高,種皮較厚的種子出苗率較高但發芽速度慢,生產中最好采取一定的催芽引發措施,以促進其出苗。
