一個分離自貴州省茶葉上的擬盤多毛孢屬新種
2019-09-13 07:52:10陳立杰陳通政
種子 2019年8期
陳立杰, 宋 玉, 陳通政, 劉 穎, 王 勇
(貴州大學, 貴陽 550025)
擬盤多毛孢屬(Pestalotiopsis)按照最新的分類系統屬于子囊菌門,盤菌亞門,糞殼菌綱,炭角菌亞綱,黑盤孢目,被毛孢科。該真菌類群地球上普遍存在,尤其在熱帶和亞熱帶生物多樣性特別豐富[1],此外,該真菌類群不具有很高的專一性宿主,因此往往能引起許多植物的不同病害癥狀,造成不同程度的減產[2-10]。根據前人研究[4],Pestalotiopsis分類的主要形態學指標涵蓋:第一,分生孢子中間有色孢的顏色特點(包括同色,異色,橄欖綠色及褐色);第二,孢子大小范圍,形狀特點及分隔數目;第三,孢子附屬絲(頂端)的特點、基部附屬絲的有無及數目;第四,產孢器(分生孢子盤)、分生孢子梗及產孢細胞結構的特點等[4]。近年來,隨著分子生物學技術的發展,在真菌類群的分類學研究中,分子系統學技術(基于DNA序列的堿基差異的分析)發揮著越來越重要的作用且優勢明顯。本研究結合形態學和分子系統學的方法對分離自貴州茶葉上的擬盤多毛孢真菌進行分類學評價。
1 材料與方法
1.1 采集標本
采集地:花溪區黔陶鄉趙司村茶場;品種:福鼎大白茶;采集時間:2011年11月28日;標本及所獲菌株均保存于貴州大學農學院植物病理教研室標本室。
1.2 分離純化及保存菌種
菌種的分離純化及保存方法參照宋玉[11]的方法。
1.3 形態學描述
所分離獲得的菌株在25 ℃條件下培養7~10 d,等到肉眼觀察有大量油滴狀黑色或深褐色顆粒(分生孢子盤及分生孢子)出現后在體視鏡下觀察菌落形態并挑取分生孢子及分生孢子梗等制作玻片,顯微鏡(奧林帕斯BX 53)在不同放大倍數下(20×,40×,100×)觀察真菌的典型結構特征并拍攝分生孢子、產孢細胞和分生孢子盤的高清圖片,采用PhotoShop軟件選擇典型的照片組成合格的圖版,Image Frame Work軟件測量擬盤多毛孢菌不同繁殖結構的形態學指標并完成相應的形態學描述。
1.4 DNA的提取、PCR擴增及測序
1.4.1基因組DNA的提取
采用DNA快速提取試劑盒提取擬盤多毛孢菌的基因組DNA,具體步驟參考宋玉[11]的方法。
1.4.2PCR反應體系,引物及擴增程序
1) 反應體系。
PCR總反應體系為25μL:不同基因正反向引物分別1μL,Template DNA 1μL,雙蒸水 9.5μL,2倍的PCR Master Mix(2×Taq DNA聚合酶,2×PCR 緩沖液以及2×dNTP)共計12.5μL[11]。
2) 不同基因引物序列及PCR程序。

注:模式標本為HGUP 4086,其中A~H為分生孢子;I~J為分生孢子梗和產孢細胞。標尺:A~J=20 μm。圖1 中國茶擬盤多毛孢
ITS 基因DNA片段:
通用引物(ITS 4和ITS 5),引物序列見參考文獻[12]。
反應程序:95 ℃下總預變性0.5 min,然后95 ℃變性反應0.5 min,52 ℃退火反應45 s,72 ℃延伸1.5 min,反應進行35個循環后72 ℃總延伸10 min[11],反應完成后4 ℃下保存或及時取出。
β-tubulin(β-微管蛋白)基因DNA片段:
特異引物Bt2a和Bt2B,引物序列見參考文獻[13-14]。
反應程序:93 ℃總預變性3 min,然后95 ℃變性1 min,55 ℃退火50 s,72 ℃延伸1 min,反應進行35個循環后72 ℃總延伸10 min[14-15],反應完成后4 ℃下保存或及時取出。
tef1(延伸因子)基因DNA片段:
特異引物EF 1-526F和EF 1-1567 R,引物序列見參考文獻[16]。
反應程序:94 ℃總預變性5 min,然后94 ℃變性0.5 min,63到53 ℃或66到56 ℃(每循環降低1 ℃)退火50 s,共10個循環;94 ℃變性0.5 min,63或66 ℃退火55 s, 72 ℃延伸1.5 min,反應進行36個循環后72 ℃總延伸7 min[16],反應完成后4 ℃下保存或及時取出。
1.5 不同基因的DNA序列分析
將編輯好的三個基因(ITS+β-tubulin+tef1)序列加合文件在PAUP* 4.0 b 10[17]中運行,算法設定為最大簡約法(Maximum Parsimony,MP)。采用啟發式搜索(heuristic search)算法并應用樹二等分再連接選項(Tree Bisection-reconnection,TBR)來進行分枝交換,自舉法(Bootstrap)檢測設置為1000次。程序運行完成后所生成的系統發育樹的拓撲結構在Treeview軟件中顯示,選擇最優樹的拓撲結構在Figtree軟件中進行編輯。
2 結果與分析
2.1 形態學特征描述
中國茶擬盤多毛孢(Pestalotiopsiscamellia-sinensisYong Wang bis, Y. Song, & K.D. Hyde, sp. nov.)擬盤多毛孢屬真菌子實體球形,黑色,未成熟前往往寄生于植物組織表皮下方,直到成熟后方外露,呈現凸起(黑色的小顆粒)。分生孢子梗很短至不明顯,產孢細胞呈現安培瓶狀。分生孢子梭形或近梭形,偶有彎曲,具4個橫隔膜(5個細胞),大小范圍是27~35 × 7~10μm (平均值為31.5 × 8.5μm); 其中中間的3個深色胞均為淺褐色,長度為17~23μm (平均值為20μm),分隔處稍縊縮,橫膈膜顏色有加深,分生孢子的壁不光滑具點突;中間深色3細胞自下而上第一個其長度是5~8μm (平均值為6.5μm),第二個細胞的長度為6~8μm (平均值為7.4μm),第三個細胞的長度為5~8μm (平均值為7μm);中間3細胞兩端的細胞近透明,亞圓錐形,長度為5~8μm (平均值為6.5μm);其中頂端細胞具4~6根管狀的附屬絲,附屬絲往往具有分枝,長度為20~40μm (平均值為30μm);分生孢子的基細胞略鈍圓,附屬絲往往缺失。
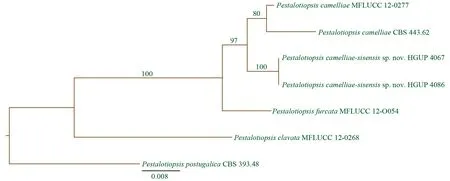
圖2 中國茶擬盤多毛孢的DNA序列分析
模式標本:貴州省貴陽市花溪區黔陶鄉趙司村,茶樹(福鼎大白茶)葉片,模式標本編號HGUP 4086,采集人:宋玉,采集時間:2011-11-28。
其他標本:貴州省貴陽市花溪區黔陶鄉趙司村,茶樹(福鼎大白茶)葉片,標本編號HGUP 4076,采集人:宋玉,采集時間:2011-11-28。
2.2 不同基因的DNA序列分析
通過ITS,β-微管蛋白以及延伸因子DNA序列構建的系統進化樹,我們發現中國茶擬盤多毛孢與山茶擬盤多毛孢(PestalotiopsiscamelliaeY.M. Zhang, Maharachch. & K.D. Hyde)和頂生分枝擬盤多毛孢(PestalotiopsisfurcataMaharachch. & K.D. Hyde)保持了比較近的遺傳進化關系,同時又保持了相對獨立的分類地位(形成3個進化分枝),分別獲得100%及97%的自舉支持率。就形態學而言,三者也存在較明顯的差別。中國茶擬盤多毛孢的分生孢子大小比頂生分枝擬盤多毛孢的小,但比山茶擬盤多毛孢的大。此外,中國茶擬盤多毛孢頂端附屬絲的數目也與其它2個種的也不相同,并且頂端附屬絲普遍會分枝,但近似種沒有表現出這些形態特征特征[6,17]。因此,結合以上DNA序列分析和形態學特征的研究,我們將中國茶擬盤多毛孢確定為擬盤多毛孢屬的一個新種。
3 討 論
中國茶擬盤多毛孢,山茶擬盤多毛孢和頂生分枝擬盤多毛孢這三個種均分離自茶樹上,其中山茶擬盤多毛孢分離自云南,頂生分枝擬盤多毛孢分離自泰國,而本研究中的新種來自貴州省[6,17]。此外,山茶擬盤多毛孢和頂生分枝擬盤多毛孢均可以引起茶樹的灰斑病,而我們這個種還沒有引起病害的直接證據。盡管形態學和分子系統學的結論已經證明我們所發現的新種與山茶擬盤多毛孢和頂生分枝擬盤多毛孢不同,然而我們不能掉以輕心,貴州省作為茶葉之鄉且種植面積全國第一[18],應該時刻關注病害的發生動態,因此我們在后續研究中會探究引起病害的可能性及危害性。
