水稻親本秈粳分化程度與雜種優勢的相關性
2019-09-10 07:22:44左生力彭玉林李齊向徐淑英鄭云峰張添運馬益虎林蕓
南方農業學報 2019年5期
關鍵詞:水稻
左生力 彭玉林 李齊向 徐淑英 鄭云峰 張添運 馬益虎 林蕓
摘要:【目的】分析水稻親本秈粳分化程度與雜種優勢的相關性,為研究雙親秈粳分化最佳配比及組配秈粳交強優勢組合提供理論參考。【方法】采用ILP分子標記和程氏形態指數法對51份水稻親本材料進行秈粳分化程度檢測,基于23個ILP分子標記進行系統聚類分析,分析兩種方法的相關性。按秈/秈、偏秈/偏秈、秈/偏秈、秈/偏粳、偏秈/偏粳和偏粳/偏粳6種類型配制150個雜交組合,綜合分析親本秈粳分化程度與雜種優勢的相關性。【結果】采用程氏形態指數法從51份水稻親本材料共檢測出秈型20份、偏秈型19份、偏粳型6份和粳型6份;利用23對擴增結果穩定、條帶清晰且只有兩種等位基因的ILP引物從51份供試親本材料中共檢測出秈型14份、偏秈型20份、偏粳型11份和粳型6份,不存在100%純秈或純粳類型。ILP分子標記法與程氏形態指數法的秈粳分類結果均與系統聚類分析結果具有較高的吻合度,但仍在差異。51份供試親本的粳稻成分指數與穗軸第1~2節長和抽穗時稃色呈顯著正相關(P<0.05),與谷粒長寬比、酚反應和程氏指數總分呈極顯著正相關(P<0.01),其中與程氏指數總分的相關系數最高,為0.81。隨著組配雙親粳稻成分的增加,6種組配類型雜交組合的生育期、株高和每穗總粒數總體呈增加趨勢,但單株有效穗數、結實率和千粒重總體呈下降趨勢,單株重則呈先增加再下降的變化趨勢。【結論】利用ILP分子標記可有效區分親本的秈粳屬性,是一種有效的水稻秈粳分類工具。采用秈/偏秈方式配組較易篩選到強優組合;雖然采用秈/偏粳方式配組獲得高產組合的概率較低,但有機會獲得極高產組合,在育種中亦可大膽嘗試。
關鍵詞: 水稻;親本;秈粳分化;ILP分子標記;雜種優勢
中圖分類號: S511.033 ? ? ? ? ? ? ? ? ? ?文獻標志碼: A 文章編號:2095-1191(2019)05-0942-08
Abstract:【Objective】The correlation between indica-japonica differentiation and heterosis was studied to provide theoretical reference for the optimum ratio of indica-japonica differentiation in parents of indica-japonica rice and inter-subspecific hybrid rice combinations. 【Method】ILP molecular marker and Cheng’s morphology index were used to detect indica-japonica differentiation level of 51 rice parents. And the correlation between the two methods was clustered by 23 ILP molecular markers. A total of 150 hybrid combinations were set based on indica/indica, partial indica/partial indica, indica/partial indica, indica/partial japonica, partial indica/partial japonica and partial japonica/partial japonica, and correlation between indica-japonica differentiation and heterosis was analyzed. 【Result】There were 20 indica types,19 partial indica types,6 partial japonica types and 6 japonica types were detected from the 51 tested parents based on Cheng’s morphology index. Twenty-three pairs of ILP primers which had stable amplification products, clear bands and two alleles were used to detect 14 indica types,20 partial indica types,11 partial japonica types and 6 japonica types out of 51 tested parents. There was no 100% pure indica or japonica types. The indica-japonica differentiation results by ILP molecular marker and Cheng’s morphology index were highly consistent with clustering analysis but still different. Indica-japonica component index of 51 test parents was significantly positively correlated with length of the first and second rachis internode and glume color at heading stage(P<0.05), and was extremely positively correlated with length-width ratio of grain, phenol reaction and Cheng’s index total score(P<0.01). The correlation with Cheng’s index total score was the highest(0.81). With the increase of parental japonica rice composition,the six types combinations all showed an overall increasing trend in growth period,plant height and total grain per panicle. But effective panicles per plant,seed-setting rate and 1000-grain weight decreased. The variation trend of single plant weight was firstly increased and then decreased. 【Conclusion】ILP molecular marker can be used to accurately detect the proportion of indica/japonica content in rice parent. It is an easy tool to classify indica and japonica rice. Fine combinations can be easily obtained through indica/partial indica combination. It is possible to obtain extremely high-yield combinations through indica/partial japonica,although the probability of obtaining high-yield combinations is low. It is worth trying in breeding.
Key words: rice; parents; indica-japonica differentiation; ILP molecular marker; heterosis
0 引言
【研究意義】秈稻和粳稻是亞洲栽培稻的兩個亞種,因其雜交后代可產生豐富變異,蘊藏著巨大雜種優勢(袁隆平等,1997;孫傳清等,2000;Li et al.,2000),因此通過綜合兩大亞種優勢進行秈粳雜交育種是進一步提高水稻產量的有效途徑,逐漸成為研究熱點(況浩池等,2007;陳芬等,2009;鄭秀娟等,2016)。然而,由于秈稻與粳稻在分類學上屬于不同的亞種,親緣關系較遠,導致雜種一代存在結實率低、植株過高、生育期偏長和籽粒充實不飽滿等問題,無法直接利用。隨著廣親和基因的發現,研究人員開始利用秈粳交培育優良雜交稻親本,目前已育成大量秈粳交水稻親本,但主要建立在隨機配組的基礎上(馬彬林,2015),導致選育效率較低。因此,研究配組雙親秈粳分化程度與雜種優勢間的相關性對選育綜合性狀優良的秈粳交衍生系(不育系或恢復系)具有重要意義。【前人研究進展】在秈粳交雜種優勢利用過程中,秈粳分化最佳配比引起普遍關注,主要從秈粳交新品種選育、不同生態條件下秈粳亞種特性、秈粳分化分子檢測、秈粳分化程度與雜種優勢關系等方面開展相關研究,并取得了長足的進展。姜廷波等(1999)分別用45個秈粳特異性RFLP分子標記和形態指數分類法分析供試親本及雜種的秈粳分化程度,結果表明兩種分類方法均可將親本和雜種分為秈和粳兩大類群。龍雯虹等(2003)以10個滇型雜交稻不育系及其相應的10個保持系和38個恢復系、不育系和恢復系組配的40個雜交種為試材,研究親本材料的秈粳分化程度與雜種優勢的相關性,結果表明親本秈粳差異與產量雜種優勢呈拋物線的關系。張建勇等(2005)利用126個SSR分子標記對42份常用雜交水稻親本材料進行秈粳分類研究,結果表明聚類結果與親本材料親緣關系基本吻合,說明SSR分子標記能較好地區分秈稻和粳稻。姜樹坤等(2006)用18個RAPD分子標記對30個水稻栽培品種進行秈粳分類研究,結果表明分類結果與品種系譜基本吻合,證明RAPD分子標記可應用于水稻的秈粳分類。張受剛和許旭明(2006)通過聚合不同廣親和基因對粳型親秈恢復材料進行改良,選育出具有良好親秈性和恢復能力的粳型親秈恢復材料gk729,在此基礎上進一步選育出具有良好親秈性和恢復能力的粳型親秈恢復材料gk419,最終育成一批綜合性狀優良的粳型親秈恢復系,并應用于生產。徐海等(2007)以中優早8號×豐錦F6重組自交系群體為試材,在遼寧和四川兩個生態區種植研究亞種特性的變化,結果表明,重組自交系群體在兩個生態區均呈偏粳分布,且亞種特性均發生明顯變化,總體表現為四川比遼寧更偏粳。許旭明等(2009)利用ILP分子標記和程氏形態指數法對18個秈粳雜交親本和39個衍生系進行秈粳分化研究,結果表明ILP分子標記可準確檢測秈(粳)成分比例,且秈粳分化可合理解釋秈粳型恢復系配組雜交稻表現出明顯雜種優勢的原因。王明軍等(2010)利用34對InDel分子標記將云南省各地的181份糯稻材料劃分為141份典型秈稻、18份秈稻、8份典型粳稻、13份粳稻和1份偏粳稻。黃明(2010)研究發現,ILP分子標記具有很強的秈粳特異性,且絕大多數只擴增出兩種條帶,具有很大的利用價值,可預測常規雜交稻產量的雜種優勢。李亞莉等(2012)利用SSR分子標記分析滇型雜交稻親本秈粳分化程度與雜種優勢的相關性,結果發現親本秈粳分化差異與產量競爭優勢無線性關系。范方軍等(2015)利用32對InDel分子標記和18對SSR分子標記將200份太湖流域水稻地方材料劃分為179份典型粳稻、17份典型秈稻、秈稻2份及粳稻和偏粳稻各1份。綜上所述,利用分子標記可有效對水稻進行秈粳分類及雜種優勢預測,尤其是ILP分子標記具有很強秈粳特異性,在雜種優勢預測方面具有很大的利用價值。【本研究切入點】前人研究多集中在利用分子標記進行秈粳分類,但秈粳分化程度與雜種優勢間的相關性研究甚少,尤其鮮見利用ILP分子標記進行兩系親本秈粳分化程度與雜種優勢間相關性的研究報道。【擬解決的關鍵問題】利用ILP分子標記檢測歷時十余載培育而成的秈粳交龍恢系列恢復系和福龍S系列不育系共51份親本材料的秈粳分化程度,探索雙親秈粳分化程度與雜種優勢間的相關性,以期明晰龍恢系列恢復系和福龍S系列不育系的秈粳分化情況,為研究雙親秈粳分化最佳配比及組配秈粳交強優勢組合提供理論參考。
1 材料與方法
1. 1 試驗材料
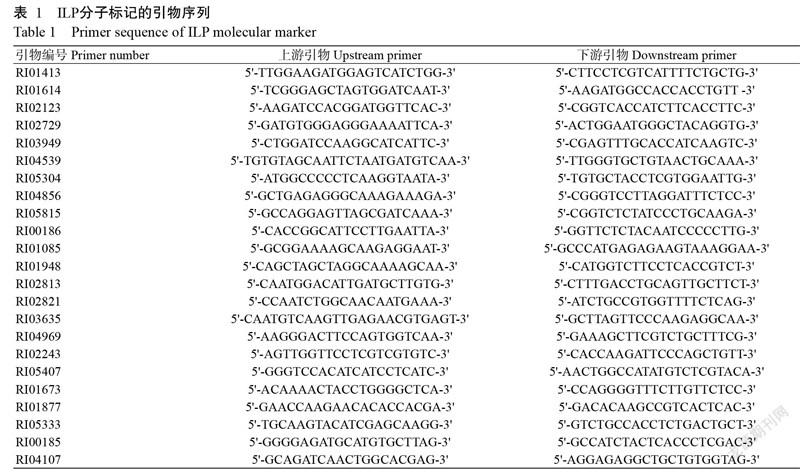
供試水稻材料為9311、蜀恢527、多系一號、明恢70、日本晴、武運粳7號、武運粳27號、鎮稻413、鹽粳48、武育粳3號、奧龍1S、12個福龍S系列不育系及28個龍恢系列恢復系。以上述材料為親本配制的150個雜交組合由福建省龍巖市農業科學研究所提供。23對ILP分子標記引物(表1)由福建省三明市農業科學院提供,由生工生物工程(上海)股分服務有限公司合成。主要試劑:核酸染料(Ultra GelRed)購自南京諾唯贊生物科技有限公司,2×Taq PCR Master Mix購自北京艾德萊生物科技有限公司,其他試劑均購自福州沃森生物技術有限公司。主要儀器:人工氣候箱(上海博迅醫療生物儀器股份有限公司)、Veriti梯度PCR儀(ABI,美國)、SCIENTIFIC電泳儀(C.B.S,美國)和Sigma3-30KS高速冷凍離心機(Sigma,德國)。
1. 2 試驗方法
1. 2. 1 親本秈粳分化程度檢測
1. 2. 1. 1 程氏形態指數法 采用程侃聲(1993)的程氏形態指數法,分別對稃毛、葉毛、抽穗時的殼色、籽粒長寬比、第1~2穗節長和酚反應等6個性狀進行評分,各性狀得分之和為程氏指數總分,以其作為分類尺度:指數值0~8分為秈型,9~13分為偏秈型,14~17分為偏粳型,18~24分為粳型。
1. 2. 1. 2 ILP標記法 采用SDS法提取DNA,將其用于PCR擴增。PCR反應體系15.00 μL:DNA模板2.00 μL,10 mmol/L Mg2+ 10×緩沖液0.75 μL,10×Buffer 1.25 μL,2.5 mmol/L dNTP 0.30 μL,50 ng/μL上、下游引物1.50 μL,5 U/μL Taq Plus DNA聚合酶0.15 μL,ddH2O補足至15.00 μL。PCR擴增程序:94 ℃預變性5 min;94 ℃ 30 s,59 ℃ 30 s;72 ℃ 1.0 min,進行10個循環;94 ℃ 30 s,56 ℃ 30 s,72 ℃ 1 min,進行10個循環;72 ℃延伸5 min,4 ℃保存。PCR產物中加入3.0 μL上樣緩沖液,上樣量3.0 μL,在SCIENTIFIC電泳儀上進行非變性聚丙烯酰胺凝膠電泳分離。凝膠成分包括0.04%過硫酸銨、0.1% TEMED和10%丙烯酰胺,電泳緩沖液為0.5×TBE,250 V電泳1.5 h。銀染法染色,定影。薄膜包好置于室溫干燥后統計條帶并照相。
1. 2. 2 農藝性狀考察 2018年春季在海南省三亞市藤橋鎮南繁基地,從14個秈型親本中選出5個不育系(福龍S3、福龍S5、福龍S16、福龍S7和奧龍1S)和5個恢復系(龍恢5928、蜀恢527、龍恢109、龍恢3381和9311),從20個偏秈型親本中選出5個不育系(福龍S8、福龍S9、福龍S2、福龍S19和福龍S12)和5個恢復系(龍恢1031、龍恢3389、龍恢20、龍恢236和龍恢29),從11個偏粳型親本中選出10個恢復系(用作母本:龍恢536、龍恢479、龍恢557、龍恢438、龍恢6315;用作父本:龍恢517、龍恢66、龍恢336、龍恢8612、龍恢779),共計30個親本。按秈/秈、偏秈/偏秈、偏粳/偏粳、秈/偏秈、秈/偏粳和偏秈/偏粳6種組配類型進行5×5不完全雙列測交(雜交),共配制150個雜交組合。2018年秋季在福建省龍巖市新羅區龍門鎮赤水村育種基地,采用隨機區組設計,3次重復,每小區單本種植32株,成熟時及時收獲并按照左生力(2016)的方法考察150個組合的生育期、株高及產量相關性狀(單株有效穗、每穗總粒、結實率、千粒重和單株重)。
1. 3 統計分析
人工統計所有清晰可辨的電泳條帶。同一引物按擴增條帶的“有”或“無”計數,遷移率相同的條帶視為1個位點,有條帶賦值為1,無條帶賦值為0,從而建立特征數據矩陣用于進一步分析。按Xi =∑Xd/(∑Xd+∑Jd),Xj =∑Jd/(∑Xd+∑Jd)公式計算秈粳成分指數,其中,Xi為秈稻成分指數(Composition index of indica rice),Xj為粳稻成分指數(Composition index of japonica rice),Xd為秈稻條帶,Jd為粳稻條帶,以成分指數計算各供試親本的秈粳分化程度。根據許旭明等(2009)的劃分標準,將粳稻成分指數≥50%的材料判定為粳型,18.0%~49.9%為偏粳型,8.0%~17.9%為偏秈型,<8.0%為秈型。
2 結果與分析
2. 1 引物篩選及供試親本擴增結果
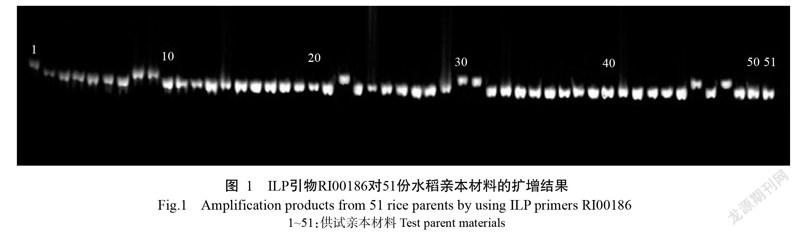
用程氏形態指數法劃分供試親本的秈粳類型,選出秈型和粳型兩種類型作為多態性條帶秈粳屬性判別依據,對許旭明等(2009)的48對ILP引物作進一步篩選,最終獲得23對擴增結果穩定、條帶清晰且只有兩種等位基因的ILP引物,利用其對51份水稻親本材料進行PCR擴增,結果顯示,每對引物均檢測到2種等位基因條帶,23對ILP引物共擴增到46條條帶(圖1)。
2. 2 供試親本秈粳分類及聚類分析結果
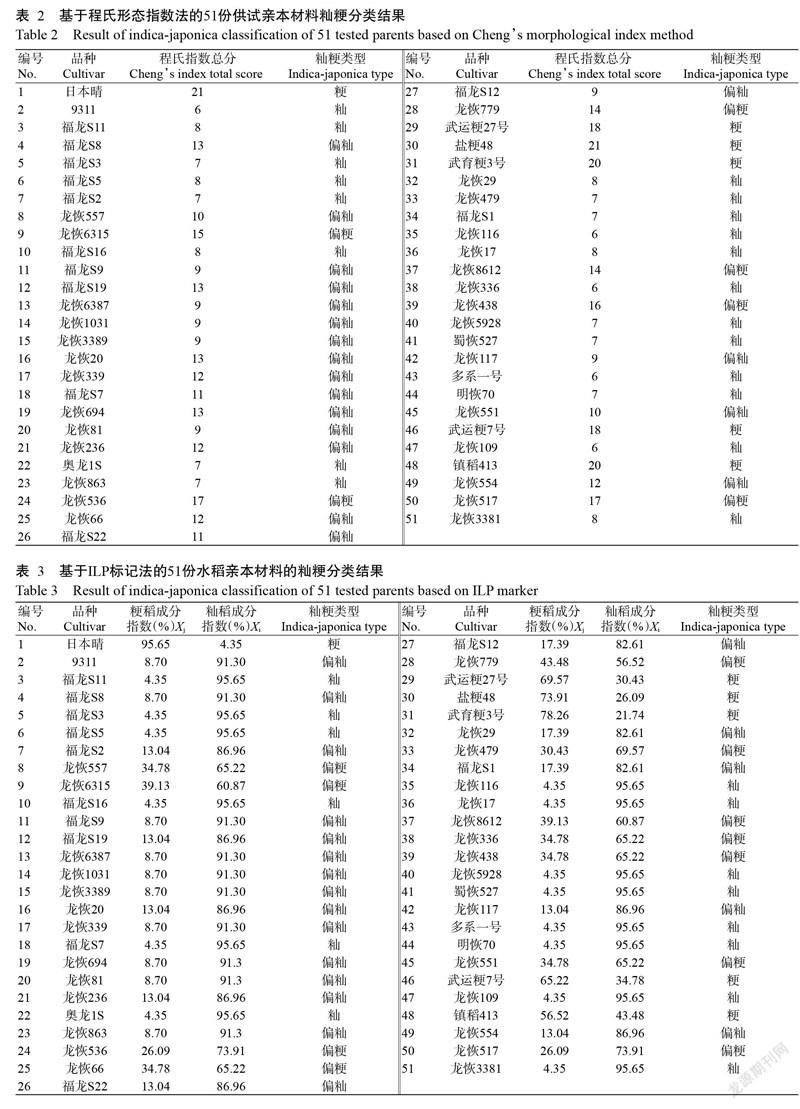
2. 2. 1 程氏形態指數法 采用程氏形態指數法對51份水稻親本材料進行秈粳類型劃分,結果如表2所示。從51份水稻親本材料共檢測出秈型20份、偏秈型19份、偏粳型6份和粳型6份。
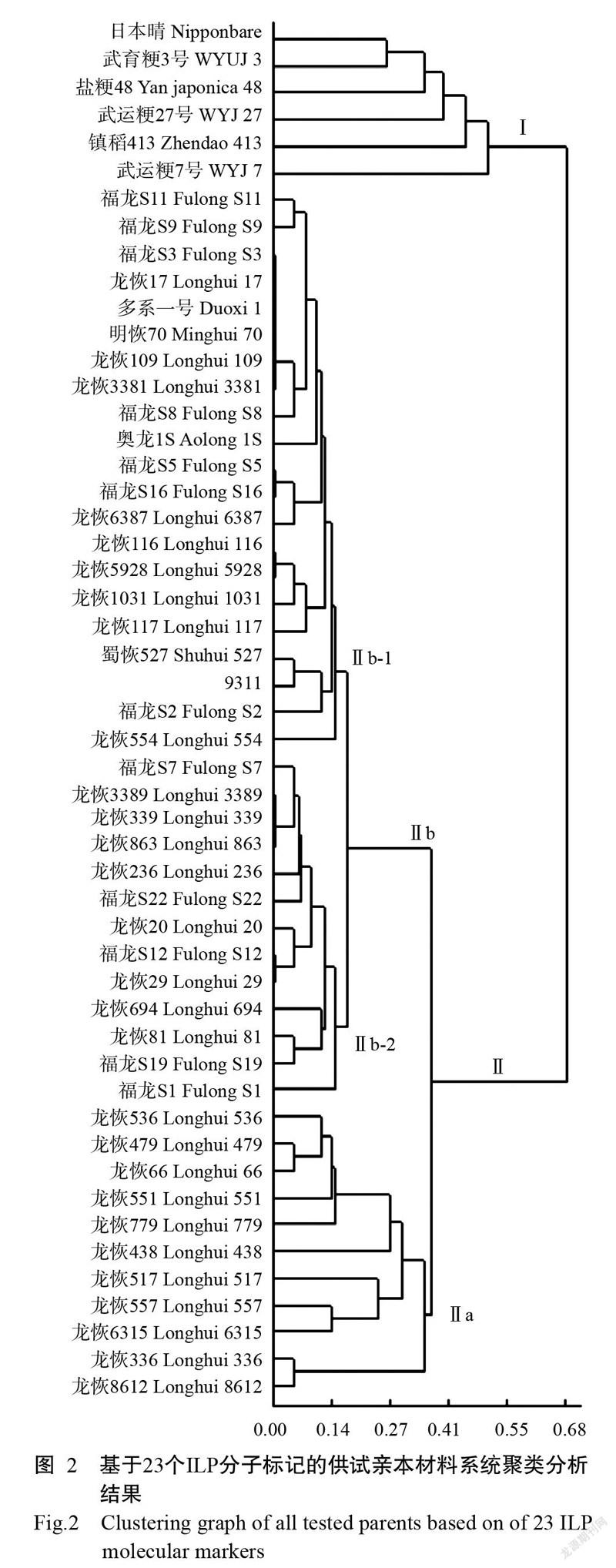
2. 2. 2 ILP分子標記法 由表3可知,51份水稻親本材料基因組DNA在所檢測位點上均存在秈粳分化,不存在100%純秈或純粳類型,共檢測出秈型14份、偏秈型20份、偏粳型11份和粳型6份,平均粳稻成分指數分別為4.35%、11.21%、34.39%和68.70%。
基于23個ILP分子標記的系統聚類分析結果如圖2所示。當閥值為0.67時,51份水稻親本材料可分為Ⅰ和Ⅱ兩個大類,Ⅰ類為粳型(6份),Ⅱ類為其他類型(45份);Ⅱ類在閥值為0.37時可進一步分為Ⅱa和Ⅱb 2個亞類,Ⅱa亞類為偏粳型(11份),Ⅱb亞類為其他類型(34份);Ⅱb亞類在閥值為0.15時又可劃分為Ⅱb-1和Ⅱb-2,分別為秈型(21份)和偏秈型(13份)。
綜上所述,利用ILP分子標記能有效區分供試親本的秈粳屬性,其分類結果和程氏形態指數分類結果均與系統聚類分析結果具有較高的吻合度,是一種有效的水稻秈粳分類工具。
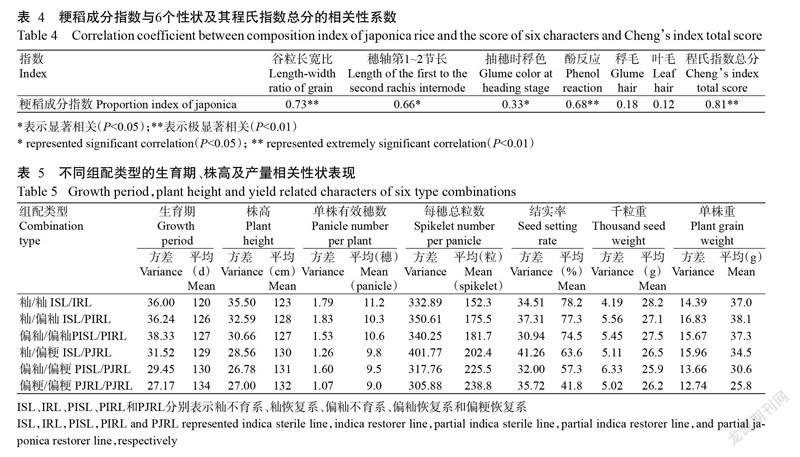
2. 2. 3 ILP分子標記法與程氏形態指數法間的相關性分析 為進一步明晰ILP分子標記法與程氏形態指數法間的相關性,本研究將粳稻成分指數分別與谷粒長寬比、穗軸第1~2節長、抽穗時稃色、酚反應、稃毛、葉毛和程氏指數總分進行相關性分析,結果(表4)顯示,51個供試親本的粳稻成分指數與穗軸第1~2節長和抽穗時稃色呈顯著正相關(P<0.05),與谷粒長寬比、酚反應和程氏指數總分呈極顯著正相關(P<0.01),其中與程氏指數總分的相關系數最高,為0.81,表明ILP分子標記法與程氏形態指數法間具有較高的吻合度,采用ILP分子標記可有效鑒別水稻親本的秈粳類型。
2. 3 親本秈粳分化程度與雜種優勢的相關性
根據ILP分子標記法的秈粳分類結果(表5),按秈/秈、秈/偏秈、偏秈/偏秈、秈/偏粳、偏秈/偏粳和偏粳/偏粳6種類型配制150個雜交組合,其生育期、株高及產量相關性狀如表5所示。隨著組配雙親粳稻成分的增加,雜交組合的生育期、株高和每穗總粒數總體呈增加趨勢,但單株有效穗數、結實率和千粒重總體呈下降趨勢。秈/秈組配類型的生育期、株高和單株有效穗數較理想,較易選配出生育期短、株高較矮且多穗型的組合;秈/偏秈組配類型在各性狀上均有較好的表現,尤其在單株重這一綜合指標上表現最佳,較易選配出強優組合;秈/偏粳組配類型的每穗總粒數較高且方差最大,較易選配出大穗型組合。6種組配類型的單株重排序為秈/偏秈>偏秈/偏秈>秈/秈>秈/偏粳>偏秈/偏粳>偏粳/偏粳,對應的各類型中單株重在40.0 g以上的組合數分別為8、6、5、2、0和0個,隨著組配雙親粳稻成分的增加,單株重呈出先增加后下降的變化趨勢,推測獲得高產組合的概率也呈類似變化趨勢。雖然秈/偏粳組配類型獲得高產組合的概率比秈/秈、秈/偏秈和偏秈/偏秈3種組配類型低,但實際測量結果顯示,150個雜交組合中單株重最重的組合為福龍S16×龍恢336,為46.8 g,表明秈/偏粳組配類型仍有機會獲得極高產組合,育種中亦可大膽嘗試。
3 討論
3. 1 ILP分子標記檢測水稻秈粳分化程度
本研究中ILP分子標記法與程氏形態指數法間的相關性分析結果顯示,51份供試親本材料的粳稻成分指數與穗軸第1~2節長和抽穗時稃色呈顯著正相關,與谷粒長寬比、酚反應和程氏指數總分呈極顯著正相關,其中與程氏指數總分的相關系數最高,為0.81,表明ILP分子標記法與程氏形態指數法間具有較高的吻合度,采用ILP分子標記可有效鑒別供試親本的秈粳類型,與許旭明等(2009)、陳惠清等(2016)的研究結果相同。本研究還與楊旺興等(2013)的研究報道相似,如供試親本外觀形態上屬于秈型的龍恢479、龍恢336、福龍S2和龍恢29,其粳稻成分指數分別為30.43%、34.78%、13.04%和17.39,說明前兩者屬于偏粳型,后兩者屬于偏秈型,表明偏秈、偏粳等中間類型介于秈型與粳型之間,兼具秈稻與粳稻的特征特性,較難區分。這一現象可很好地解釋福龍S2所配組合選育出的品種如福龍兩優29和福龍兩優6387等生產上的強優勢表現,其原因是所配組合較好地利用了部分秈粳雜種優勢。
此外,本研究中水稻品種9311的粳稻成分指數為8.70%,根據許旭明等(2009)的劃分標準,應劃為偏秈型。但據許旭明等(2009)的研究結果顯示,9311粳稻成分指數4.17%,應劃為秈型。兩者存在差異的原因主要有兩個方面:一是使用的ILP分子標記不同導致檢測的粳稻成分指數存在差異;二是劃分標準不夠準確和完善,有待進一步優化。這也是導致本研究中秈粳分類結果和系統聚類分析結果存在差異的原因。
3. 2 親本秈粳分化程度與雜種優勢的關系
本研究發現,隨著組配雙親粳稻成分的增加,秈/秈、偏秈/偏秈、秈/偏秈、偏粳/偏粳、秈/偏粳和偏秈/偏粳6種類型組配雜交組合的生育期、株高和每穗總粒數總體呈增加趨勢,而單株有效穗數、結實率和千粒重總體呈下降趨勢,且這些變化趨勢在秈/秈、秈/偏秈、秈/偏粳組配類型更加明顯。除生育期和結實率外,上述結果與馬彬林(2015)的研究結果基本相同。在結實率方面,隨著秈/秈、秈/偏秈和秈/偏粳組配類型雙親間粳稻成分指數差值的增加,結實率呈明顯下降趨勢,與陳惠清等(2016)的研究結果一致。
關于親本秈粳分化程度與產量雜種優勢間的相關性已有大量研究,但其結果因供試材料不同而各異(黃明,2010;李亞莉等,2012)。本研究發現,當不育系同為秈型時,秈/秈、秈/偏秈和秈/偏粳3種組配類型雙親間的粳稻成分指數差值逐漸增大,雜種優勢先增強后減弱,且對應的各類組合中單株重在40.0 g以上的組合數分別為5、6和8個,當恢復系為偏秈類型時較易選配出強優組合,與龍雯虹等(2003)的觀點相似。另外,本研究結果顯示,雖然采用秈/偏粳方式配組獲得高產組合的概率較低,但有機會獲得極高產組合。通過進一步分析各雜交組合雙親粳稻成分指數發現,在150個雜交組合中,單株重最重的組合為福龍S16×龍恢336,為46.8 g,其父本粳稻成分指數和母本粳稻成分指數的差值為30.43%,福龍S16×龍恢66組合的父本粳稻成分指數和母本粳稻成分指數的差值也為30.43%,但其平均單株重僅為33.6 g,以上兩個組合的雙親粳稻成分差值均未在秈/偏秈組配類型的差值范圍(4.35%~13.04%)內,該現象值得進一步深入研究。
4 結論
利用ILP分子標記可有效區分供試親本的秈粳屬性,是一種有效的水稻秈粳分類工具。采用秈/偏秈方式配組較易篩選到強優組合;雖然采用秈/偏粳方式配組獲得高產組合的概率較低,但有機會獲得極高產組合。
參考文獻:
陳芬,陳雨,曲延英,潘大建,楊慶文,井趙斌,劉維,范芝蘭,陳建酉,李晨. 2009. 粵北普通野生稻秈粳分化的SSR分析[J]. 基因組學與應用生物學,28(2):51-56. [Chen F,Chen Y,Qu Y Y,Pan D J,Yang Q W,Jing Z B,Liu W,Fan Z L,Chen J Y,Li C. 2009. SSR analysis of indica-japonica differentiation of Oryza rufipogon Griff. in Yuebei[J]. Genomics and Applied Biology,(2):51-56.]
陳惠清,黃榮裕,王天生,謝旺有,陳錦文,謝少和,吳建梅,林荔輝. 2016. 水稻秈粳交后代的秈粳分化與雜種F1結實率的相關分析[J]. 福建農業學報,31(3):211-216. [Chen H Q,Huang R Y,Wang T S,Xie W Y,Chen J W,Xie S H,Wu J M,Lin L H. 2016. Correlation between F1 seed-setting and genetic differentiation of indica japonica progenies[J]. Fujian Journal of Agricultural Sciences,31(3):211-216.]
程侃聲. 1993. 亞洲稻秈粳亞種的鑒別[M]. 昆明:云南科技出版社. [Cheng K S. 1993. Indentificating of Subspecies of Indica and Japonica in Oryza sativa L.[M]. Kunming:Science Publishing House of Yunnan Press.]
范方軍,王妍,王芳權,王軍,朱金燕,李文奇,仲維功,胡海燕,楊杰. 2015. 太湖流域水稻地方品種秈粳分化和遺傳多樣性分析[J]. 江蘇農業科學,43(12):68-73. [Fan F J,Wang Y,Wang F Q,Wang J,Zhu J Y,Li W Q,Zhong W G,Hu H Y,Yang J. 2015. Analysis on indica-japonica differentiation and genetic diversity of rice varieties from Taihu lake basin[J]. Jiangsu Agricultural Sciences,43(12):68-73.]
黃明. 2010. 分子標記與水稻遺傳多樣性及雜種優勢的相關性研究[D]. 長沙:湖南農業大學. [Huang M. 2010. Co-rrelation analysis of molecular markers and genetic diversity and heterosis in rice[D]. Changsha:Hunan Agricultural University.]
姜樹坤,鐘鳴,徐正進,張麗,馬慧,劉少霞. 2006. 基于RAPD標記進行水稻秈粳分類研究[J]. 沈陽農業大學學報,37(4):639-644. [Jiang S K,Zhong M,Xu Z J,Zhang L,Ma H,Liu S X. 2006. Classification of rice cultivars with RAPD molecular markers[J]. Journal of Shenyang Agricultural University,37(4):639-644.]
姜廷波,孫傳清,李任華,李自超,王象坤. 1999. 利用RFLP標記對兩系雜交水稻及其親本的分類研究[J]. 中國農業科學,32(6):8-15. [Jiang T B,Sun C Q,Li R H,Li Z C,Wang X K. 1999. Study on classification of crosses and their parents by RFLP markers in two-line system hybrid rice[J]. Scientia Agricultura Sinica,32(6):8-15.]
況浩池,曾正明,劉國民,文紹山,羅俊濤,陳光珍,楊揚. 2007. 水稻耐熱、高配合力秈粳交恢復系瀘恢17的創制與應用[J]. 種子,(10):60-63. [Kuang H C,Zeng Z M,Liu G M,Wen S S,Luo J T,Chen G Z,Yang Y. 2007. Creation and application of the heat tolerance and high combining ability indica japonica rice restorer line Luhui 17[J]. Seed,(10):60-63.]
李亞莉,馬繼武,楊曉曦,許明輝,趙豐萍. 2012. 利用SSR分析滇型雜交稻親本秈粳分化與雜種優勢的關系[J]. 西南農業學報,25(2):347-353. [Li Y L,Ma J W,Yang X X,Xu M H,Zhao F P. 2012. Analysis of relationship between indica-japonica differentiation of parents and hetero-sis in Dian type hybrid rice by SSR markers[J]. Southwest China Journal of Agricultural Sciences,25(2):347-353.]
龍雯虹,許明輝,張樹華. 2003. 滇型雜交稻親本秈粳分化與雜種優勢關系的初步研究[J]. 中國農業科學,36(1):1-6. [Long W H,Xu M H,Zhang S H. 2003. A preliminary study on the relationship between the indica-japonica differentiation of parents and heterosis in dian type hybrid rice by RAPD markers[J]. Scientia Agricultura Sinica,36(1):1-6.]
馬彬林. 2015. 秈粳衍生恢復系的研制與利用[J]. 福建農業學報,30(4):322-325. [Ma B L. 2015. Development and utilization of the restorer lines derived from indica-japonica cross[J]. Fujian Journal of Agricultural Sciences,30(4):322-325.]
孫傳清,姜廷波,陳亮,吳長明,李自超,王象坤. 2000. 水稻雜種優勢與遺傳分化關系的研究[J]. 作物學報,26(6):641-649. [Sun C Q,Jiang T B,Chen L,Wu C M,Li Z C,Wang X K. 2000. Studies on the relationship between heterosis and genetic differentiation in hybrid rice(Oryza sativa L.)[J]. Acta Agronomica Sinica,26(6):641-649.]
王明軍,王云月,陸春明,楊慧,王云濤. 2010. 利用秈粳稻特異InDel標記分析云南糯稻品種的秈粳特性[J]. 云南農業大學學報(自然科學版),25(3):333-337. [Wang M J,Wang Y Y,Lu C M,Yang H,Wang Y T. 2010. The indica-japonica characteristics of yunnan glutinous rice varieties were analyzed by indica-japonica InDel markers[J]. Journal of Yunnan Agricultural University(Natural Science),25(3):333-337.]
徐海,劉宏光,楊莉,朱春杰,王嘉宇,楊乾華,徐正進,鄭家奎. 2007. 不同生態條件下秈粳稻雜交后代亞種特性的比較研究[J]. 作物學報,33(3):370-377. [Xu H,Liu H G,Yang L,Zhu C J,Wang J Y,Yang Q H,Xu Z J,Zheng J K. 2007. Subspecies characteristics in filial gene-ration of cross between indica and japonica under different environments[J]. Acta Agronomica Sinica,33(3):370-377.]
許旭明,梁康逕,張受剛,尚偉,張瑛英,韋新宇,柯蓓. 2009. 利用ILP標記分析水稻秈粳雜交親本和衍生系的秈粳分化[J]. 中國農業科學,42(10):3388-3396. [Xu X M,Liang K J,Zhang S G,Shang W,Zhang Y Y,Wei X Y,Ke B. 2009. Analysis of indica-japonica differentiation in rice parents and derived lines using ILP markers[J]. Scientia Agricultura Sinica,42(10):3388-3396.]
楊旺興,許旭明,祁建民,卓偉,張受剛,馬彬林,韋新宇,鄒文廣. 2013. 水稻秈粳交衍生系秈粳分化分析及其在DNA水平上劃分標準初探[J]. 西南農業學報,26(4):1301-1307. [Yang W X,Xu X M,Qi J M,Zhuo W,Zhang S G,Ma B L,Wei X Y,Zou W G. 2013. Analysis of indica-japonica classification in rice derived lines and division standard in DNA level[J]. Southwest China Journal of Agricultural Sciences,26(4):1301-1307.]
袁隆平,武小金,顏應成,羅孝和. 1997. 水稻廣譜廣親和系的選育策略[J]. 中國農業科學,30(4):1-8. [Yuan L P,Wu X J,Yan Y C,Luo X H. 1997. A strategy for developing wide spectrum compatibility rice line[J]. Scientia Agricultura Sinica,30(4):1-8.]
張建勇,袁佐清,李仕貴. 2005. 微衛星標記分析秈粳亞種間的遺傳多樣性[J]. 山東理工大學學報,9(2):22-27. [Zhang J Y,Yuan Z Q,Li S G. 2005. The genetic diversity and relationship between indica and japonica rice(Oryza sativa L.) using SSR markers[J]. ?Journal of Shandong University of Technology,9(2):22-27.]
張受剛,許旭明. 2006. 粳型親秈恢復系的選育與利用[J]. 福建稻麥科技,24(1):3-5. [Zhang S G,Xu X M. 2006. Breeding and utilization indica compatible restorer lines[J]. Fujian Science and Technology of Rice and Wheat,24(1):3-5.]
鄭秀娟,解慧芳,曹紅瑞,楊桂英,謝長榮,梁康逕,孫新立. 2016. 水稻秈粳亞種分化關聯性狀的QTL分析[J]. 福建農林大學學報(自然科學版),45(1):1-7. [Zheng X J,Xie H F,Cao H R,Yang G Y,Xie C R,Liang K J,Sun X L. 2016. QTL analysis of the differentiation characteri-stics of indica and japonica rice and associated traits[J]. Journal of Fujian Agriculture and Forestry University(Natural Science Edition),45(1):1-7.]
左生力. 2016. 幾個秈型兩系雜交稻親本的配合力及遺傳力分析[J]. 福建農業學報,31(12):1299-1307. [Zuo S L. 2016. Analysis on combining ability and heritability of 5 new PTGMS lines and 6 new restorer lines in Indica hybrid rice[J]. Fujian Journal of Agricultural Sciences,31(12):1299-1307.]
Li R H,Jiang T B,Xu C G,Li X H,Wang X K. 2000. Relationship between morphological and genetic differentiation in rice(Oryza sativa L.)[J]. Euphytica,114(1):1-8.
(責任編輯 陳 燕)
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
