水稻棉絮狀花序突變體ecp1的鑒定與基因定位
2019-09-10 07:22:44郭新睿楊紹華劉華清李剛
福建農業科技 2019年9期
關鍵詞:水稻
郭新睿 楊紹華 劉華清 李剛



摘 要:通過對水稻秈型恢復系明恢86組培變異后代的篩選,得到1個棉絮狀花序突變體ecp1(encapsulated cotton panicle 1)。棉絮狀花序突變體表現生育期內水稻抽穗異常,其小穗部被白色類棉絮物質囊狀包裹,無法形成正常的稻穗,進而導致無法結實。遺傳分析表明,ecp1受單一隱性核基因控制。利用秈稻品種93-11與ecp1突變體雜交的F2群體,通過SSR和InDel標記,將ecp1定位于水稻第8染色體約240 kb的區間。研究結果為ecp1基因的克隆和功能分析奠定了基礎。
關鍵詞:水稻;棉絮狀花序;突變體;鑒定;基因定位
中圖分類號:S511 文獻標志碼:A 文章編號:0253-2301(2019)09-002
Abstract: A encapsulated cotton panicle mutant (ecp1) was obtained by screening the tissue culture and variant progenies of restorer line Minghui 86 of indica rice. The encapsulated cotton panicle mutant showed abnormal rice heading during the growth period, and its spikelet was enveloped by white cottonlike material, which could not form normal rice spike, and thus could not seed. The genetic analysis showed that ecp1 was controlled by a single recessive nuclear gene. F2 colony from a hybrid of indica rice variety 93-11 and ecp1 mutant was used to locate ecp1 in the region of about 240 kb on chromosome 8 of rice by SSR and InDel markers. The results of this study laid a foundation for the cloning and functional analysis of ecp1 gene.
Key words: Rice; Encapsulated cotton panicle; Mutant; Identification; Gene localization
水稻被廣泛栽培,是世界范圍內最重要的糧食作物之一。其產量不但同光照、溫度、水分等環境因素相關,而且同自身分蘗、粒型、穗型等息息相關。鑒于水稻的小穗著生于側生分枝上,而不是著生在主穗軸上,故其花序稱為總狀花序。又因為水稻花序形似圓錐,所以又被稱為圓錐花序[1]。水稻花序是否可以正常發育影響著育種學家特別關心的產量問題。因此,對水稻的花序以及花器官發育研究一直以來都是水稻分子遺傳學研究的熱點問題之一。
至今,一些調控水稻花序發育的相關基因或數量性狀位點(QTL)陸續被克隆。其中,短穗基因SP1(short panicle1) 調控水稻枝梗分生組織的伸長。此突變體枝梗原基的發育起始正常,但在枝梗延伸時出現嚴重障礙,因而導致短穗[2-3]。DEP2(dense and erect panicle2) 基因則編碼了1個植物特有的未知蛋白,其主要影響一、二級枝梗和穗軸的伸長[2-4]。OsAPO1/SCM2(aberrant panicle organization 1) 基因造成枝梗分生組織退化時間的延長,正調節小穗數和一級枝梗的數目[2-6]。IPA1(ideal plant architecture) 、TAW1(tawawa 1) 和OsASP1(aberrant spikelet and panicle 1) 等基因則是在水稻花序發育過程中,目前已發現的影響枝梗分生組織向小穗分生組織轉換的主要基因[7-9]。上述一系列花分生組織決定基因的發現為理解植物花的形成發育機制提供了分子基礎。目前為止,雖然已發現和克隆了部分花序相關基因,但是對花在花序中著生位置和花序發育的遺傳調控機制并不清楚,調控花序發育的相關基因有待進一步發現和研究。因此繼續對植物花序形態突變體的研究很有必要。
本實驗室獲得了1例水稻棉絮狀花序突變體,對該突變體進行遺傳分析,結果表明突變體ecp1定位于水稻第8染色體約240 kb的區間。本研究對水稻的棉絮狀花序突變體進行了形態特征和性狀觀察、遺傳分析以及基因初步定位,為克隆和研究該基因打下了基礎。
1 材料與方法
1.1 研究材料
ecp1突變體經秈稻恢復系明恢86組織培養的后代中篩選得到;其他供試材料:ecp1的原始野生型品種明恢 86、 秈稻品種93-11及粳稻品種02428,均由福建省農業遺傳工程重點實驗室保留和提供。
1.2 表型鑒定
研究材料ecp1突變體、秈稻品種93-11、野生型明恢86以及粳稻品種02428均種植于福建省農業科學院生物技術研究所福州壽山基地,在整個生長發育期內,對野生型和突變體材料的表型進行觀察并記載。
1.3 群體構建和遺傳分析
分別以野生型明恢 86、秈稻品種93-11和粳稻品種02428為母本,ecp1突變體為父本,配制F2分離群體。于抽穗期分別調查和統計F2群體中正常株與突變株的數量,繼而計算出正常株和突變株的分離比,然后進行卡平方測驗。
1.4 分子標記試驗
根據Gramene 網站(http://www.gramene.org/)提供的RM系列水稻的SSR標記進行連鎖分析和初定位。繼而利用Grame網站的SSRIT軟件掃描粳稻品種Nipponbare(http://rapdb.dna.affrc.go.jp/download/)的基因組序列,在目標基因所在區域內查找出新的SSR位點,并通過BLAST對Nipponbare和秈稻品種93-11(http://rice.genomics.org.cn/rice/link/download.jsp) 的基因組序列進行比對,尋找插入缺失(InDel)位點。最后利用Web-Premier (http://www.yeastgenome.org/cgibin/webprimer)軟件,在SSR和InDel 的兩側設計特異引物來開發新的SSR和InDel標記。
1.5 ecp1基因定位
對由水稻ecp1突變體和秈稻品種93-11配制出的F2群體進行基因定位。運用BSA(Bulked segregant analysis)原理,從F2群體中隨機抽樣,選取出正常植株和突變植株各10株,通過CTAB法提取出總DNA分別構建正常DNA池和ecp1突變DNA池。PCR反應體系及PCR產物檢測參照林艷等[10]的方法。以兩親本為對照,根據本實驗室已經篩選出來的多態性分子標記,選取水稻的12條染色體上均勻分布的480對分子標記,快速尋找到其中與目標基因連鎖的部分標記。進一步選取F2群體中的20個隨機水稻棉絮狀花序突變體單株,對其進行驗證,確定基因所在染色體。最后利用該標記所在區域的多態性,新開發出InDel標記并分析F3分離群體的180個單株,對目標基因進行了定位。
2 結果與分析
2.1 ecp1突變體的形態特征和性狀分析
自然條件下,ecp1突變體表現為生長勢弱,分蘗數少,穗不能正常發育,呈棉絮狀。在苗期營養生長正常,表型和野生型(明恢86)相似,分蘗期表現為少分蘗。抽穗期,突變體穗不能正常抽出,不能形成正常枝梗及小穗,類似棉絮(圖1)。因此,將該突變體命名為棉絮狀花序突變體encapsulated cotton panicle 1(ecp1)。
2.2 ecp1突變體的遺傳分析
ecp1突變體與其野生型明恢86、秈稻品種9311和粳稻品種02428進行雜交,F1代植株均發育正常,可以正常抽穗結實。F1自交獲得的F2,經調查統計,ecp1/明恢86、ecp1/9311和ecp1/02488的F2代均出現正常植株與棉絮狀花序突變植株的分離,分離比例分別為416∶120、423∶138和730∶233,卡平方分析表明3個不同群體分離比均符合3∶1的分離比(表1),說明該突變性狀是由單隱性核基因控制。并且因為在不同的遺傳背景下,突變體均能表現出明顯的棉絮狀花序表型,表明該突變性狀能夠穩定遺傳。
2.3 ecp1突變體的基因定位
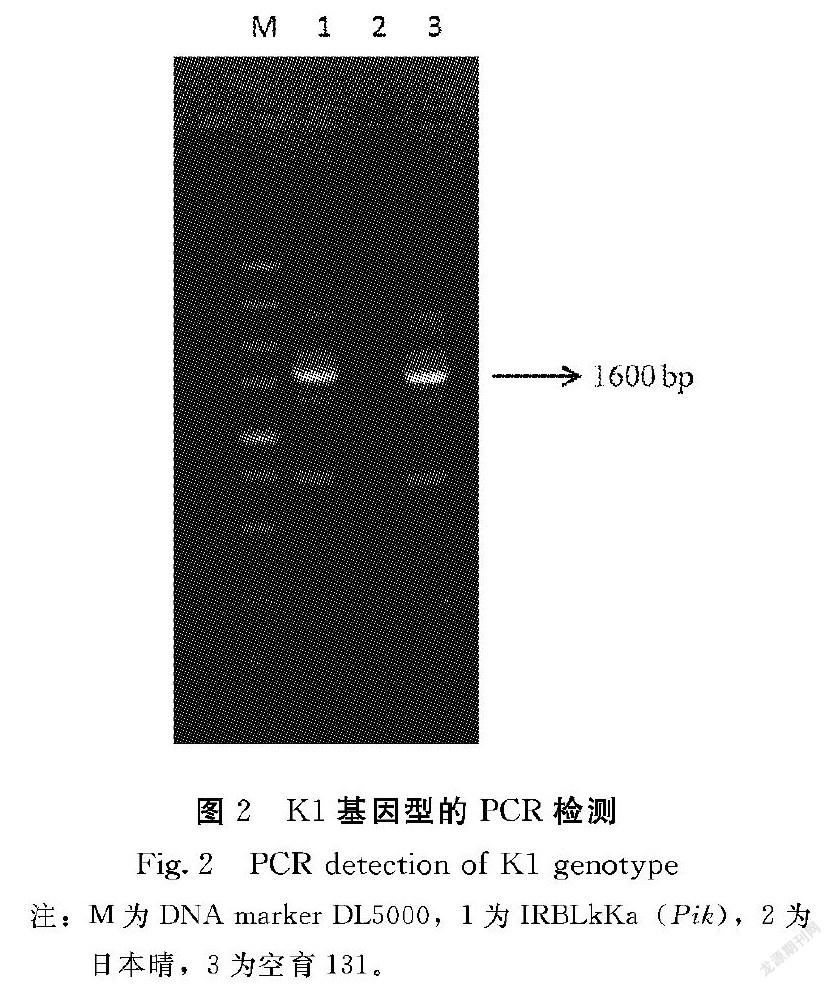
利用在兩親本間有多態性,分布于水稻12條染色體的480對SSR引物對兩個近等基因池進行篩選,發現位于第8染色體相鄰的3對SSR引物(RM1376、RM3572和RM3215)對兩基因池擴增出具有明顯多態性的DNA片段(圖2),說明它們可能與目標基因ecp1連鎖。進一步用這3個標記對138個F2突變株進行分析。結果表明,ecp1基因位于RM1376和RM3572之間,與RM1376遺傳距離為3.1 cM,與RM3572的遺傳距離則為3.3 cM。
將分離群體進一步擴大到180個隱性單株,參考明恢63和9311的基因組序列,根據分子標記RM3572在9311基因組序列中對應的物理位置,在其上游附近開發了19對大小在80~150 bp的 InDel標記。其中InDel0451、InDel0581和InDel5143這3對引物在群體中擴增出多態性條帶(表2),利用RM3572和這3對InDel標記引物對上述交換單株進行擴增,最終將突變基因定位在InDel0451和InDel0581之間,與InDel5143共分離,突變基因與兩標記InDel0451和InDel0581之間的物理距離分別為32 kb與108 kb(圖3)。
3 討論與結論
植物的花序是一個比營養器官更為復雜的存在,是在植物花的發育過程中,營養生長向生殖生長轉變所形成的結構。近些年來,隨著分子生物學的發展,多個與植物花發育相關的基因以擬南芥和金魚草為模式植物被發現和獲得,其中一系列花分生組織決定基因的發現尤為重要,為解釋植物花的形成發育機制提供了分子基礎[11]。對水稻花序突變體及其調控基因進行研究,有助于解析水稻花序發育的調控機理,對水稻高產分子設計育種研究具有重要的指導意義。
本研究結果發現,ecp1突變體出現了穗長變短,花序被白色類棉絮狀葉鞘包裹,無法識別正常小穗結構,單穗無法結實等突變表型。遺傳分析顯示,ecp1受1對隱性單基因控制,并將其定位在水稻第8染色體上前段約240 kb 區間內,與已報道的水稻花序相關突變基因asp1的定位區間相近。Asp1 突變體枝梗發育無序,花序排列不規則,小穗畸形[9]。本研究ecp1突變體與其表型相似,但呈現出特有的棉絮狀包裹花序的現象,導致完全無法結實。至于ecp1突變體與asp1突變體是否是同一基因的突變位點不同導致的表型不同,還是因突變體遺傳背景不同所致,還需要進一步對ecp1突變體進行序列測定和遺傳共分離分析。同時,對ecp1突變體與其他突變體的遺傳背景差異進行研究,將有助于深入了解ecp1基因調控水稻花序形成及發育的調控機制。
參考文獻:
[2]高繼平,祁 澎,林鴻宣.水稻產量數量性狀的遺傳調控機制研究進展[J].中國科學:生命科學,2013,43(12):1007-1015.
[3]汪 慶,汪得凱,陶躍之.一個新的水稻半矮化小穗突變體的遺傳分析與基因定位[J].中國水稻科學,2011,25(6):677-680.
[4]盧 寰,時振英.水稻穗發育的分子生物學研究進展[J].植物生理學報,2013,49(2):111-121.
[5]DOI K,IZAWA T,FUSE T,et al.Ehd1,a Btype response regulator in rice,confers shortday promotion of flowering and controls FTlike gene expression independently of Hd1[J].Genes & Development,2004,18(8):926-936.
[6]KOJIMA S,TAKAHASHI Y,KOBAYASHI Y,et al.Hd3a,a rice ortholog of the Arabidopsis FT gene,promotes transition to flowering downstream of Hd1 under shortday conditions[J].Plant & Cell Physiology,2002,43(10):1096-1105.
[7]JIAO Y Q,WANG Y H,XUE D W,et al.Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice[J].Nature Genetics,2010,42(6):541-544.
[8]LU Z F,YU H,XIONG G S,et al.Genomewide binding analysis of the transcription activator ideal plant architecture1 reveals a complex network regulating rice plant architecture[J].The Plant Cell,2013,25(10):3743-3759.
[9]YOSHIDA A,OHMORI Y,KITANO H,et al.ANERRANT SPIKELET and PANICLE1,encoding a TOPLESS related transcriptional corepressor,is involved in the regulation of meristem fate in rice[J].The Plant Journal:Cell and Molecular Biology,2012,70(2):327-339.
[10]林艷,陳在杰,田大剛,等.水稻類病斑及早衰突變體lms1的鑒定及基因初步定位[J].福建農業學報, 2014,29(1):29-34.
[11]盧海敏,高廷旺,單世華,等.植物開花相關基因研究進展[J].安徽農業科學,2008,36(5):1803-1805.
(責任編輯:柯文輝)
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
