BmNPV對家蠶抗氧化酶基因表達及其酶活性的影響
2019-09-10 06:14:59唐芬芬楊偉克朱峰邵榆嵐張永紅白興榮
南方農業學報 2019年10期
唐芬芬 楊偉克 朱峰 邵榆嵐 張永紅 白興榮

摘要:【目的】探究喂食家蠶核型多角體病毒(BmNPV)后家蠶血淋巴和中腸組織抗氧化酶[超氧化物歧化酶(SOD)和過氧化氫酶(CAT)]的活性及其基因表達變化規律,明確BmNPV侵染對家蠶抗氧化系統的影響,為解析BmNPV的致病機理提供理論依據。【方法】以五齡家蠶為研究對象,經口喂食BmNPV,分別于添食后6、12、18、24、30、36、42和48 h采集家蠶的血淋巴和中腸組織,采用實時熒光定量PCR檢測兩種抗氧化酶基因(Bmsod和Bmcat)的表達水平,同時以抗氧化酶活性測試盒測定SOD和CAT的活性變化情況。【結果】BmNPV侵染五齡家蠶能引起典型的血液型膿病,主要表現為蠶體環節腫脹,狂躁爬行,體壁易破,體液呈乳白色等。家蠶血淋巴和中腸組織中Bmsod和Bmcat基因均在感染BmNPV中期開始上調表達,在感染后期呈下調表達趨勢,其相對表達量也呈先增加后急劇減少的變化趨勢。添食BmNPV后,家蠶血淋巴和中腸組織的SOD活性在感染18和24 h時極顯著高于對照組(添食等量滅菌水)家蠶(P<0.01,下同),但從感染30 h起SOD活性開始急劇下降,至感染48 h時降至最低值。家蠶血淋巴和中腸組織的CAT活性在感染早期(6 h)略有下降,從感染12 h起CAT活性開始呈上升趨勢,血淋巴CAT活性在感染18~30 h極顯著高于對照組,隨后急劇下降且顯著低于對照組家蠶(P<0.05,下同);中腸組織CAT活性僅在感染18和24 h時顯著或極顯著高于對照組家蠶,從感染30 h起開始持續下降,至感染48 h時降至最低值,約為對照組家蠶的18%。【結論】BmNPV侵染能影響家蠶機體相關保護酶活性及其基因轉錄水平,SOD和CAT活性及其基因表達量在感染中期增加,感染后期則急劇降低,提示抗氧化酶SOD和CAT在家蠶的抗病毒過程中發揮重要作用。
關鍵詞: 家蠶;家蠶核型多角體病毒(BmNPV);SOD;CAT;酶活性;基因表達
中圖分類號: S884.51? ? ? ? ? ? ? ? ? ? ?文獻標志碼: A 文章編號:2095-1191(2019)10-2308-06
Effects of BmNPV on the antioxidant enzyme gene expression and enzyme activity of Bombyx mori
TANG Fen-fen, YANG Wei-ke, ZHU Feng, SHAO Yu-lan,
ZHANG Yong-hong, BAI Xing-rong*
(Institute of Sericulture and Apiculture, Yunnan Academy of Agricultural Sciences, Mengzi, Yunnan? 661101, China)
Abstract:【Objective】In order to provide references for further study on the infection and pathogenesis of BmNPV, this study was explored change rules of the hemolymph and mesenteron antioxidant enzyme activities[superoxide dismutase(SOD), catalase(CAT)] and the gene expression in silkworm larvae infected with Bombyx mori nuclear polyhedrovirus(BmNPV), and the effects of BmNPV infection on B. mori antioxidant system were also analyzed. 【Method】The real-time fluorescent quantitative PCR was applied to analyze the expression of two kinds of? antioxidant enzyme genes Bmsod and Bmcat, meanwhile the activity of the SOD and CAT were determined in heamolymph and midgut of 5th instar silkworm larvae after feeding BmNPV at 6, 12, 18, 24, 30, 36, 42 and 48 h. 【Result】BmNPV infection of silkworm in the fifth instar could cause typical blood pyogenic disease, mainly manifested as swollen silkworm body parts, manic crawling, body wall easily broken, body fluid milky liquid. The Bmsod and Bmcat genes were up-regulated since the mi-ddle of BmNPV infection, and down-regulated in the end of infection, their relative expression also showed increase-sharp decrease trend. The activities of SOD in hemolymph and midgut of B. mori was extremely higher than the control group (feeding the same amount of sterilized water) at 18 and 24 h after infection with BmNPV(P<0.01, the same below), but it started to decrease sharply at 30 h, and dropped to the lowest value at 48 h. The activities of CAT in hemolymph and midgut of silkworm decreased slightly at the early stage after infection(at 6 h), and increased from 12 h. In hemolymph, the CAT activity was extremely higher than the control group from 18 h to 30 h after infection, but decreased sharply and was significantly lower than the control group(P<0.05, the same below). In midgut, the CAT activity was only significantly or extremely significantly higher than the control group at 18 and 24 h after infection, and continued to decline from 30 h to the lowest value at 48 h, which was about 18% of the control group. 【Conclusion】BmNPV infection can affect the activity of related protective enzymes and its gene transcription level in silkworm.The expression levels of activities of SOD and CAT and Bmsod and Bmcat genes all increase first and then decrease sharply in the midgut and heamolymph of silkworm after infection with BmNPV. It suggests that the antioxidant enzymes SOD and CAT play an important role in the antiviral process of silkworm.
Key words: Bombyx mori; Bombyx mori nuclear polyhedrovirus(BmNPV); SOD; CAT; enzyme activity; gene expression
0 引言
【研究意義】家蠶是一種重要的經濟昆蟲,同時是鱗翅目昆蟲的模式生物,加之遺傳背景清晰且飼育簡單,已發展成為研究宿主—病原互作機制的理想材料(Geng et al.,2016a,2016b)。超氧化物歧化酶(Superoxide dismutase,SOD)和過氧化氫酶(Catalase,CAT)在維持生物體內自由基代謝平衡的過程中發揮重要作用,在一定程度上能反映生物的抗逆能力(Lesser,2006;方允中和鄭榮梁,2008)。因此,研究抗氧化酶活性及其基因表達在家蠶抵抗病原微生物侵染過程中的變化規律,尤其是明確其在家蠶抵抗家蠶核型多角體病毒(Bombyx mori nucleopolyhedrovirus,BmNPV)時的功能和作用,可為揭示BmNPV侵染及其致病機理提供理論依據。【前人研究進展】BmNPV感染主要引發家蠶血液型膿病,傳染性強,危害嚴重,每年給桑蠶產業帶來巨大經濟損失(邵榆嵐等,2017)。據家蠶流行病學調查發現,BmNPV是云南蠶區發生較廣且危害嚴重的主要病原(白興榮等,2010),生產上多在三齡蠶后感染發病,且一旦發病即迅速蔓延。雖然關于家蠶抵抗病毒的研究已有大量報道,但抗病機制至今尚未完全明確。昆蟲遭受外源病原菌侵染時,其機體內正常的代謝平衡被打破,誘發活性氧(ROS)過度積累,進而損傷宿主細胞(Kumar et al.,2003)。抗氧化酶CAT和SOD在消除昆蟲體內過量氧自由基方面發揮著重要作用,其中,CAT能催化H2O2分解為H2O和O2,減少羥自由基的產生,防止過量羥自由基損傷機體(Zhao and Shi,2009);SOD能催化超氧陰離子自由基([O2] )發生歧化反應,生成H2O2和O2,H2O2再經CAT作用轉化為H2O和O2,進而維持機體的正常生理代謝過程(Wang et al.,2001)。李世廣等(2016)研究發現,菜青蟲感染金龜子綠僵菌后其體內SOD和CAT活性呈先上升后下降的變化趨勢,表明蟲體在侵染初期防御能力增強,但后期防御能力降低。張仙紅等(2006)研究證實,菜青蟲感染玫煙色擬青霉后體內SOD和CAT活性迅速提高,隨后急劇下降并顯著低于未感染組,同時出現感病癥狀,說明昆蟲SOD和CAT等保護酶系統與病原菌對宿主的侵染存在一定關聯,但具體作用機制尚未明確。王樹昌等(2017)研究發現,家蠶感染球孢白僵菌后其血淋巴、中腸等組織的總抗氧化活力、SOD和CAT活性均顯著降低,Bmsod和Bmcat基因表達水平顯著下調,而添加外源SOD和CAT等抗氧化劑能有效延長家蠶存活時間,故推測球孢白僵菌的致病機制主要是通過破壞家蠶抗氧化系統平衡,損傷組織器官,降低宿主的免疫防御功能,最終導致蟲體發病死亡。【本研究切入點】家蠶抵抗BmNPV涉及的防御反應極其復雜,BmNPV侵染不僅誘導家蠶中腸相關抗菌肽基因的表達,還能上調血淋巴中酚氧化酶原基因表達,進而提高酚氧化酶活性(唐芬芬等,2015,2016)。目前,有關家蠶對BmNPV的免疫防御及免疫相關因子研究已較深入,但遭受BmNPV侵染時蠶體抗氧化酶活性及其基因表達變化規律的研究鮮見報道。【擬解決的關鍵問題】以五齡家蠶為研究對象,探究添食BmNPV后家蠶血淋巴和中腸組織抗氧化酶SOD和CAT的活性及其基因表達變化規律,明確BmNPV侵染對家蠶抗氧化系統的影響,為解析BmNPV的致病機理提供理論依據。
1 材料與方法
1. 1 試驗材料
家蠶品系菁松×皓月和BmNPV均由云南省農業科學院蠶桑蜜蜂研究所家蠶病害防控研究室提供。Total RNA提取試劑盒(OMEGA公司,貨號R6834-02)、反轉錄試劑盒PrimeScript RT Reagent Kit with gDNA Eraser(TaKaRa公司,貨號RR047A)、FastStart SYBR Green Master(Roche公司,貨號04673484001)、SOD活性測試盒(貨號A001-1)和CAT活性測試盒(貨號A007-1)均購自南京建成生物工程研究所。
1. 2 添食處理及樣品采集
家蠶按常規方法飼養至五齡起蠶,取體型和體重相近的家蠶隨機分為兩組(試驗組和對照組),試驗組每頭家蠶經口添食10 μL預先制備好的病毒懸液(5.35×108個/mL),對照組添食等量滅菌水,添食后在同等條件下進行飼育。于添食后6、12、18、24、30、36、42和48 h采集家蠶的血淋巴和中腸組織,每次取樣均分成兩份,一份用于抽提RNA,另一份用于制備酶液。每次取樣設4個重復,每個重復5頭家蠶,樣品收集后-80.0 ℃保存備用。
1. 3 總RNA提取及反轉錄
按照Total RNA提取試劑盒操作說明提取各組織RNA,以紫外分光光度計測定RNA樣品濃度,取OD260/OD280為1.80~2.00的樣品,根據反轉錄試劑盒說明將RNA反轉錄合成cDNA。
1. 4 實時熒光定量PCR檢測
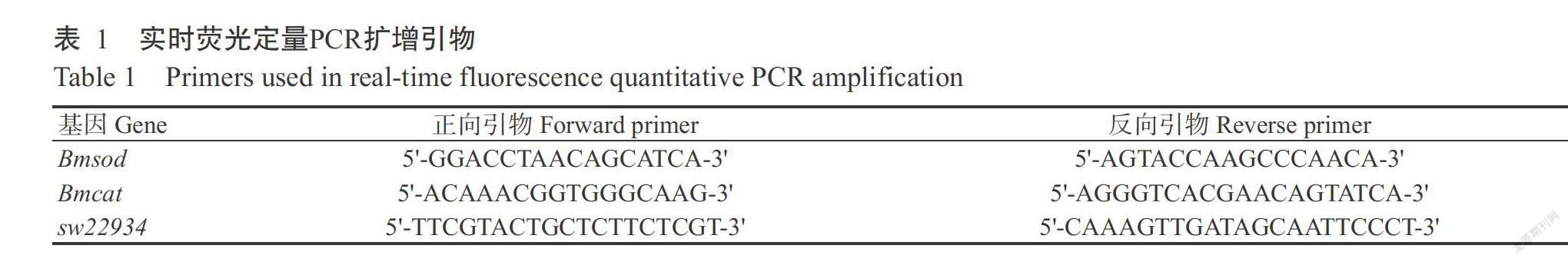
以真核翻譯起始因子4A基因(家蠶芯片探針ID:sw22934)為內參基因、酚氧化酶原基因Bmsod(NCBI登錄號NM_001043619.1)和Bmcat(NCBI登錄號NM_001043447.1)為靶標基因。利用Primer Premier 5.0設計引物(表1)。參照FastStart SYBR Green Master試劑盒操作說明進行實時熒光定量PCR檢測,擴增程序:95.0 ℃預變性30 s;95.0 ℃ 5 s,60.0 ℃ 30 s,進行40個循環;65.0 ℃ 5 s(每次加0.5 ℃,直至95.0 ℃)。以CFX-96實時熒光定量PCR檢測儀記錄試驗數據,根據2-△△Ct法計算目的基因相對表達量(Livak and Schmittgen,2001)。
1. 5 抗氧化酶活性測定
酶液制備:取家蠶幼蟲各組織樣品,分別加入1.2 mL預冷的磷酸鹽緩沖液(0.1 mol/L,pH 7.0),在冰上進行研磨,使樣品充分破碎,于4 ℃下2500 r/min離心15 min,收集上清液,即為待測酶液。按照SOD活性測試盒和CAT活性測試盒使用說明,分別測定SOD和CAT的活性。
1. 6 統計分析
采用Excel 2016進行數據處理及作圖,同時利用SPSS 19.0進行統計分析。
2 結果與分析
2. 1 家蠶感染BmNPV后的主要臨床癥狀
BmNPV侵染五齡家蠶能引起典型的血液型膿病,從感染后第4 d起開始發病,其臨床癥狀主要表現為蠶體環節腫脹,狂躁爬行,體壁易破,體液呈乳白色等(圖1)。
2. 2 家蠶感染BmNPV后Bmsod和Bmcat基因的表達變化
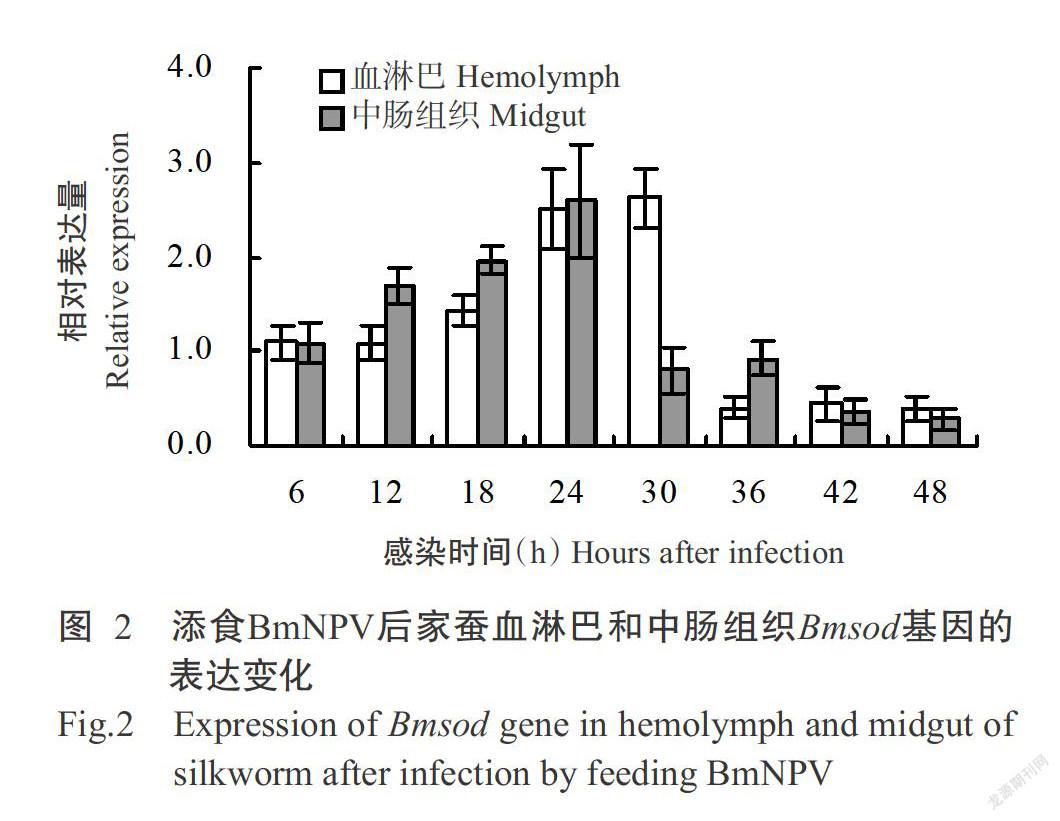
采用實時熒光定量PCR測定添食BmNPV后家蠶血淋巴和中腸組織中Bmsod和Bmcat基因的相對表達量,將對照組家蠶目的基因的相對表達量設定為1.0,當試驗組家蠶目的基因的相對表達量大于1.0時,表明該基因上調表達,反之則下調表達。由圖2可看出,家蠶血淋巴Bmsod基因在感染BmNPV前期(6~12 h)的相對表達量接近于1.0,至感染18 h時開始上調表達,其相對表達量逐漸增加,于感染30 h時達最高值,隨后家蠶血淋巴Bmsod基因相對表達量急劇下降,呈下調表達趨勢。在家蠶中腸組織中,Bmsod基因在感染BmNPV后12~24 h呈持續上調表達趨勢,且在感染24 h時達最高值,至感染后期(30~48 h)Bmsod基因呈下調表達。
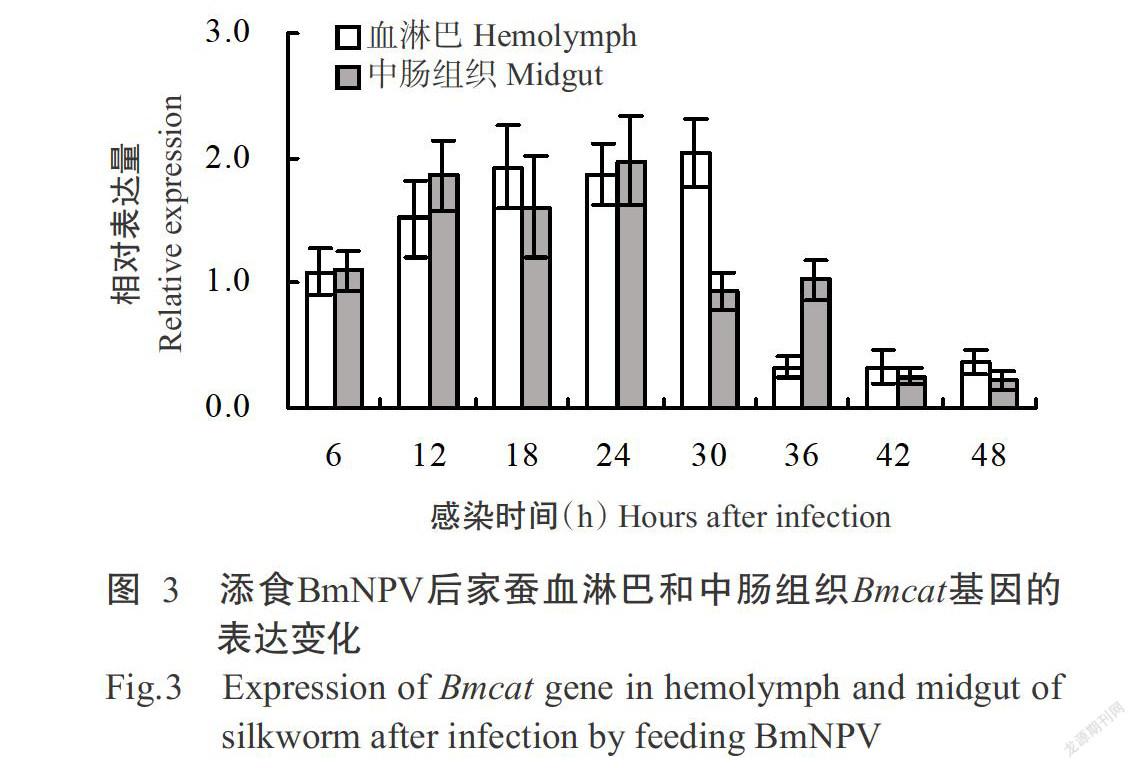
由圖3可看出,在BmNPV侵染前6 h,家蠶血淋巴和中腸組織中的Bmcat基因相對表達量均無明顯變化,但從感染12 h起開始上調表達,其中,血淋巴Bmcat基因約上調1.5倍,中腸組織Bmcat基因約上調1.8倍。家蠶血淋巴Bmcat基因在感染BmNPV后12~30 h均呈上調表達趨勢,在感染36~48 h呈持續下調表達趨勢;中腸組織Bmcat基因在感染12~24 h呈上調表達,從感染30 h起開始下調表達,至感染42和48 h其相對表達量降至最低值。
綜上所述,家蠶血淋巴和中腸組織中的Bmsod和Bmcat基因均在感染BmNPV中期開始上調表達,至感染后期則下調表達,其相對表達量也呈先增加后急劇減少的變化趨勢。
2. 3 家蠶感染BmNPV后血淋巴和中腸組織SOD活性的變化
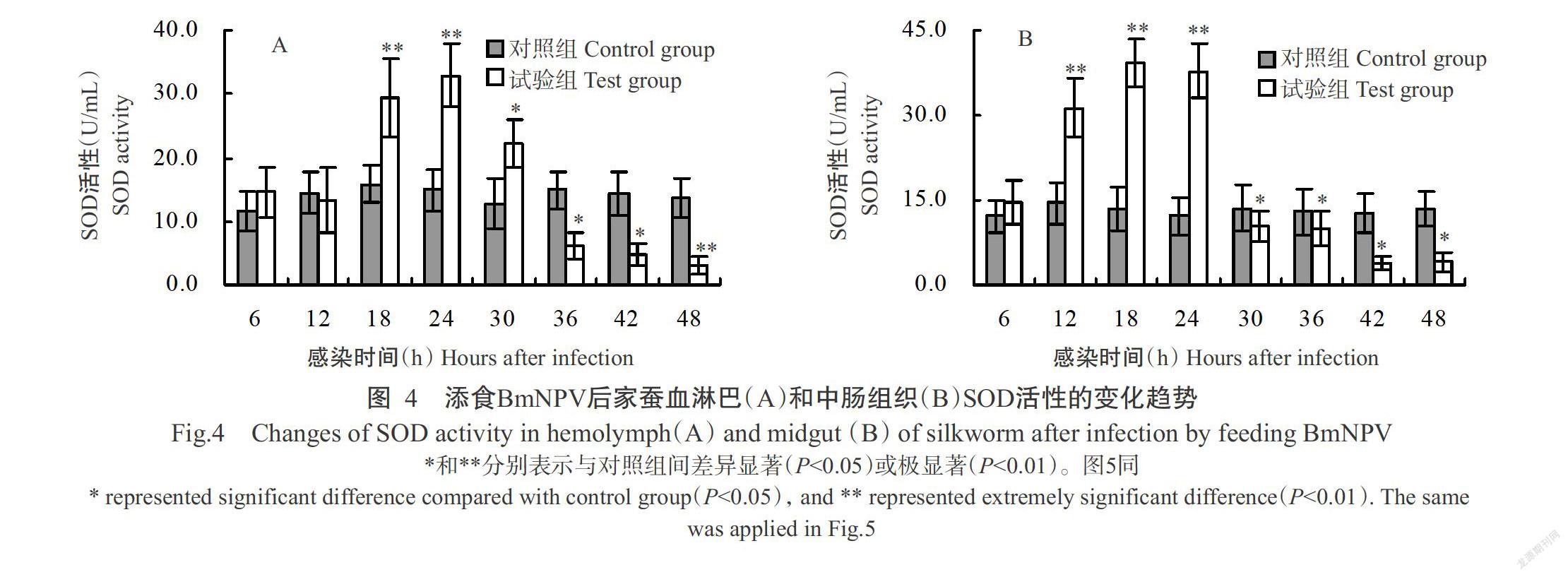
添食BmNPV后,家蠶血淋巴和中腸組織的SOD活性變化如圖4所示。其中,家蠶血淋巴SOD活性在感染初期(6~12 h)與對照組家蠶間無顯著差異(P>0.05,下同),感染中期(18~30 h)SOD活性則顯著高于對照組家蠶(P<0.05,下同),尤其在感染18和24 h時酶活性分別是對照組家蠶的1.9和2.3倍,其差異達極顯著水平(P<0.01,下同),但感染后期(36~48 h)SOD活性顯著或極顯著低于對照組家蠶(圖4-A);家蠶中腸組織SOD活性在感染12、18和24 h分別是對照組家蠶的2.2、2.9和3.1倍,其差異均達極顯著水平,從感染30 h起SOD活性開始急劇下降,至感染42和48 h時酶活性最低,約為對照組家蠶的30%(圖4-B)。
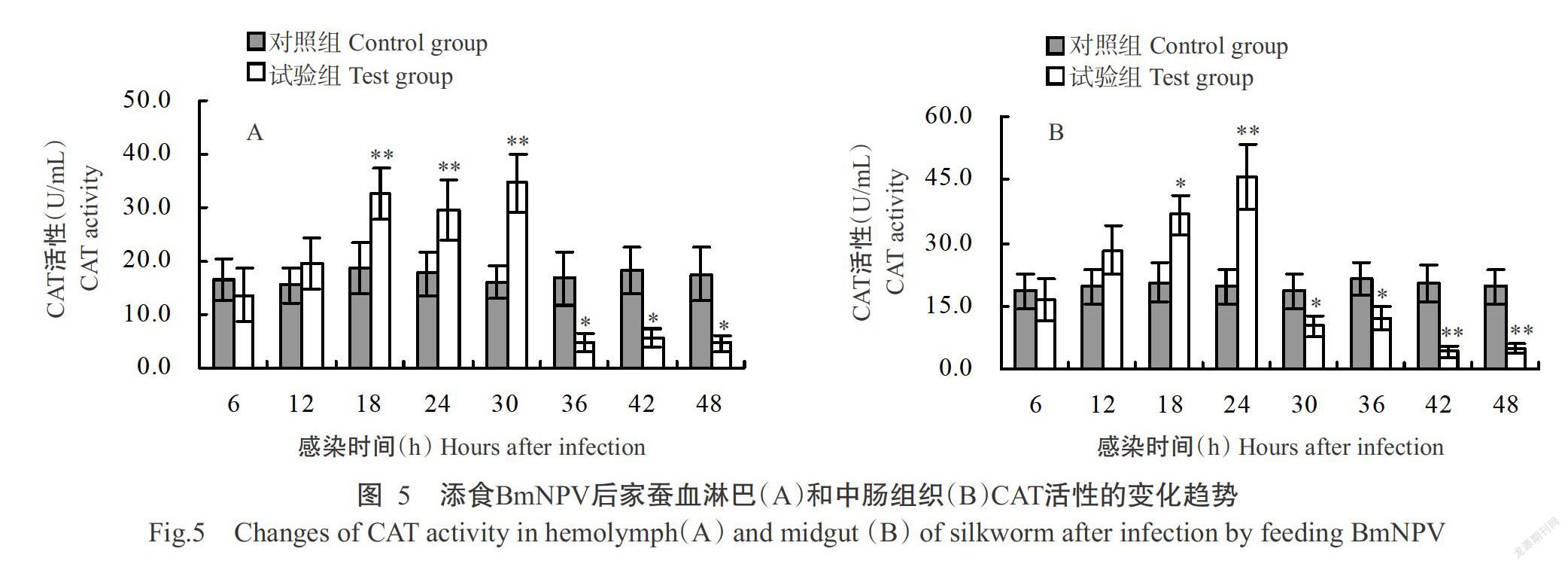
2. 4 家蠶感染BmNPV后血淋巴和中腸組織CAT活性的變化
家蠶感染BmNPV后6 h內,其血淋巴和中腸組織CAT活性略有下降,從感染12 h起CAT活性開始呈上升趨勢(圖5)。其中,家蠶血淋巴CAT活性在感染中期(18~30 h)持續高于對照組家蠶,且差異達極顯著水平,至感染后期(36~48 h)則顯著低于對照組家蠶(圖5-A);家蠶中腸組織CAT活性僅在感染18和24 h時顯著或極顯著高于對照組家蠶,從感染30 h起開始持續下降,至感染48 h降至最低值,約為對照組家蠶的18%,其差異達極顯著水平(圖5-B)。
3 討論
當昆蟲遭受外源病原微生物侵染時,一方面其先天性免疫防御通路被激活,產生抗菌物質或其他免疫活性因子,以保護機體免受傷害(Xia et al.,2017);另一方面,蟲體正常生理代謝過程被擾亂,活性氧平衡系統被破壞,自由基濃度升高,而機體保護酶SOD和CAT在消除過量氧自由基方面發揮重要作用(Kumar et al.,2003)。已有研究表明,高溫或低溫脅迫均會引起家蠶、柞蠶及琥珀蠶的血淋巴SOD和CAT活性在一定時間范圍內出現先升高后降低的變化趨勢(夏潤璽等,2009;袁燕萍等,2010;楊偉克等,2017)。此外,在真菌侵染早期,昆蟲體內的SOD和CAT活性呈上升趨勢,即自身防御能力增強;而在侵染后期,抗氧化能力急劇減弱并出現明顯的感病癥狀(張仙紅等,2006;李世廣等,2016;王樹昌等,2017)。
本研究結果顯示,家蠶中腸組織Bmsod和Bmcat基因的表達及SOD和CAT活性從感染BmNPV后12 h開始增加,而在血淋巴中Bmsod和Bmcat基因表達及SOD和CAT活性的增加相對滯后。中腸是家蠶阻止外源病原微生物及其他毒素等物質侵入的第一道屏障,而血淋巴處于一個相對封閉的循環流動狀態,貫穿于整個蠶體,其中含有許多免疫活性物質和抗氧化酶類,以維持機體正常的生理代謝(馮麗春和沈衛德,2015)。經口感染BmNPV,病毒首先攻擊家蠶的中腸,破壞其抗氧化防御系統的平衡,進而侵染血淋巴系統,誘發抗氧化反應應答,即家蠶中腸的抗氧化響應略早于血淋巴。
添食BmNPV后,家蠶血淋巴和中腸組織的Bmcat和Bmsod基因表達均在感染中期上調表達、感染后期下調表達,SOD和CAT活性也呈先增加后急劇降低的變化趨勢,與BmNPV侵染家蠶的生理進程一致,說明在病毒感染早期,病毒粒子相對較少,不足以打破家蠶抗氧化系統的平衡,抗氧化酶活性仍維持在正常水平;隨著感染時間的延長,病毒不斷增殖,機體抗氧化系統開始受損,此時抗氧化酶相關基因被誘導表達,SOD和CAT活性迅速增強以消除活性氧自由基;至病毒感染后期,病毒已大量增殖,產生的毒素等突破蠶體免疫防線,其抗氧化系統嚴重紊亂,Bmcat和Bmsod基因及其酶活性急劇下降,最終呈現發病癥狀甚至死亡。
4 結論
BmNPV侵染能影響家蠶機體相關保護酶活性及其基因轉錄水平,SOD和CAT活性及其基因表達量在感染中期增加,感染后期則急劇降低,提示抗氧化酶SOD和CAT在家蠶抗病毒過程中發揮重要作用。
參考文獻:
白興榮,冉瑞法,董占鵬,董家紅,黃平. 2010. 云南不同地區BmNPV對家蠶致病力的研究[J]. 西南農業學報,23(6):2098-2101. [Bai X R,Ran R F,Dong Z P,Dong J H,Huang P. 2010. Study on virulence of BmNPV to Bombyx mori in different areas of Yunnan[J]. Southwest China Journal of Agricultural Sciences,23(6):2098-2101.]
方允中,鄭榮梁. 2008. 自由基生物學的理論與應用[M]. 北京:科學出版社. [Fang Y Z,Zheng R L. 2008. Theory and application of free radical biology[M]. Beijing:Science Press.]
馮麗春,沈衛德. 2015. 蠶體解剖生理學[M]. 北京:高等教育出版社. [Feng L C,Shen W D. 2015. Anatomy and physio-logy of silkworm[M]. Beijing:Higher Education Press.]
李世廣,竇婷婷,付小伶,劉祝琴,何超. 2016. 菜青蟲感染金龜子綠僵菌后體內幾種保護酶活性的變化[J]. 植物保護,42(3):133-136. [Li S G,Dou T T,Fu X L,Liu Z Q,He C. 2016. Changes of protective enzyme activities in Pieris rapae infection by Metarhizium anisopliae[J]. Plant Protection,42(3):133-136.]
邵榆嵐,唐芬芬,張一川,張永紅,朱峰,白興榮. 2017. 云南蠶區家蠶品種資源對家蠶核型多角體病毒的抗性評價分析[J]. 南方農業學報,48(3):517-523. [Shao Y L,Tang F F,Zhang Y C,Zhang Y H,Zhu F,Bai X R. 2017. Resistance of silkworm variety resources from Yunnan sericultural areas to Bombyx mori nucleopolyhedrovirus(BmNPV)[J]. Journal of Southern Agriculture,48(3):517-523.]
唐芬芬,楊偉克,張永紅,朱峰,邵榆嵐,白興榮. 2015. 喂食家蠶核型多角體病毒誘導家蠶抗菌肽基因表達[J]. 南方農業學報,46(12):2191-2195. [Tang F F,Yang W K,Zhang Y H,Zhu F,Shao Y L,Bai X R. 2015. Expression of antimicrobial peptide genes in Bombyx mori induced by feeding B. mori nucleopolyhedrovirus[J]. Journal of Southern Agriculture,46(12):2191-2195.]
唐芬芬,楊偉克,朱峰,邵榆嵐,張永紅,白興榮. 2016. 家蠶核型多角體病毒對家蠶酚氧化酶活性及其基因表達的影響[J]. 中國農學通報,32(32):25-28. [Tang F F,Yang W K,Zhu F,Shao Y L,Zhang Y H,Bai X R. 2016. BmNPV affecting the activity and gene expression of phenoloxidase in Bombyx mori[J]. Chinese Agricultural Science Bulletin,32(32):25-28.]
王樹昌,赫榮帆,陸敏泉,黃華平,王娜玉,郭錫杰,耿濤. 2017. 球孢白僵菌侵染對家蠶抗氧化水平的影響[J]. 熱帶作物學報,38(11):2136-2144. [Wang S C,He R F,Lu M Q,Huang H P,Wang N Y,Guo X J,Geng T. 2017. Effects on antioxidant levels of silkworm larvae infected with Beauveria bassiana[J]. Chinese Journal of Tropical Crops,38(11):2136-2144.]
夏潤璽,曹慧穎,劉限,楊瑞生,劉彥群,秦利,王學英. 2009. 高溫條件下柞蠶血淋巴過氧化氫酶活性的變化[J]. 蠶業科學,35(2):415-417. [Xia R X,Cao H Y,Liu X,Yang R S,Liu Y Q,Qin L,Wang X Y. 2009. Change of catalase activity in haemolymph of Antheraea pernyi under high temperature[J]. Science of Sericulture,35(2):415-417.]
楊偉克,唐芬芬,劉增虎,董占鵬,鐘鍵. 2017. 高溫和低溫條件下琥珀蠶血淋巴SOD及CAT活性的變化[J]. 江蘇農業科學,45(1):153-155. [Yang W K,Tang F F,Liu Z H,Dong Z P,Zhong J. 2017. Changes of the CAT and SOD activities in the haemolymph of Antheraea assamensis under low and high temperature[J]. Jiangsu Agricultural Sciences,45(1):153-155.]
袁燕萍,趙林川,魏廣衛,王麗,張彩霞,鮑忠贊,司馬楊虎. 2010. 家蠶品種7532和大造在高溫沖擊下中腸抗氧化酶活性的變化[J]. 蠶業科學,36(4):692-696. [Yuan Y P,Zhao L C,Wei G W,Wang L,Zhang C X,Bao Z Z,Sima Y H. 2010.? Activity variations of antioxidant enzymes in midguts of Bombyx mori strians 7532 and Dazao under high temperature shock[J]. Science of Sericulture,36(4):692-696.]
張仙紅,王宏民,李文英,賀運春,郝赤. 2006. 菜青蟲感染玫煙色擬青霉后血淋巴蛋白質含量及幾種保護酶活力的變化[J]. 昆蟲學報,49(2):230-234. [Zhang X H,Wang H M,Li W Y,He Y C,Hao C. 2006. Changes in the protein content in haemolymph and activities of protective enzymes of Pieris rapae infented by Paecilomyces fumo-soroseus(Deuteromycotina:Hyphomycetes)[J]. Acta Entomologica Sinica,49(2):230-234.]
Geng T,Huang Y X,Hou C X,Qin G X,Lü D D,Guo X J. 2016a. Inductive expression patterns of genes related to Toll signaling pathway in silkworm(Bombyx mori) upon Beauveria bassiana infection[J]. Journal of Asia-Pacific Entomology,19(3):861-868.
Geng T,Lv D D,Huang Y X,Hou C X,Qin G X,Guo X J. 2016b. JAK/STAT signaling pathway-mediated immune response in silkworm(Bombyx mori) challenged by Beauveria bassiana[J]. Gene,595(1):69-76.
Kumar S,Christophides G K,Cantera R,Charles B,Han Y S,Meister S,Dimopoulos G,Kafatos F C,Barillas-Mury C. 2003. The role of reactive oxygen species on Plasmodium melanotic encapsulation in Anopheles gambiae[J]. Procee-dings of the National Academy of Sciences of the United States of America,100(24):14139-14144.
Lesser M P. 2006. Oxidative stress in marine environments:Biochemistry and physiological ecology[J]. Annual Re-
view of Physiology,68(1):253-278.
Livak K J,Schmittgen T D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method[J]. Methods,25(4):402-408.
Wang Y,Oberley L W,Murhammer D W. 2001. Antioxidant defense systems of two lipidopteran insect cell lines[J]. Free Radical Biology and Medicine,30(11):1254-1262.
Xia X F,You M S,Rao X J,Yu X Q. 2017. Insect C-type lectins in innate immunity[J]. Developmental and Comparative Immunology,83:70-79.
Zhao L C,Shi L G. 2009. Metabolism of hydrogen peroxide in univoltine and polyvoltine strains of silkworm(Bombyx mori)[J]. Comparative Biochemistry and Physiology(Part B),152:339-345.
(責任編輯 蘭宗寶)
