祁連山中段退化高寒草地土壤細(xì)菌群落分布特征
2019-08-30 02:27:00李海云姚拓馬亞春張慧榮路曉雯楊曉蕾夏東慧張建貴高亞敏
草業(yè)學(xué)報(bào) 2019年8期
李海云,姚拓,馬亞春,張慧榮,路曉雯,楊曉蕾,夏東慧,張建貴,高亞敏
(甘肅農(nóng)業(yè)大學(xué)草業(yè)學(xué)院,甘肅 蘭州 730070)
草地在陸地生態(tài)系統(tǒng)中起著重要作用,尤其是在營(yíng)養(yǎng)物質(zhì)和能量循環(huán)過(guò)程扮演著重要角色[1]。近年來(lái)由于對(duì)草地資源的過(guò)度放牧和濫耕亂伐,使草地植被破壞嚴(yán)重,水土流失加劇,草原鼠蟲(chóng)害嚴(yán)重[2-3],導(dǎo)致草地退化、沙化、鹽堿化面積日益擴(kuò)大,近年來(lái)草地退化等生態(tài)問(wèn)題已受到人們的普遍關(guān)注和高度重視[4]。放牧活動(dòng)是影響高寒草地退化的主要驅(qū)動(dòng)因素之一,由于家畜的采食、踐踏和排泄糞便等改變了地表植被覆蓋狀況,造成地表特征的改變,進(jìn)而影響土壤結(jié)構(gòu)的破壞、養(yǎng)分的損耗、土壤微生物區(qū)系和數(shù)量的改變,最終導(dǎo)致草地退化[5]。隨著研究和實(shí)踐的深入,許多研究者認(rèn)識(shí)到草地退化不僅是地表植被和土壤理化特性的退化,更重要的是草地土壤微生物群落結(jié)構(gòu)和多樣性的變化。土壤微生物作為土壤生態(tài)系統(tǒng)的重要組成部分,在土壤有機(jī)質(zhì)分解和營(yíng)養(yǎng)物質(zhì)循環(huán)等生態(tài)系統(tǒng)功能過(guò)程中起主導(dǎo)作用[6]。由于土壤微生物對(duì)生存的微環(huán)境十分敏感,能對(duì)土壤環(huán)境變化做出快速反應(yīng),進(jìn)而影響植物群落物種多樣性和土壤結(jié)構(gòu)的形成。因此,土壤微生物群落結(jié)構(gòu)和多樣性的變化可作為衡量草地生態(tài)系統(tǒng)的健康變化和草地退化或恢復(fù)程度的重要指標(biāo)[7]。目前,關(guān)于草地退化對(duì)土壤微生物的研究,主要集中在可培養(yǎng)微生物、磷脂脂肪酸(phospholipid fatty acid,PLFA)、PCR-DGGE、16S rRNA基因文庫(kù)構(gòu)建技術(shù)等方面[8-11]。這些傳統(tǒng)的研究方法所能反映出的土壤微生物信息十分有限,在很大程度上低估了土壤微生物的物種組成并高估其豐度[12]。隨著高通量測(cè)序技術(shù)的興起,其具有高準(zhǔn)確性、高靈敏度等優(yōu)勢(shì),能夠較為全面和準(zhǔn)確地反映土壤微生物群落結(jié)構(gòu),被廣泛應(yīng)用于各研究領(lǐng)域中[13],彌補(bǔ)了前期土壤微生物傳統(tǒng)研究方法的不足。因此,為探究祁連山中段不同退化草地土壤細(xì)菌群落結(jié)構(gòu)變化規(guī)律,采用高通量測(cè)序技術(shù)分析退化高寒草地土壤微生物群落變化趨勢(shì),為高寒草地退化與恢復(fù)治理提供理論依據(jù)和數(shù)據(jù)參考價(jià)值。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于祁連山中部地段(37°53′ N、101°45′ E),隸屬于甘肅省張掖市肅南縣皇城鎮(zhèn),其南部為冷龍嶺山地,北部為蓋掌達(dá)坂山地,東連天祝藏族自治縣,西接山丹軍馬場(chǎng)。海拔2600~3500 m,屬高原大陸性氣候,全年平均氣溫0.6~3.8 ℃,無(wú)霜期不足140 d,絕對(duì)無(wú)霜期僅45~60 d,年降水量361.6 mm,氣候寒冷多變,四季不分明[14]。
1.2 樣地設(shè)置及土樣采集
依據(jù)草地退化程度劃分相關(guān)劃價(jià)標(biāo)準(zhǔn)[15],在2017年8月對(duì)肅南縣皇城草地植被進(jìn)行調(diào)查,選擇3種退化草地類(lèi)型,分別為:輕度退化草地(lightly degraded grassland,LDG)、中度退化草地(moderately degraded grassland,MDG)和重度退化草地(severely degraded grassland,SDG),具體樣地信息見(jiàn)表1。在每種樣地中,設(shè)置3個(gè)大樣方(50 m×50 m),其內(nèi)隨機(jī)設(shè)置3個(gè)小樣方(50 cm×50 cm),記錄小樣方中植物種類(lèi)、物種數(shù)、蓋度和高度。地上部生物量采用齊地面刈割,將鮮樣在65 ℃下烘干24 h后稱(chēng)重。用土鉆(內(nèi)徑53 mm)采集0~20 cm土壤樣品,土樣混勻后裝入已滅菌袋中迅速帶回,在-80 ℃冰箱中保存以備用。土樣分為兩份,一份用于土壤養(yǎng)分及酶活測(cè)定,另一份用于土壤微生物總DNA提取。

表1 樣地基本信息Table 1 Basic information of the soil samples
1.3 土壤理化性質(zhì)及酶活性測(cè)定
采用鮑士旦[16]的方法測(cè)定土壤理化性質(zhì)pH、含水量、電導(dǎo)率、有機(jī)質(zhì)、全量養(yǎng)分和速效養(yǎng)分含量;采用關(guān)松蔭[17]的方法測(cè)定土壤脫氫酶、氧化還原酶、脲酶、中性磷酸酶、纖維素酶和蔗糖酶活性。
1.4 土壤總DNA提取及細(xì)菌16S rRNA基因擴(kuò)增
采用OMEGA土壤DNA提取試劑盒(上海翊圣生物科技有限公司,上海)對(duì)土壤DNA基因組進(jìn)行提取,每個(gè)樣品3次重復(fù)并進(jìn)行混合,以降低DNA提取過(guò)程中造成的誤差。以提取的DNA為模板(1 ng·μL-1),對(duì)細(xì)菌16S rRNA基因V4~V5區(qū)采用帶Barcode的特異性引物[18](515F和907R)進(jìn)行PCR擴(kuò)增,每個(gè)樣品3次重復(fù)。PCR擴(kuò)增條件參照趙帆等[19]的方法進(jìn)行擴(kuò)增。PCR產(chǎn)物根據(jù)產(chǎn)物濃度進(jìn)行等量混樣,充分混勻后使用2%的瓊脂糖凝膠電泳檢測(cè)PCR產(chǎn)物并切膠回收,產(chǎn)物送至天津諾禾致源生物信息科技有限公司,采用Illumina HiSeq PE250高通量測(cè)序技術(shù)平臺(tái)進(jìn)行測(cè)序分析。
1.5 生物信息學(xué)分析
根據(jù)Barcode序列和PCR擴(kuò)增引物序列從測(cè)定數(shù)據(jù)中拆分出各樣品數(shù)據(jù),截去Barcode和引物序列后對(duì)每個(gè)樣品的reads進(jìn)行拼接[20]后得到原始測(cè)序數(shù)據(jù);原始測(cè)序數(shù)據(jù)經(jīng)質(zhì)檢和嵌合體去除得到有效數(shù)據(jù)(effective tags)。利用Uparse軟件對(duì)樣品的有效序列進(jìn)行OTUs聚類(lèi)(相似度97%以上),用Mothur方法[21]與SILVA軟件的SSUrRNA數(shù)據(jù)庫(kù)[22]進(jìn)行物種注釋(閾值:0.8~1.0);采用PyNAST軟件[23]與GreenGene數(shù)據(jù)庫(kù)中數(shù)據(jù)信息進(jìn)行多序列比對(duì),最后對(duì)測(cè)序數(shù)據(jù)進(jìn)行標(biāo)準(zhǔn)化處理,采用Qiime 1.7.0軟件計(jì)算Alpha多樣性指數(shù)和Beta多樣性。采用R 2.15.3軟件的Vegan軟件包繪制稀釋曲線(xiàn),并進(jìn)行無(wú)度量多維標(biāo)定法分析(non-metric multi-dimensional scaling,NMDS)。
1.6 數(shù)據(jù)分析
所有數(shù)據(jù)均采用SPSS 21.0軟件進(jìn)行處理分析,One-Way ANOVA和Duncan氏新復(fù)極差法分析差異顯著性。采用CANOCO 4.5軟件對(duì)細(xì)菌優(yōu)勢(shì)群落進(jìn)行去趨勢(shì)對(duì)應(yīng)分析(DCA),根據(jù)第一排序軸的梯度范圍(lengths of gradient)數(shù)值,小于3.0,采用冗余分析(RDA);為3.0~4.0,采用RDA分析和典型相關(guān)分析(CCA)均可;大于4.0,采用CCA分析,對(duì)細(xì)菌群落與土壤環(huán)境指標(biāo)間的相互關(guān)系進(jìn)行分析。
2 結(jié)果與分析
2.1 植被特征
3種高寒草地植被特征間差異顯著(P<0.05)。隨著退化程度的加重,植被蓋度、高度、地上生物量均明顯降低,依次為輕度退化>中度退化>重度退化(表2);群落物種多樣性和均勻度指數(shù)的變化為輕度退化>中度退化>重度退化。
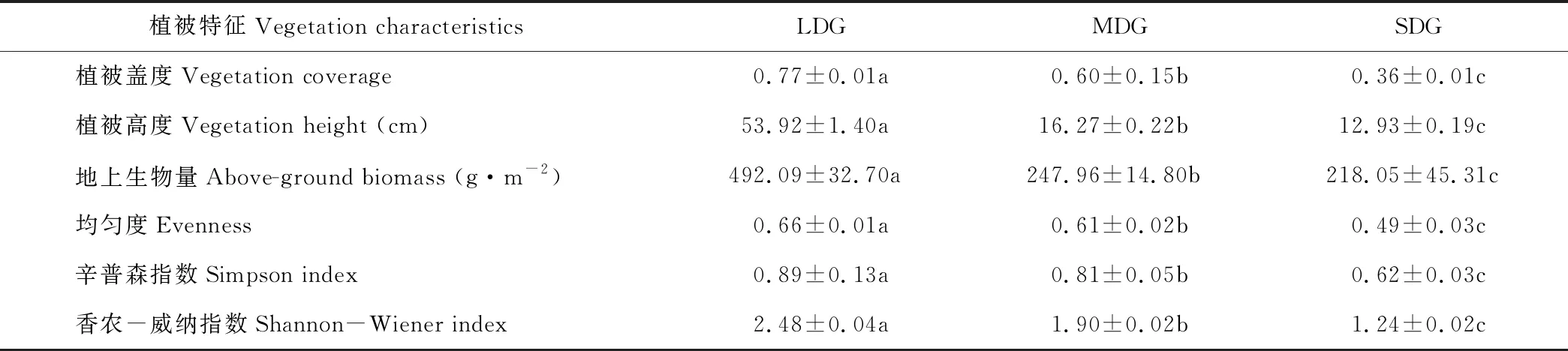
表2 各樣地植被特征Table 2 The basic vegetation characteristics of different soil sampling sites
注:同行不同字母表示差異顯著(P<0.05),下同。
Note: Different letters in the same line mean significant differences at 0.05 level, the same below.
2.2 土壤理化性質(zhì)
從表3可以看出,各樣地土壤理化性質(zhì)及酶活性間均差異顯著(P<0.05)。隨著草地退化程度的加重,土壤含水量、有機(jī)碳、有機(jī)質(zhì)、全氮、全磷、全鉀、速效磷、速效氮、速效鉀均呈逐漸減小趨勢(shì);土壤電導(dǎo)率呈逐漸增大趨勢(shì);土壤pH、脲酶、中性磷酸酶、脫氫酶、蔗糖酶和氧化還原酶均呈先增大后減小趨勢(shì);土壤纖維素酶呈先減小后增大趨勢(shì)。
2.3 土壤細(xì)菌群落豐度與Alpha多樣性分析
通過(guò)Illumina HiSeq PE250高通量測(cè)序,共得到有效序列257971條,質(zhì)控過(guò)濾和去除嵌合體后得到219017條優(yōu)質(zhì)序列,聚類(lèi)共得到2004個(gè)OTUs。各樣品文庫(kù)的覆蓋度均在99.6%以上,并結(jié)合樣品稀釋曲線(xiàn)均趨于平緩,說(shuō)明本研究測(cè)序數(shù)據(jù)合理,能夠準(zhǔn)確反映出土壤細(xì)菌群落的真實(shí)信息(圖1)。3種退化草地土壤細(xì)菌群落豐富度指數(shù)(Chao1和ACE指數(shù))依次為L(zhǎng)DG>MDG>SDG(表4);細(xì)菌群落多樣性指數(shù)(Simpson和Shannon-Wiener指數(shù))依次為L(zhǎng)DG>SDG>MDG。如圖2所示,所有樣品中共有OTUs數(shù)目為1317個(gè),其中LDG、MDG和SDG中所特有的OTUs數(shù)目分別為112、100和47個(gè)。
2.4 不同退化草地土壤細(xì)菌群落分布特征分析
2.4.1門(mén)水平上的群落組成 通過(guò)測(cè)序發(fā)現(xiàn),3種退化草地土壤細(xì)菌中相對(duì)豐度>1%的菌門(mén)為:放線(xiàn)菌門(mén)(Actinobacteria)、厚壁菌門(mén)(Firmicutes)、酸桿菌門(mén)(Acidobacteria)、變形菌門(mén)(Proteobacteria)、芽單胞菌門(mén)(Gemmatimonadetes)、綠彎菌門(mén)(Chloroflexi)、疣微菌門(mén)(Verrucomicrobia)、浮霉菌門(mén)(Planctomycetes)、擬桿菌門(mén)(Bacteroidetes)和硝化螺旋菌門(mén)(Nitrospirae),豐度低于1%的類(lèi)群占1.57%~3.07%。3種退化草地土壤細(xì)菌在門(mén)分類(lèi)水平上,相對(duì)豐度存在一定的變化趨勢(shì)(圖3)。隨著草地退化程度加劇,放線(xiàn)菌門(mén)、酸桿菌門(mén)、芽單胞菌門(mén)、綠彎菌門(mén)、浮霉菌門(mén)、擬桿菌門(mén)相對(duì)豐度呈先減小后增大趨勢(shì),厚壁菌門(mén)、疣微菌門(mén)和硝化螺旋菌門(mén)相對(duì)豐度呈先增大后減小趨勢(shì),變形菌門(mén)相對(duì)豐度呈逐漸減小趨勢(shì);功能菌群Proteobacteria/Acidobacteria呈先增大后減小的趨勢(shì)。

表3 各樣地土壤基本理化性質(zhì)Table 3 The basic soil physiochemical indexes of different soil sampling sites
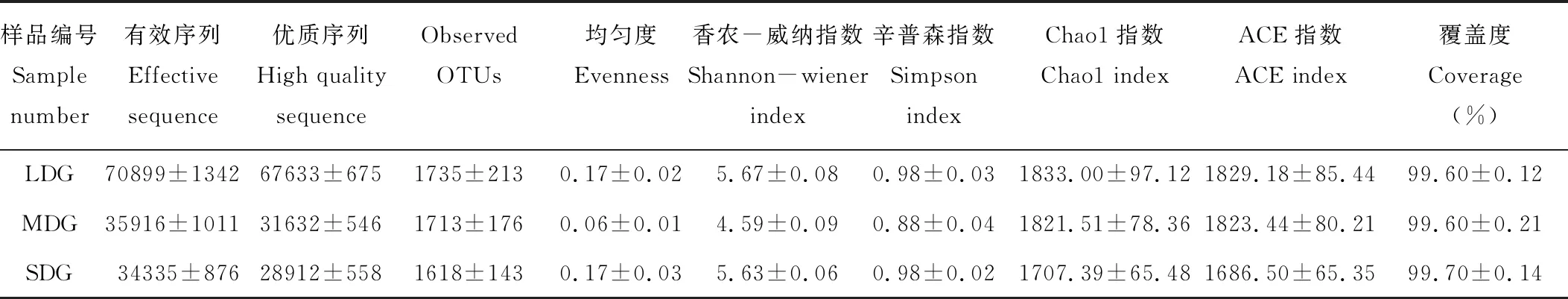
表4 樣品序列數(shù)統(tǒng)計(jì)、豐富度與多樣性指數(shù)Table 4 Sample sequence numbers statistics, richness and diversity index
2.4.2屬水平上的群落組成 在屬分類(lèi)水平上,相對(duì)豐度>1%的菌屬為:乳球菌屬(Lactococcus)、土壤紅桿菌屬(Solirubrobacter)、未分類(lèi)RB41屬、Gaiella屬、類(lèi)諾卡氏屬(Nocardioides)、假單胞菌屬(Pseudomonas)、鞘氨醇單胞菌屬(Sphingomonas)、未分類(lèi)H16屬、芽球菌屬(Blastococcus)、Pseudarthrobacter、鏈球菌屬(Streptococcus)和芽胞桿菌屬(Bacillus),相對(duì)豐度低于1%的類(lèi)群占48.94%~73.94%。3種退化草地土壤在屬分類(lèi)水平上,相對(duì)豐度存在一定變化趨勢(shì)(圖4)。隨著退化程度加劇,乳球菌屬、假單胞菌屬、鞘氨醇單胞菌屬、未分類(lèi)H16屬、Pseudarthrobacter和鏈球菌屬相對(duì)豐度呈先增大后減小趨勢(shì),土壤紅桿菌屬、未分類(lèi)RB41屬、Gaiella、類(lèi)諾卡氏屬和芽球菌屬相對(duì)豐度呈先減小后增大趨勢(shì),芽胞桿菌屬相對(duì)豐度呈逐漸減小趨勢(shì)。
2.5 Beta多樣性分析
LDG與MDG樣地間加權(quán)Unifrac距離相異系數(shù)分別為0.247和0.245(圖5);LDG與SDG樣地間加權(quán)Unifrac距離相異系數(shù)分別為0.184和0.216;MDG和SDG樣地間加權(quán)Unifrac距離相異系數(shù)分別為0.294和0.258。結(jié)果表明,3種退化草地土壤間物種多樣性差異為:LDG>SDG>MDG。為了研究不同樣品間的相似性,對(duì)樣品進(jìn)行了UPGMA聚類(lèi)分析。在門(mén)分類(lèi)水平上,通過(guò)加權(quán)Unifrac距離的UPGMA聚類(lèi)分析,LDG與SDG樣地間細(xì)菌群落組成及豐度相似性較高,與MDG樣地間細(xì)菌群落組成差異較大(圖6)。
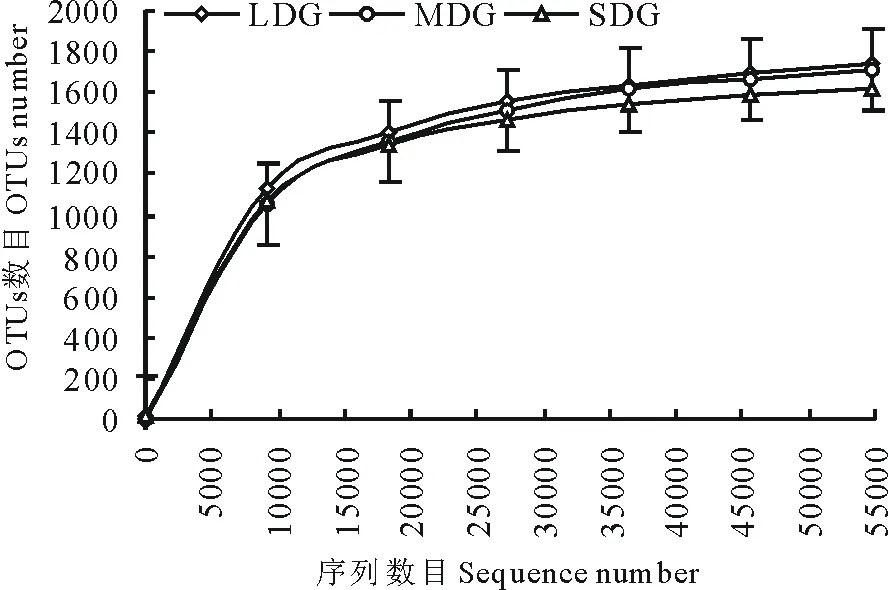
圖1 樣品稀釋曲線(xiàn)Fig.1 Rarefaction curves for samples
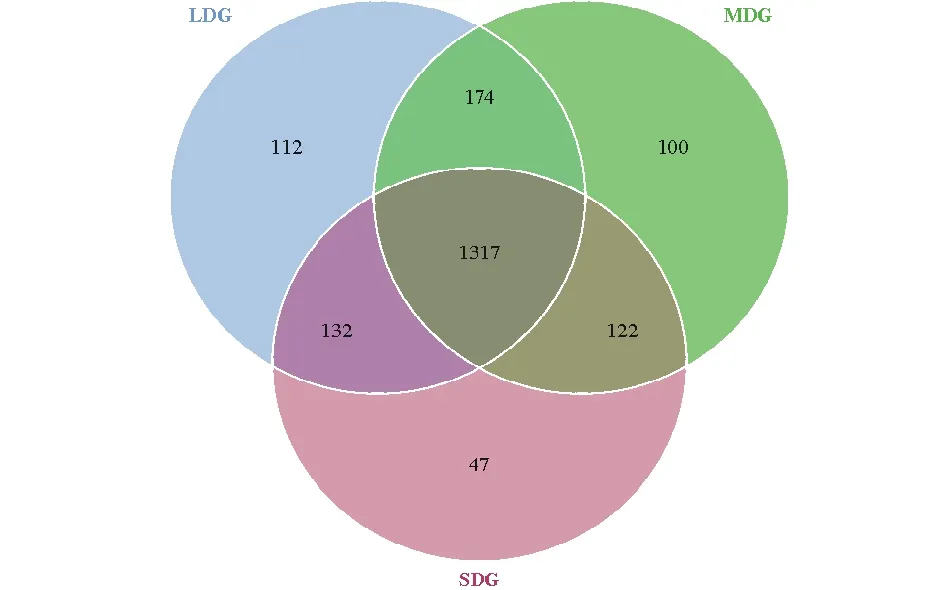
圖2 樣品韋恩圖Fig.2 Venn diagrams of samples
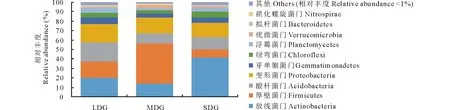
圖3 門(mén)分類(lèi)水平下的細(xì)菌群落相對(duì)豐度Fig.3 Relative abundance of bacterial community at phylum level
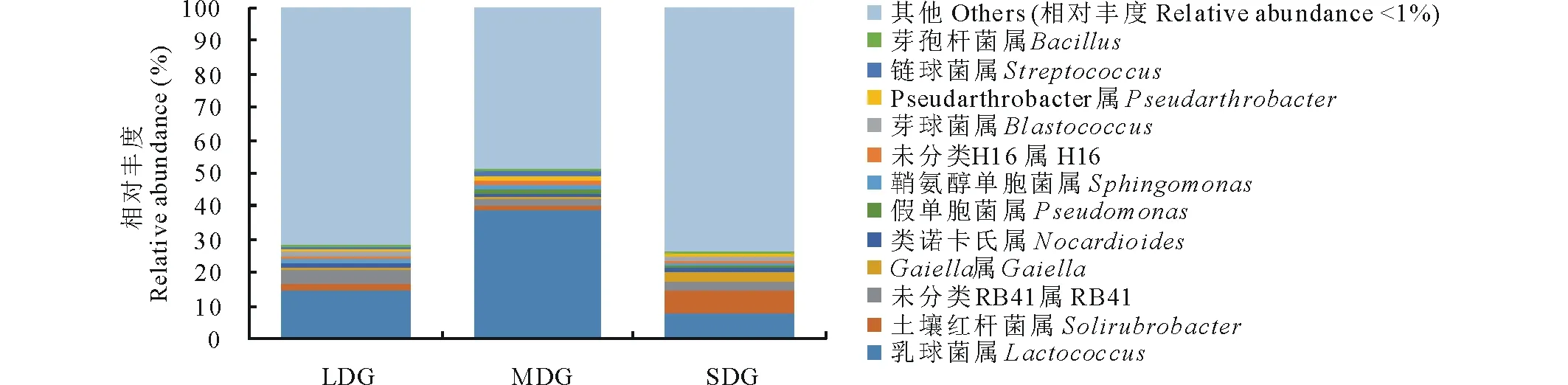
圖4 屬分類(lèi)水平下的細(xì)菌群落相對(duì)豐度Fig.4 Relative abundances of bacterial community at genus level
2.6 細(xì)菌優(yōu)勢(shì)類(lèi)群與土壤環(huán)境因子間RDA分析
通過(guò)RDA分析表明:第一、二排序軸累計(jì)解釋率分別為68.1%和11.9%(圖7)。其中,厚壁菌門(mén)與氧化還原酶呈顯著正相關(guān)(P<0.05);放線(xiàn)菌門(mén)與中性磷酸酶、蔗糖酶、氧化還原酶、含水量呈極顯著負(fù)相關(guān)(P<0.01),與電導(dǎo)率呈極顯著正相關(guān)(P<0.01),與速效磷、速效鉀呈顯著負(fù)相關(guān)(P<0.05);變形菌門(mén)與脲酶、磷酸酶、脫氫酶、蔗糖酶、含水量、全N、全P、全K、速效N、速效P、速效K、有機(jī)質(zhì)、有機(jī)碳呈極顯著正相關(guān)(P<0.01),與pH、電導(dǎo)率和纖維素酶呈極顯著負(fù)相關(guān)(P<0.01),與氧化還原酶呈顯著正相關(guān)(P<0.05);酸桿菌門(mén)與脲酶、脫氫酶、全N、全P、全K、速效N、有機(jī)質(zhì)、有機(jī)碳呈極顯著正相關(guān)(P<0.01),與pH、纖維素酶呈極顯著負(fù)相關(guān)(P<0.01),與蔗糖酶、速效P、速效K呈顯著正相關(guān)(P<0.05);芽單胞菌門(mén)與pH呈極顯著負(fù)相關(guān)(P<0.01);浮霉菌門(mén)與全P、速效N呈極顯著正相關(guān)(P<0.01),與pH呈極顯著負(fù)相關(guān)(P<0.01),與脲酶、全N、有機(jī)質(zhì)、有機(jī)碳呈顯著正相關(guān)(P<0.05);疣微菌門(mén)與pH呈顯著正相關(guān)(P<0.05);硝化螺旋菌門(mén)與脲酶、全N、全P、速效N、有機(jī)質(zhì)、有機(jī)碳呈極顯著負(fù)相關(guān)(P<0.01),與pH呈極顯著正相關(guān)(P<0.01);擬桿菌門(mén)與pH呈顯著負(fù)相關(guān)(P<0.05)。綜上,土壤酶活性和土壤理化性質(zhì)均對(duì)細(xì)菌群落的分布均有影響,其中,土壤pH是土壤細(xì)菌群落分布的主要驅(qū)動(dòng)因子。
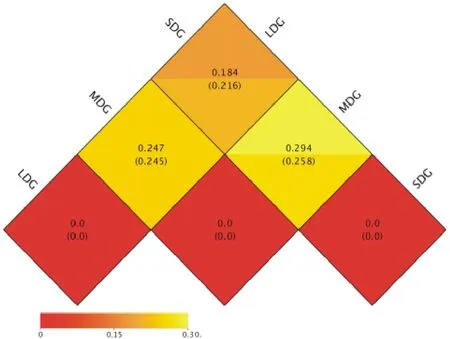
圖5 各樣地OTUs間的Heatmap分析Fig.5 Heatmap analysis between OTUs of sample plots level
3 討論
草地植被退化和土壤理化性質(zhì)的改變,將直接影響土壤微生物的生物量和群落結(jié)構(gòu)多樣性[24-27]。戴雅婷等[13]采用PCR-DGGE技術(shù)對(duì)內(nèi)蒙古退化荒漠草原土壤細(xì)菌群落結(jié)構(gòu)特征進(jìn)行研究發(fā)現(xiàn),隨退化程度的加劇,細(xì)菌多樣性指數(shù)由高到低依次為:輕度>重度>中度,本研究所得結(jié)果與其是一致的。本研究中相對(duì)豐度較高的細(xì)菌菌門(mén)為厚壁菌門(mén)、放線(xiàn)菌門(mén)、變形菌門(mén)和酸桿菌門(mén),真菌菌門(mén)為子囊菌門(mén)和擔(dān)子菌門(mén),這與多數(shù)有關(guān)高寒草地土壤微生物群落組成所得研究結(jié)果是基本一致的。Li等[28]對(duì)青藏高原地區(qū)高寒草地土壤微生物群落結(jié)構(gòu)組成進(jìn)行研究發(fā)現(xiàn)變形菌門(mén),放線(xiàn)菌門(mén)和酸桿菌門(mén)是該地區(qū)的優(yōu)勢(shì)種群。Zhou等[29]對(duì)青藏高原地區(qū)不同退化高寒草地土壤微生物群落結(jié)構(gòu)組成進(jìn)行研究發(fā)現(xiàn)放線(xiàn)菌門(mén)、變形菌門(mén)、酸桿菌門(mén)和綠彎菌門(mén)是該地區(qū)的優(yōu)勢(shì)種群。這表明不同高寒地區(qū)的退化草地土壤類(lèi)型間微生物種群結(jié)構(gòu)分布雖存在明顯差異,但優(yōu)勢(shì)菌群基本相同。隨著退化程度加劇,厚壁菌門(mén)整體上呈先升高后降低的趨勢(shì)。這是因?yàn)楹癖诰T(mén)適于生長(zhǎng)在可利用養(yǎng)分低的環(huán)境中,而高寒地區(qū)的低溫環(huán)境又限制了有機(jī)物的可利用性,不利于它們的生長(zhǎng);另外,厚壁菌門(mén)在惡劣環(huán)境條件時(shí)進(jìn)入休眠狀態(tài),尤其是能形成芽孢和孢子。隨著退化程度加劇,放線(xiàn)菌門(mén)、酸桿菌門(mén)和變形菌門(mén)的豐度呈先減小后增大趨勢(shì),是由于高寒環(huán)境中土壤整體營(yíng)養(yǎng)狀況較低,且土壤pH屬堿性導(dǎo)致的[30]。許多研究表明,在多種土壤生態(tài)系統(tǒng)中pH通常與細(xì)菌群落結(jié)構(gòu)有顯著的相關(guān)性,在近中性時(shí)多樣性最高,pH一旦偏離中性,微生物群落會(huì)受到環(huán)境脅迫壓力,多樣性也因環(huán)境脅迫的選擇而降低[31]。變形菌門(mén)是土壤中最主要的細(xì)菌類(lèi)群,該類(lèi)群的代謝活動(dòng)是土壤中最主要的細(xì)菌活動(dòng)。酸桿菌門(mén)一般存在于營(yíng)養(yǎng)比較匱乏的環(huán)境,放線(xiàn)菌門(mén)能夠降解纖維素和幾丁質(zhì),是土壤養(yǎng)分供給的主要來(lái)源,并且產(chǎn)生的孢子能夠抵抗外界不利的環(huán)境條件,使得它成為高寒草地土壤的優(yōu)勢(shì)菌群[32]。地上植被通過(guò)影響土壤中的有機(jī)碳、有機(jī)氮、土壤水分、溫度、通氣性和pH值等來(lái)影響土壤微生物多樣性。植物群落多樣性越豐富,凋落物和根系分泌物組成就越豐富,土壤微生物多樣性也就越高[33]。植物多樣性的增加往往伴隨著植物生物量的增加,增加了凋落物和根系等有機(jī)物質(zhì)的吸積,從而有利于土壤OC和N的積累[34]。本研究樣地地處高寒環(huán)境,不同退化程度草地中土壤微生物群落結(jié)構(gòu)的變化也相應(yīng)地受到地上植被的影響,植被特征和細(xì)菌群落結(jié)構(gòu)多樣性顯著相關(guān)(P<0.05),這說(shuō)明地上植被群落結(jié)構(gòu)的發(fā)展變化對(duì)細(xì)菌多樣性產(chǎn)生影響。本研究發(fā)現(xiàn),隨著草地退化演替的發(fā)展,土壤細(xì)菌群落結(jié)構(gòu)組成和多樣性差異較大,這與植被和土壤理化性質(zhì)是顯著相關(guān)的。通過(guò)植被、土壤理化性質(zhì)與細(xì)菌群落結(jié)構(gòu)組成間的RDA分析發(fā)現(xiàn),影響祁連山高寒草地土壤細(xì)菌群落結(jié)構(gòu)組成的重要驅(qū)動(dòng)因子是pH。大量研究也報(bào)道了pH對(duì)微生物的分布規(guī)律存在著普遍的影響[30]。本次研究也發(fā)現(xiàn),pH對(duì)土壤微生物群落有顯著性影響,但微生物種群對(duì)pH的響應(yīng)是不同的,如土壤中酸桿菌門(mén)的豐度與pH值呈負(fù)相關(guān),放線(xiàn)菌門(mén)的豐度與pH值呈正相關(guān)[30]。

圖6 門(mén)水平上物種組成的UPGMA聚類(lèi)樹(shù)圖Fig.6 UPGMA clustering tree diagram of species composition at the gate
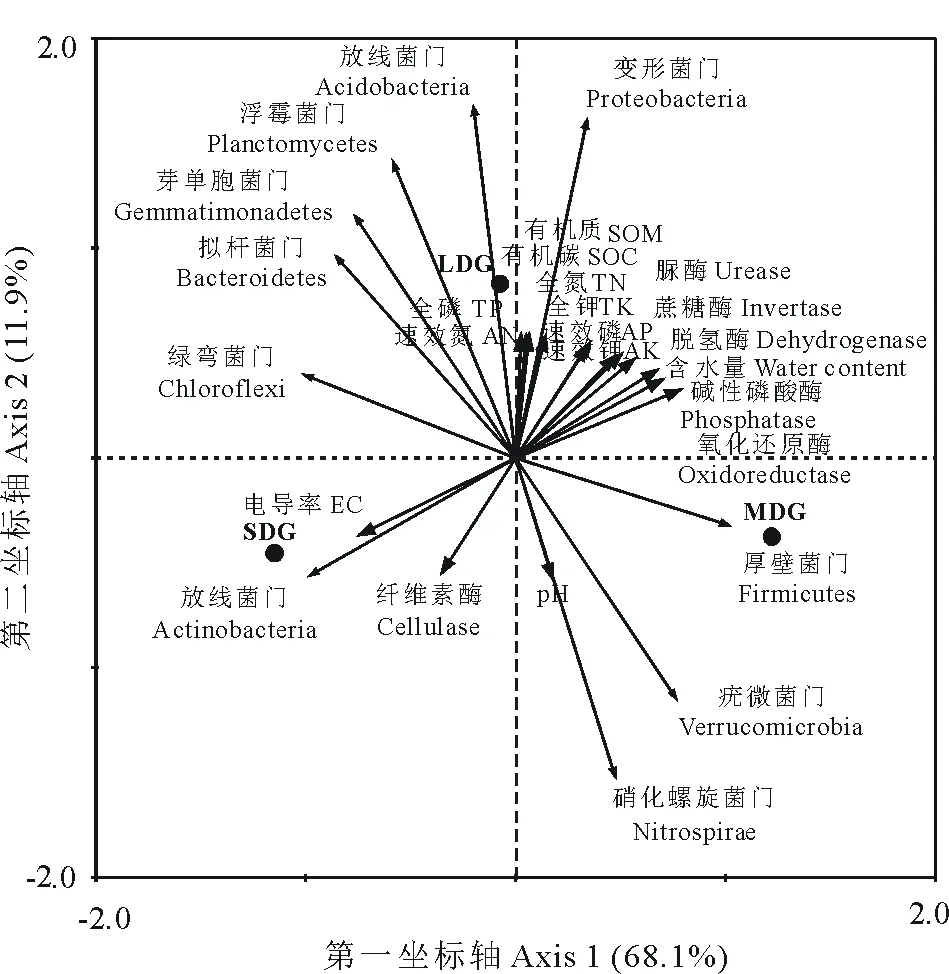
圖7 細(xì)菌群落與土壤環(huán)境因子的冗余分析Fig.7 Redundancy analysis for bacterial community and soil environmental factors
4 結(jié)論
1) 植被蓋度、高度、地上生物量和植物多樣性指數(shù)隨退化程度加劇均明顯降低,依次為輕度退化>中度退化>重度退化;各樣地土壤理化性質(zhì)及酶活性間均差異顯著(P<0.05)。
2) 通過(guò)高通量測(cè)序,共得到有效序列257971條,優(yōu)質(zhì)序列219017條,OTUs 2004個(gè);土壤細(xì)菌群落豐富度指數(shù)表現(xiàn)為L(zhǎng)DG>MDG>SDG;多樣性指數(shù)表現(xiàn)為L(zhǎng)DG>SDG>MDG;Beta多樣性為L(zhǎng)DG>SDG>MDG;3種退化草地土壤細(xì)菌群落的變化主要在于微生物量的變化,對(duì)細(xì)菌群落結(jié)構(gòu)的影響并不明顯。
3) RDA分析表明:土壤酶活性和土壤理化性質(zhì)均對(duì)細(xì)菌群落的分布具有影響。其中土壤pH是影響土壤細(xì)菌群落分布的主要驅(qū)動(dòng)因子。
