自然發酵腐乳中細菌多樣性評價
2019-08-27 07:44:56李娜崔夢君馬佳佳余海忠張振東郭壯趙慧君
食品研究與開發 2019年16期
關鍵詞:分析
李娜,崔夢君,馬佳佳,余海忠,張振東,郭壯,2,趙慧君,*
(1.湖北文理學院食品科學技術學院鄂西北傳統發酵食品研究所,湖北 襄陽 441053;2.恩施市公共檢驗檢測中心,湖北 恩施 445000)
腐乳,又稱豆腐乳,是一種中國傳統的大豆發酵食品,也是由微生物作用的代表性豆制品[1]。根據加工方式不同,腐乳可分為細菌型腐乳、霉菌型腐乳、酶法發酵以及自然發酵腐乳[2]。在腐乳自然發酵過程中,由于制作環境和人工因素等的影響,導致其蘊含的微生物的種類豐富多樣,從而使腐乳具有別樣風味。眾多學者對腐乳中微生物群落構成進行了研究,其中程永強等[3]在低溫發酵腐乳中發現1 株嗜低溫的毛霉——黃色毛霉(Mucor flavus),同時姚翔等[4]在益陽自然發酵腐乳中分離出總狀毛霉(Mucor racemosus)和魯氏毛霉(Mucor roxianus),而魯菲等[5]在青方腐乳中分離出植物乳桿菌(Lactobacillus plantarum)和短小奇異菌,然而關于湖北地區腐乳微生物多樣性的研究較少。
變性梯度凝膠電泳(polymerase chain reaction-denaturing gradient gel electrophoresis,PCR-DGGE)技術是一種可以對微生物的群落結構及遺傳多樣性進行連續分析的技術[6],同時具有重復性好、檢測結果可靠以及應用范圍廣等優點[7],目前應用于葡萄酒[8]、香腸[9]和奶酪[10]等領域。Illumina Miseq 高通量測序技術可以從宏基因組層面對樣品中的微生物多樣性進行全方位且客觀的分析評價,同時克服了傳統微生物學手段耗時長、工作量大等缺點[11],廣泛應用于研究腸道菌群[12]、發酵食品[13]以及環境檢測[14]等方面。
本試驗以恩施地區自然發酵腐乳為研究對象,利用PCR-DGGE 結合Illumina Miseq 高通量測序技術相結合的手段對腐乳中的微生物群落組成及多樣性進行解析,同時利用傳統微生物學方法分離鑒定腐乳中的乳酸菌。通過本研究的開展,可促進腐乳產業化生產,為大豆發酵食品領域提供一個重要的理論支持。
1 材料與方法
1.1 材料與試劑
樣品:采購于恩施市菜市場。
三羥甲基氨基甲烷(分析純)、乙酸(分析純),乙二胺四乙酸(分析純)、丙烯酰胺(分析純)、甲叉雙丙烯酰胺(分析純)、去離子甲酰胺(分析純)、(分析純)、過硫酸銨(分析純)、四甲基乙二(分析純)、乙醇(分析純)、冰醋酸(分析純)、甲醛(分析純)、硝酸銀(分析純)、氫氧化鈉(分析純)、MRS 合成培養基:國藥集團化學試劑有限公司;D5625-01 DNA 提取試劑盒、DNA marker、PCR 清潔試劑盒:北京科博匯智生物科技發展有限公司;2xPCR mix:南京諾唯贊生物科技有限公司;rTaq、dNTP mix、pMD18-T vector:大連寶生物技術有限公司;正向引物338F(加入7 個核苷酸標簽barcodes)和反向引物806R、PCR 引物合成和測序:武漢天一輝遠生物科技有限公司。
1.2 儀器與設備
VeritiTM 96-well thermal cycler PCR 儀、NanoDrop 2000/2000c:美國 Thermo Fisher 公司;DCodeTM System:美國Bio Red 公司;DYY-12 電泳儀:北京六一儀器廠;Miseq PE300 高通量測序平臺:美國Illumina 公司;R920 機架式服務器:美國 DELL 公司;CT15RE 冷凍離心機:日本HITACHI 公司;Bio-5000 plus 掃描儀:上海中晶科技有限公司;Whitley DG250 厭氧工作站:英國DWS 公司。
1.3 試驗方法
1.3.1 樣品宏基因組提取與檢測
采用試劑盒方法提取腐乳樣品中的宏基因組,用0.8%瓊脂糖凝膠進行電泳檢測,測定各樣品宏基因組DNA 濃度。
1.3.2 PCR-DGGE
將宏基因組DNA 濃度調整為一致后作為模板細菌16S rRNA V3 區域基因片段PCR 擴增。采用25 μL體系進行 PCR 擴增:10×PCR Buffer(含 Mg2+)2.5 μL,dNTP 2 μL,上下游引物各 0.5 μL,rTaq 0.5 μL,模板1 μL,滅菌超純水補充至25 μL。其中上游引物為ALL-GC-V3F(5’-CGCCCGGGGCGCGCCCCGGGCG GCCCGGGGGCACCGGGGGCCTACGGGAGGCAGCAG-3’),下游引物為 ALL-V3R(5’-ATTACCGCGGCTGCTGG-3’)。擴增程序:95 ℃ 4 min,95 ℃ 30 s,55 ℃30 s,72 ℃ 30 s,30 個循環,72 ℃ 10 min。擴增結束后,PCR 擴增產物用2%的瓊脂糖凝膠電泳檢測。
采用8%的聚丙烯酰胺(丙烯酰胺∶甲叉雙丙烯酰胺=38.93 ∶1.07,質量比)、變性范圍為35%~52%(100%變性劑:尿素42 g,去離子甲酰胺40 mL)的凝膠進行分析。將凝膠置于溫度為60 ℃的0.5 x TAE 電泳緩沖液中,于每個膠孔點樣10 μL,先電壓120 V,持續80 min 后,電壓80 V,持續13 h。電泳結束后,采用硝酸銀法染色,使用掃描儀對電泳圖進行觀察拍照,找出各泳道優勢條帶并切膠,將膠塊搗碎于50 μL 無菌超純水中,4 ℃靜置過夜。用不含GC 夾的引物(ALL-V3F 和ALL-V3R)將回收膠塊進行PCR 擴增,擴增體系及條件同DGGE 擴增。用清潔試劑盒純化PCR 產物,并與載體(PMD18-T)連接后轉化到感受態細胞中進行克隆培養,篩選陽性克隆進行測序。使用BioEdit 軟件將去除載體序列后在NCBI 中進行同源性比對。
1.3.4 樣品細菌16S rRNA PCR 擴增及Miseq 高通量測序
參考王玉榮等[15]方法進行樣品細菌16S rRNA PCR擴增及Miseq 高通量測序。采用20 μL 擴增體系:5×PCR 緩沖液 4 μL,dNTP mix 2 μL,上游引物 338F(5’-ACTCCTACGGGAGGCAGCA-3’)和下游引物 806R(5’-GGACTACHVGGGTWTCTAAT-3’)各 0.8 μL,rTaq 酶0.4 μL,模板 10 ng,滅菌超純水補充至 20 μL。擴增條件為:95 ℃預變性 3 min,95 ℃變性 30 s,55 ℃退火 30 s,72 ℃延伸 45 s,共運行 30 個循環,72 ℃延伸 10 min。擴增結束后用1.0%瓊脂糖凝膠電泳檢測PCR 擴增產物,合格后進行高通量測序。
1.3.5 序列拼接及質量控制
參照于丹等[16]和陳澤斌等[17]的方法,在除去不合格序列、barcode 序列和引物序列的基礎上,將數據序列進行拼接。同時利用PyNAST 軟件將所有的序列對齊,采用UCLUST 算法將相似度>97%序列劃分為一個操作單元(operational taxonomic units,OTU),從而進行物種鑒定和相對含量分析,對腐乳中的微生物多樣性進行解析。
1.3.6 腐乳中乳酸菌的分離與鑒定
腐乳樣品中乳酸菌的分離采用倍比稀釋涂布法。將各樣品稀釋液涂布于改良MRS 固體培養基(含1.5%CaCO3)上,置于37 ℃厭氧培養48 h,挑選具有不同特征且透明圈現象明顯的菌落劃線純化。純化后的菌株進行革蘭氏染色鏡檢、過氧化氫試驗及凍存。參考文獻[18]提取各菌株DNA,并參照張曉輝等[19]方法使用通用正向引物 27F(5’-AGAGTTTGATCCTGGCTCAG-3’)和反向引物 1495R(5’-CTACGGCTACCTTGTTACGA-3’)進行PCR 擴增和測序。測序結果分析同1.3.3。
1.4 數據處理
通過Origin 8.5 軟件對DGGE 圖譜特征條帶序列進行統計,同時對樣品的稀釋曲線及香農指數曲線(shannon diversity index curve)的作圖。系統發育樹由BioEdit 軟件和MEGA 5.0 軟件共同繪制。使用Office 2016 繪制平均相對含量>5%的屬水平餅圖。Venn 圖由在線繪圖網頁(http://bioinfogp.cnb.csic.es/tools/venny/index.html)進行繪制。相對含量>1.5%的核心OTU熱圖由Matlab 2010b 繪制。
2 結果與分析
2.1 腐乳中細菌DGGE圖譜及分析
本研究首先使用變性梯度凝膠電泳技術對樣品16SrRNAV3 區域細菌群落組成進行研究,如圖1所示。
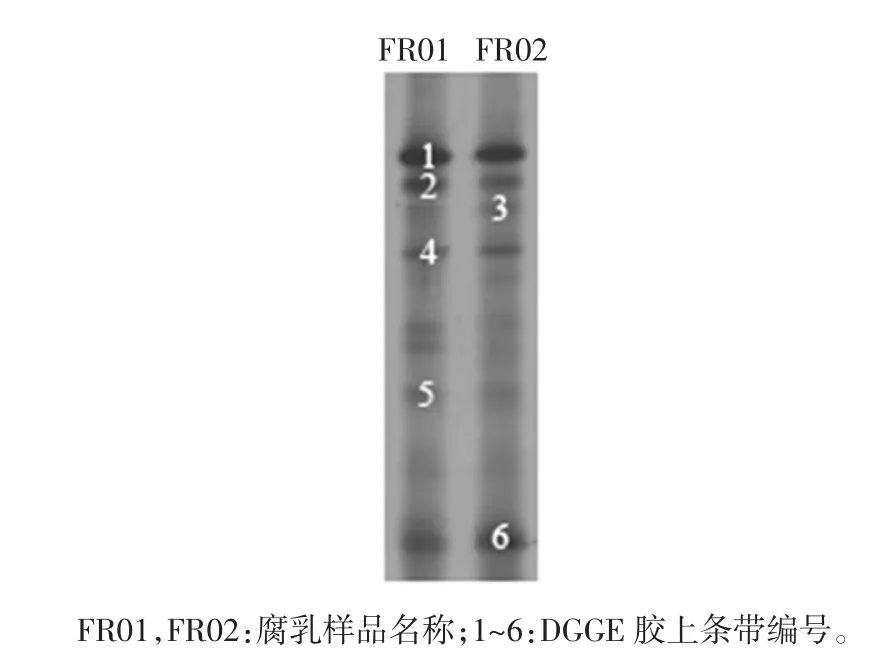
圖1 腐乳細菌PCR-DGGE 圖譜Fig.1 PCR-DGGE analysis of bacteria in sufu
由圖1可知,圖譜中共出現6 條明亮的條帶,同時條帶 1、2、4、5 和 6 是兩個樣品的共有條帶,說明不同腐乳樣品中存在一些共有的細菌種群,其中條帶1 最亮且存在于兩個樣品中,說明此條帶所對應的細菌在腐乳中發揮著重要的作用。值得一提的是各條帶亮度不同,條帶2 和條帶5 在FR01 中亮度較高,條帶6 在FR02 中亮度較高,表明在細菌種群豐富度存在差異。而條帶3 僅存在FR02 樣品中,說明不同腐乳樣品中存在著不同的細菌菌群。進一步將各條帶進行序列分析,結果如表1所示。

表1 腐乳細菌DGGE 比對結果Table 1 Blast results of bacteria DGGE in sufu
由表1可知,各條帶序列與數據庫中16S rRNA序列均具有較高的相似度。其中條帶1 和2 為植物乳桿菌(Lactobacillus plantarum),條帶3 為嗜鹽芽孢桿菌屬細菌(Halobacillus karajiensis),條帶 4 為不動桿菌屬細菌(Acinetobacter oryzae),條帶5 為熒光假單胞桿菌(Pseudomonas fluorescens),條帶6 為嗜鹽四聯球菌(Tetragenococcus halophilus)。由此可知,腐乳樣品中微生物構成具有多樣性,而且隸屬于乳桿菌屬(Lactobacillus)的植物乳桿菌(Lactobacillus plantarum)為優勢細菌。陳穎慧[20]利用PCR-DGGE 技術對4 種不同品牌腐乳中的細菌多樣性進行了研究,結果發現乳酸菌屬、藤黃微球菌(Micrococcus luteus)和屎腸球菌(Enterococcus Faecium)在各腐乳樣品中均存在,進一步發現乳酸桿菌屬(Lactobacillus)為優勢細菌。陳浩等[21]利用構建16S rRNA 基因文庫的方法對豆醬樣品進行研究, 結果發現嗜鹽四聯球菌(Tetragenococcus halophilus)為優勢細菌,同時不動桿菌(Acinetobacter baylyi)也被檢測到存在于樣品中。王夫杰等[22]在青方腐乳中分離出植物乳桿菌(Lactobacillus plantarum)和干酪乳桿菌(Lactobacillus casei)等乳酸菌。這與上述結論相一致。
本研究進一步將各條帶序列與模式菌株序列進行系統發育樹的構建,結果圖2所示。
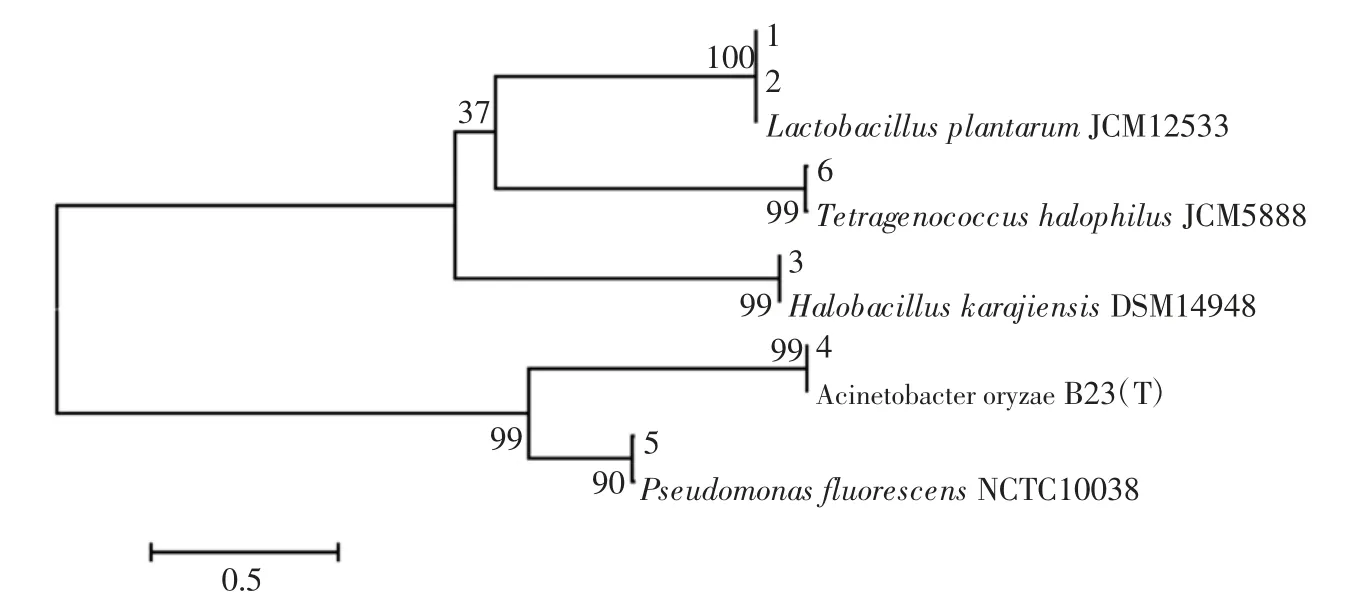
圖2 腐乳中細菌系統發育樹Fig.2 Phylogenetic tree of bacteria in sufu
由圖2可知,系統發育樹被分為2 大分支,其中條帶1 和2 與Lactobacillus plantarum 聚為一類,條帶6與Tetragenococcus halophilus 聚為一類,條帶 3 與Halobacillus karajiensis 聚為一類。而條帶4 和條帶5聚集在另外一支上,其中條帶4 和Acinetobacter oryzae聚為一類,條帶5 和Pseudomonas fluorescens 聚為一類。由此可知。不同腐乳樣品中細菌群落構成存在一定的差異性。
2.2 序列豐富度及多樣性分析
通過Miseq 高通量測序發現,兩個樣品共產生78 412 條高質量16S rRNA 序列。本研究采用兩步UCLUST 算法分別以100%和97%的相似度進行序列劃分并建立OTU,首先根據100%相似度進行序列劃分得到33 254 條序列,根據97%相似度進行OTU 劃分后得到2 317 個OTU,平均每個樣品1 158 個OTU。當樣品測序量為36 219 條序列時,FR02 樣品具有最大的細菌物種豐富度同時細菌多樣性最高,其Chao 1指數為611,Shannon 指數為5.64。進一步通過稀疏曲線和香農指數曲線圖對各樣品產生的數據量來判定是否滿足后續生物信息學分析,其結果如圖3所示。
由圖3A 可知,隨著測序量不斷的增加,各樣品被發現OTU 的數量也隨之增加,而由圖3B 可知,當序列數達到10 000 條時,各樣品的香農多樣性曲線已處于飽和狀態,由此可知隨著測序序列數的增加,盡管會有新的細菌種系型出現,但其多樣性不再發生變化,可以反映樣品中絕大多數微生物物種信息。因而本研究中每個樣品產生的序列數是可以將樣品中細菌微生物多樣性表現出來,同時可以滿足后續生物信息學分析需求。
2.3 基于不同分類地位腐乳樣品核心細菌菌群相對含量分析
納入本研究的序列被鑒定為14 個門,24 個綱,53 個目,84 個科,145 個屬,其中只有5.3%的序列不能鑒定到屬水平。研究發現腐乳樣品中平均相對含量>1%的細菌門為變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)和硬壁菌門(Firmicutes),其含量分別為80.45%、10.7%和6.81%。同時在2 個樣品中隸屬于變形菌門(Proteobacteria)的細菌相對含量分別為77.29%和83.61%,隸屬于擬桿菌門(Bacteroidetes)的細菌相對含量為17.19%和4.22%,而隸屬于硬壁菌門(Firmicutes)的細菌相對平均含量為3.76%和9.87%。由此可知,在門水平上各樣品中的微生物存在差異,本試驗進一步對各樣品在屬水平上進行分析,如圖4所示。
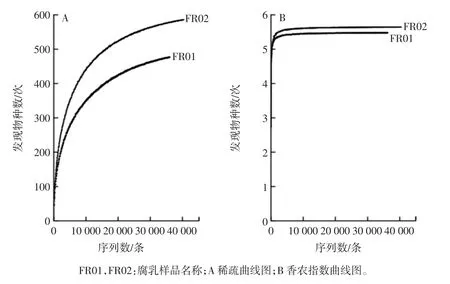
圖3 稀疏曲線圖和香農指數曲線圖Fig.3 Rarefaction curve and Shannon diversity index curve
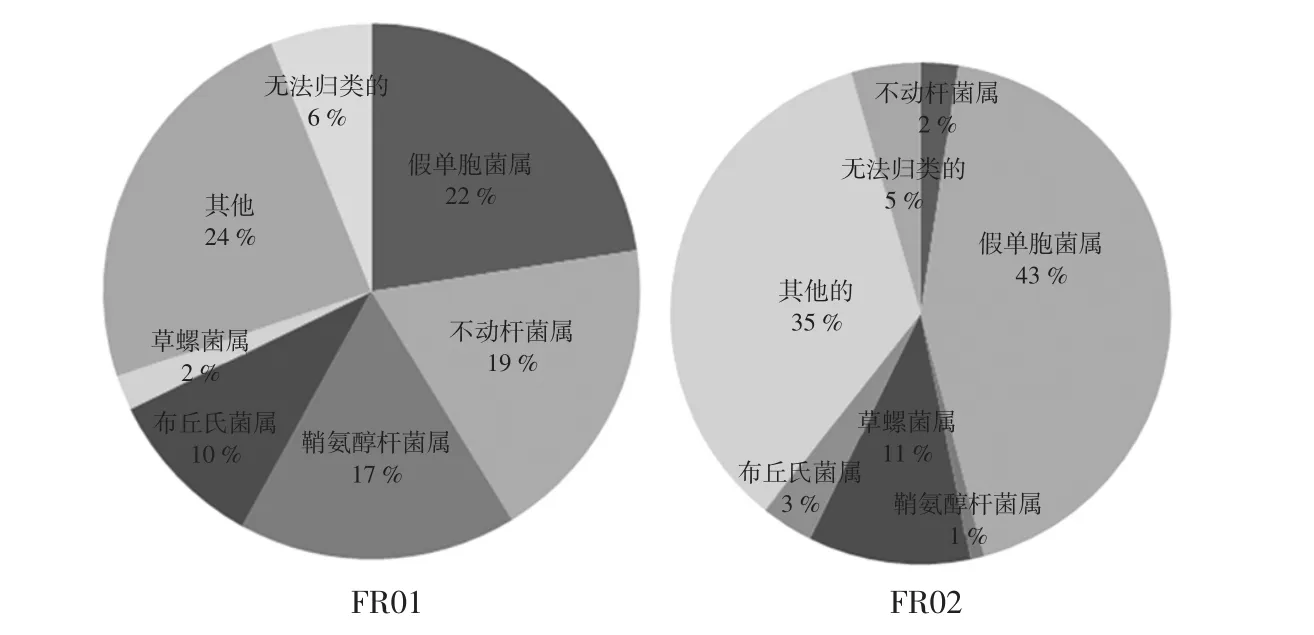
圖4 腐乳中優勢細菌屬平均相對含量比較分析Fig.4 Comparative analysis the dominant bacterial genera with average relative abundance in sufu
由圖4可知,腐乳樣品中平均相對含量>5%的細菌屬包括綠膿桿菌屬(Pseudomonas)、不動桿菌屬(Acinetobacter)、鞘氨醇桿菌屬(Sphingobacterium)、布丘氏菌屬(Buttiauxella)和草螺菌屬(Herbaspirillum),其平均相對含量分別為33%、10.56%、8.82%、6.57%和6.32%。然而各細菌屬在2 個樣品中的相對含量均存在很大差異,其中綠膿桿菌屬(Pseudomonas)的相對含量分別為22.56 %和43.46 %,同時不動桿菌屬(Acinetobacter)的相對含量分別為18.68%和2.43%,鞘氨醇桿菌屬(Sphingobacterium)的相對含量分別為16.77%和0.88%,這與PCR-DGGE 結果相一致。劉亞棟[23]利用16S rDNA 測序的方法對腐乳中的微生物多樣性進行鑒定分析,結果發現變形菌門(Proteobacteria)、硬壁菌門(Firmicutes)、放線菌門(Actinobacteria)和擬桿菌門(Bacteroidetes)為腐乳樣品中的優勢細菌門,進一步分析發現四聯球菌屬(Tetragenococcus)、鹽單胞菌屬(Halanaerobium)、Rummeliibacillus 屬和乳酸桿菌屬(Lactobacillus)、不動桿菌屬(Acinetobacter)和假單胞菌屬(Pseudomonas)均為腐乳匯樣品中的優勢屬,這與本文結論一致。
本試驗進一步統計了OTU 在兩個樣品中出現次數,如圖5所示。
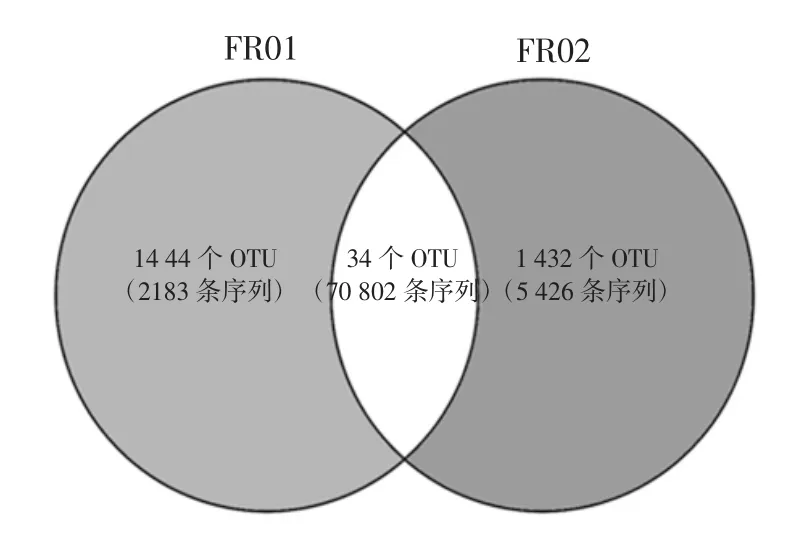
圖5 基于OTU 水平的Venn 圖Fig.5 Venn diagram based on OTU level
由圖5可知,在兩個腐乳樣品中出現1 次的OTU數量分別為1 144 個和1 432 個,分別占OTU 總數的43.8%和54.9%,序列數分別為2 183 條和5 426 條。同時核心OTU 共有34 個,占OTU 總數的1.3%,包含70 802 條序列。進一步分析發現有9 個核心OTU 的相對含量>1.5%,結果如圖6所示。

圖6 相對含量>1.5%的核心OTU 熱圖Fig.6 Heat map of the relative abundance more than 1.5%of cores OTUs
由圖6可知,OTU874 和 OTU1711 隸屬于綠膿桿菌屬(Pseudomonas),OTU1671 和 OTU894 隸屬于不動桿菌屬(Acinetobacter),OTU577 隸屬于沙雷菌屬(Serratia),OTU1973隸屬于短波單胞菌屬(Brevundimonas),OTU1907 隸屬于草螺菌屬(Herbaspirillum),OTU635 隸屬于鞘氨醇桿菌屬(Sphingobacterium),而OTU346 未鑒定在屬水平,只鑒定在Enterobacteriaceae。同時由圖6可以發現,OTU874、OTU635 和 OTU1671 在FR01 中含量較高,其相對含量分別為16.89 %、16.73 %和 11.32 %,同時 OTU874 和 OTU1907 在FR02 中含量較高,其相對含量分別為28.56%和11.85%。
2.4 腐乳中乳酸菌分離鑒定結果及系統發育分析
通過傳統微生物培養手段結合16S rDNA 測序方法對腐乳樣品中的乳酸菌進行分離與鑒定,其鑒定結果和數據庫中模式菌的系統發育樹如圖7所示。
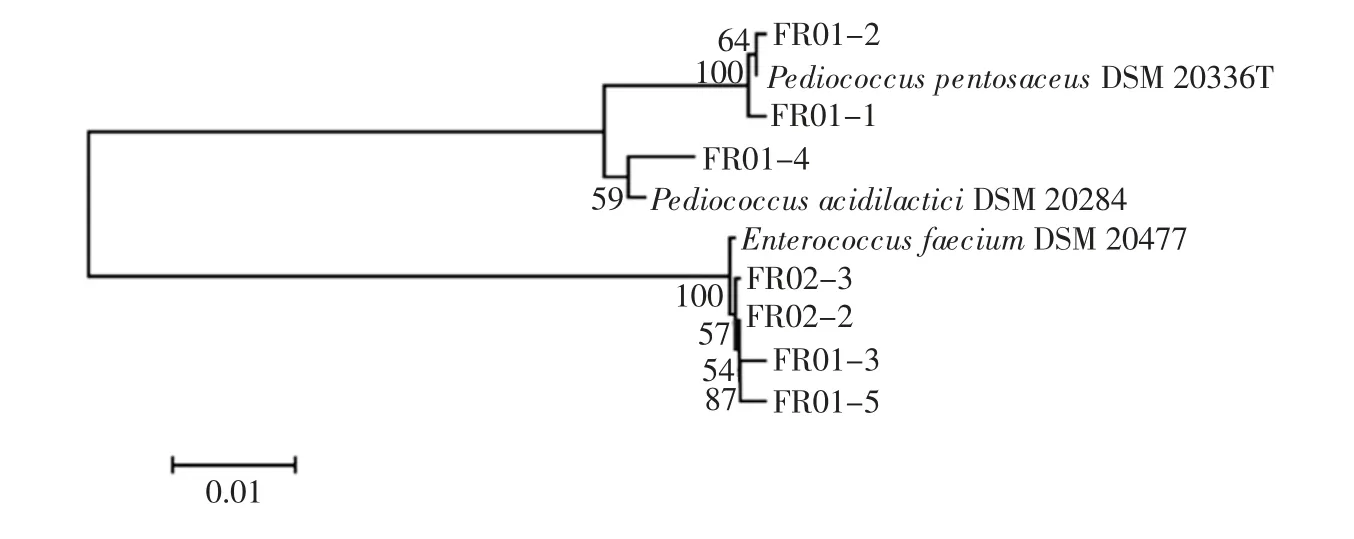
圖7 腐乳中乳酸菌系統發育樹Fig.7 Phylogenetic tree of Lactobacillus in sufu
由圖7可知,在腐乳樣品中共分離出7 株乳酸菌,其中2 株為戊糖片球菌(Pediococcus pentosaceus),1株為乳酸片球菌(Pediococcus acidilactici),4 株為屎腸球菌(Enterococcus faecium)。由此可知,腐乳中乳酸菌種類具有多樣性。
3 結論
本文使用PCR-DGGE 技術和Illumina Miseq 第二代高通量測序技術相結合的手段對恩施地區腐乳中的微生物群落組成及多樣性進行解析,同時利用傳統純培養的方法對其乳酸菌資源進行發掘。結果表明:隸屬于變形菌門的綠膿桿菌屬、不動桿菌屬、鞘氨醇桿菌屬、布丘氏菌屬和草螺菌屬為腐乳樣品中的優勢細菌屬,同時PCR-DGGE 技術與傳統微生物培養方法顯示腐乳中的微生物資源較為豐富且具有多樣性。通過本研究的開展,可為傳統大豆發酵食品提供優秀的菌種資源,同時更好地促進其產業化生產。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
