遮陰對玉帶草光合特性的影響
2019-08-21 01:13:57安佰義范愛淇吳雙
江蘇農業科學 2019年12期
安佰義 范愛淇 吳雙


摘要:以分株繁殖自然生長的玉帶草(Phalaris arundinacea var. Picta)為試驗材料,在30%、70%、90%遮陰度下處理 50 d 后,以全光照對其進行光合特性研究。結果表明,不同遮陰條件下的玉帶草葉片光合日變化曲線均為單峰型。隨遮陰程度的增加,玉帶草葉片的最大凈光合速率(Pnmax)、日均凈光合速率(Pn)、水分利用率(WUE)逐漸減小,遮陰度達90%時達最小值;30%與70%遮陰度下的玉帶草葉片光合光響應曲線與表觀量子效率(AQY)均高于對照,90%遮陰時低于對照。氣孔導度(Gs)、蒸騰速度均隨遮陰程度增加而先增后降,90%遮陰度時為最小值;70%遮陰度處理的CO2響應曲線與羧化效率(CE)高于對照,其他處理均低于對照。遮陰處理降低了葉片的CO2補償點和暗呼吸速率(Rd);遮陰處理提高了玉帶草葉片的葉綠素的含量,且70%遮陰度處理的葉片葉綠素a、葉綠素b與類胡蘿卜素含量較高。
關鍵詞:玉帶草;遮陰;光合特性
中圖分類號: S184? 文獻標志碼: A? 文章編號:1002-1302(2019)12-0157-04
玉帶草(Phalaris arundinacea var. picta),又名絲帶草,為禾本科草屬多年生草本植物,分布于我國東北、華北、華中及歐洲和北美洲部分地區。生性喜光耐寒。近年來玉帶草作為觀賞草在園林綠化中的應用逐漸增多,可以布置路邊花鏡或花壇鑲邊,主要用作水景園背景材料,也可點綴于橋、亭、榭四周,可盆栽用于庭院觀賞[1]。
目前,國內對玉帶草的研究僅限于光合特性、耐鹽性、耐旱性、解剖結構觀察以及花鏡選材等方面[2-4],而對玉帶草遮陰下的光合特性研究未見報道。本試驗研究了玉帶草在吉林長春地區的引種栽培過程中不同光照條件對其光合特性的影響,旨在明確玉帶草對光環境的適應性,以明確遮陰環境對玉帶草光合生理性狀的影響,以及玉帶草葉片適陰性的變化特點,并判斷蔭蔽的環境是否利于玉帶草的生存生長,進而為園林設計中喬、灌、草多層次景觀結構以及林陰植物種的選擇提供合理的依據。
光是植物光合作用所必需的環境因子。近年來,隨著城市的快速發展,高大建筑物及高層建筑設施迅速增多,城市園林綠地面積越來越小,城市弱光環境限制了植物的生存和生長。導致喜光的玉帶草不得不脫離原有的最適合生境,光成了影響玉帶草生長發育的重要環境因子之一。
1 材料與方法
1.1 試驗地概況和樣地描述
試驗在吉林省長春市吉林農業大學園藝學院園林實驗基地內進行。該試驗地位于125°24′~125°36′E、43°47′~43°61′N,海拔為251 m,為大陸性季風氣候;年均氣溫為 4.9 ℃,年均降水量為565 mm,年日照時間為2 698 h。
1.2 試驗設計和樣本采集
供試材料為2014年6月分株繁殖自然生長的玉帶草植株,培養土為園土,pH值為7.3。于2016年5月16日對生長良好的玉帶草進行遮光處理。試驗設4個遮陰度處理(用TES-1335型數位式照度計測定各處理的透光率):0(CK)、30%、70%、90%,用不同遮光度的遮陰網搭建遮陰棚(長 3.8 m×寬1.6 m×高1.5 m),遮陰棚南北朝向,2個遮陰棚間隔3 m,以避免交叉遮光。遮陰處理期間進行正常水分管理。處理50 d后測定玉帶草光合響應參數及色素含量。
1.3 試驗方法
2016年7月5—8日均為晴朗無云天氣,采用美國PP-Systems公司生產的CIRAS-2型光合測定系統測定玉帶草的凈光合速率(Pn)、氣孔導度(Gs)、細胞間隙CO2濃度(Ci)等光合參數。
1.3.1 光合日變化的測定 2016年7月5日,晴朗無云。于6:00~18:00進行凈光合速率的日變化測定,每隔 2 h 測定1次。測定指標包括凈光合速率、氣孔導度、細胞間隙CO2濃度、蒸騰速率,計算水分利用效率(WUE)=Pn/Tr;氣孔限制值(Ls)按照Berry等的方法計算[5],即Ls=1-Ci/Ca(Ca為大氣CO2濃度);表觀量子效率(AQY)=Pn/光合有效輻射(PAR)。測定時每處理選3 株,為3次重復。
1.3.2 光響應曲線的測定 2016年7月6日,晴朗無云。在09:00—11:30間,控制CIRAS-2型光合測定系統的CO2濃度為380 μmol/mol,溫度控制在(25±1) ℃,用發光=極管(LED)紅藍光源控制光合有效輻射為0~2 000 μmol/(m2·s),并測定玉帶草葉片凈光合速率。設置14個光合有效輻射梯度,分別為 2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、300、200、100、50、0 μmol(m2·s)。制作Pn-PAR響應曲線,在每個光照度下適應 3 min 后測定。Pn-PAR響應曲線由直角雙曲線修正模型擬合[6],決定系數均達到0.96以上,擬合度良好。根據擬合模型得出表觀量子效率等。
1.3.3 CO2響應曲線的測定 2016年7月7—8日,在 09:00—11:30晴朗無風時,測定不同處理組內健康成熟葉片的光合特征曲線,每個測定重復3次。在設定1 200 μmol/(m2·s)的光照度下測定在0、50、100、150、200、300、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol/mol CO2濃度條件下的Pn和Ci,制作Pn-Ci響應曲線,在每個CO2濃度下適應3 min后測定。Pn-Ci響應曲線均由直角雙曲線修正模型擬合[7],決定系數均在0.96以上,擬合度良好。根據擬合模型得出羧化效率(CE)等。
1.3.4 色素含量的測定 從進行光合測定的每株植物上采集1~5張完全展開的健康葉片,參考蘇正淑等的乙醇丙酮混合液法進行光合色素含量的測定[8]。
1.4 數據處理
試驗數據采用Excel 2010和SPSS 18.0軟件進行數據統計與分析。
2 結果與分析
2.1 遮陰處理對玉帶草葉片光合日變化的影響
凈光合速率日變化常因植物種類和環境條件的不同而表現出差異,同一植物不同部位的葉片在不同月份、不同環境條件下可呈現不同的類型[9],通常在自然條件下,凈光合速率日變化可分為“雙峰”型、“單峰”型和“三峰”型。從圖1、表1可以看出,不同遮陰處理下玉帶草葉片的凈光合速率均表現出先增加后減少的單峰型變化規律,沒有光合午休現象,峰值隨遮陰程度加大而逐漸延后。最大凈光合速率(Pnmax)、日均Pn與WUE均隨著遮陰程度增加而減小。由于光能明顯降低,90%遮陰度的玉帶草日均Gs、Pn、WUE和Pnmax均為最小值,與其他3個處理間存在極顯著差異,表明玉帶草受重度遮陰影響最敏感。氣孔的行為直接影響大氣與表皮層之間的CO2和水汽的交換,氣孔阻力占葉片與大氣之間CO2和水汽交換總阻力相當大的部分,蒸騰作用是光合作用不可避免的結果,蒸騰流從根到蒸騰器官的流動使土壤中的營養物質和溶液運輸到根表[10]。90%遮陰度的日均Gs、Tr與WUE最小,可能是過度遮陰使玉帶草葉片的氣孔關閉,減少了蒸騰作用,使玉帶草根部從土壤中吸收的水分減少,從而也降低了水分利用效率。
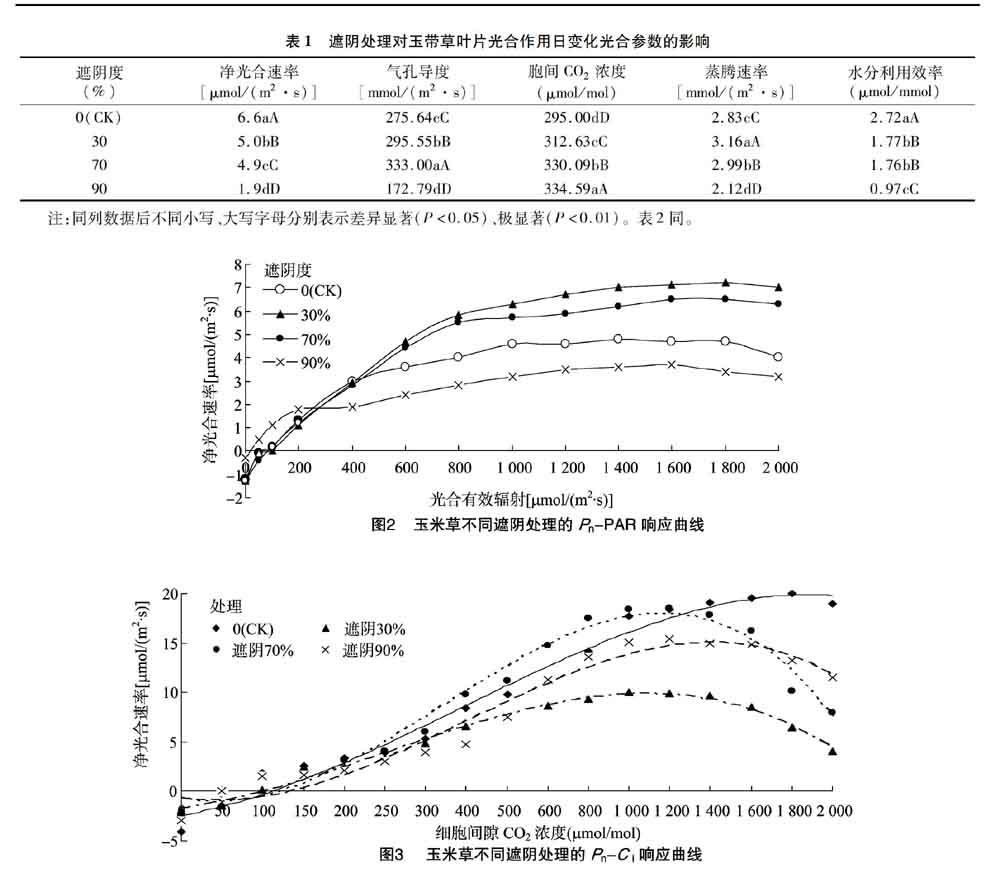
2.2 遮陰處理對玉帶草葉片光合光響應的影響
從圖2可以看出,30%遮陰度的Pn-PAR曲線整體高于CK,90%遮陰度的最低,但在初始階段,90%遮陰度的凈光合速率明顯高于其他3個處理,可能是由于重度遮陰下玉帶草葉片的應激反應。植物的光補償點越低,越能在弱光條件下順利進行光合作用,開始增加有機物質;而植物的飽和光照度越小則表明植物葉片的光合速率隨光合有效輻射的增加很快達到最大值,然后光合速率隨光合有效輻射的增加而降低[11-12]。玉帶草葉片光補償點在90%遮陰度環境下驟減,說明90%遮陰度下玉帶草對弱光的利用能力較強;WUE與Pn均隨遮陰程度的增加呈遞減趨勢,90%遮陰度下玉帶草葉片的Pn驟減,依據許大全的報道[13],遮陰環境下玉帶草葉片的Pn下降的原因之一是氣孔限制,Gs下降阻止了CO2的供應。表觀量子效率是光合作用中光能轉化最大效率的一種度量,可以正確反映光合機構機能的變化,也可以反映葉片對弱光的利用能力[14]。玉帶草葉片的AQY隨遮陰程度的增加呈現先上升后下降趨勢,70%遮陰度下AQY達最大值,為0.015 1,這與張哲等在60%透光下處理的白三葉的結果[15]相似,但遮陰處理的玉帶草葉片AQY均低于全光照處理,說明玉帶草葉片對弱光有一定的適應和利用能力。最大凈光合速率越大,植物葉片在遮陰脅迫下的光合潛力就越大。本試驗中不同處理的Pnmax分別為4.7、7.2、6.4、4.1 μmol/(m2·s),說明30%、70%遮陰度下玉帶草的光合潛力較大,但90%遮陰度影響玉帶草葉片的光合能力。
2.3 遮陰處理對玉帶草葉片光合CO2響應的影響
從圖3可以看出,70%遮陰度下的玉帶草葉片的Pn整體上高于對照,其他處理整體上均低于對照。暗呼吸是植物維持自身正常代謝而進行物質氧化分解獲得能量的過程,植物可以通過降低暗呼吸速率來適應低光環境[16]。玉帶草葉片暗呼吸速率(Rd)隨遮陰程度增加而減小,說明玉帶草能通過降低Rd來適應弱光環境。CO2是光合作用的原料,在一定濃度范圍內,其含量越高光合作用就越強。CO2濃度的升高,一方面可以增加CO2對核酮糖-1,5-二磷酸羧化酶結合位點的競爭,從而提高羧化效率;另一方面可以抑制光呼吸來提高光合效率[17]。70%遮陰度處理的玉帶草葉片的羧化效率(CE)最高,其他處理的玉帶草葉片的CE均低于對照,90%遮陰度處理的玉帶草葉片的CE明顯低于CK,說明70%遮陰度能提高玉帶草的碳同化能力,但重度遮陰對其影響很大。遮陰處理的玉帶草葉片CO2補償點均低于CK,70%遮陰度處理的玉帶草葉片CO2補償點最低。
2.4 遮陰處理對玉帶草葉片光合色素的影響
從表2可以看出,遮陰處理的玉帶草葉片葉綠素a、葉綠素a+b、與葉綠素b含量整體上隨遮陰程度的增加而增加,葉綠素a/b的變化規律則相反,這與宋洋等的研究結果[18]一致。其中,70%遮陰度的葉綠素a+b與類胡蘿卜素含量最高,均與CK差異極顯著。類胡蘿卜素所吸收的光傳遞給葉綠素后才能啟動光合作用的輔助性作用,亦有利于遮陰葉提高光能利用效率。遮陰處理的類胡蘿卜素含量均高于CK,其中70%遮陰度的類胡蘿卜素含量最高,表明70%遮陰度環境能提高玉帶草葉片的光能捕獲能力從而提高光能利用效率,但90%遮陰度處理會使其能力減弱。
3 討論與結論
光合作用日變化是植物生產過程中物質積累與生理代謝的基本單元,常因植物種類和環境條件的變化而有差異,它是分析環境因素影響植物生長和代謝的重要手段[19]。玉帶草的凈光合速率日變化均呈單峰型曲線,不存在光合午休現象。90%遮陰度使玉帶草的Pn峰值推遲到14:00,玉帶草對30%、70%的遮陰環境有一定的適應性,但90%遮陰度對玉帶草的生長代謝明顯造成傷害。
光響應曲線在一定程度上可以反映不同光照下植物光合速率的變化規律,還可直觀地表示遮陰對植物凈光合速率的抑制情況。30%、70%遮陰度有利于玉帶草葉片的光合作用,而90%遮陰度已是過度遮陰,抑制了玉帶草葉片的光合作用。
CO2濃度升高不僅為光合作用提供了較多的原料,而且提高了1,5-二磷酸核酮糖(RuBp)羧化酶的活性,增加了對CO2的固定能力;同時抑制了RuBP加氧酶的活性,減少了光合呼吸底物乙醇酸的生成,降低了植物的光呼吸強度,從而提高了光合作用效率[20]。30%、70%遮陰度的玉帶草葉片通過提高對CO2的固定能力,降低了呼吸強度,從而提高了凈光合速率,但90%的遮陰度則大大降低了玉帶草葉片的光合作用效率。
比較玉帶草的光合特征值與C4、C3陽性和陰性植物的光合生理生態特征值[21],玉帶草的光飽和點、CO2補償點與最大凈光合速率均在C3陰性植物的光合生理特征值范圍內,而光補償點則符合C4植物范圍。綜合判斷后,玉帶草更近似于C3陰性植物。
植物吸收光能主要是由植物的葉綠素完成的,所以,其含量的多少直接影響植物光合作用的強弱。葉綠素的重要功能是能選擇性地吸收光,葉綠素a在紅光部分的吸收帶偏向長紅波方面,葉綠素b則在藍紫光部分的吸收帶較寬[22]。本試驗中遮陰下的玉帶草葉片中葉綠素的含量均高于CK,說明遮陰環境有利于玉帶草葉片葉綠素的合成與積累,其中70%遮陰度處理的玉帶草葉綠素含量最高,葉色更具觀賞性,表明70%遮陰度的效果最好。
參考文獻:
[1]張相偉. 天津市花境植物材料選擇及設計研究[D]. 天津:天津大學,2013.
[2]劉曉嘉,范愛淇,安佰義,等. 玉帶草光合特性的研究[J]. 湖北農業科學,2017,56(15):2893-2896,2901.
[3]李洋洋. 水分脅迫對玉帶草生理特性的影響[D]. 哈爾濱:東北林業大學,2012.
[4]劉曉東,李洋洋,何 淼. PEG模擬干旱脅迫對玉帶草生理特性的影響[J]. 草業科學,2012,29(5):687-693.
[5]葉子飄,于 強. 一個光合作用光響應新模型與傳統模型的比較[J]. 沈陽農業大學學報,2007,38(6):771-775.
[6]葉子飄,于 強. 光合作用對胞間和大氣CO2響應曲線的比較[J]. 生態學雜志,2009,28(11):2233-2238.
[7]Berry J A,Downton W J S. Environmental regulation of photosynthesis[J]. Photosynthesis,1982(2):263-343.
[8]蘇正淑,張憲政. 幾種測定植物葉綠素含量的方法比較[J]. 植物生理學通訊,1989(5):77-81.
[9]郭寶林,楊俊霞,魯韌強,等. 遮光處理對扶芳藤生長和光和特性的影響[J]. 園藝學報,2007,34(4):1033-1036.
[10]黃振英,董學軍,蔣高明,等. 沙柳光合作用和蒸騰作用日動態變化的初步研究[J]. 西北植物學報,2002,22(4):93-99.
[11]彭曉邦,蔡 靖,姜在民,等. 渭北黃土區農林復合系統中大豆辣椒的光合生理特性[J]. 生態學報,2009,29(6):3173-3180.
[12]孫 穎. 百子蓮繁殖生物學研究[D]. 哈爾濱:東北林業大學,2009.
[13]許大全. 光合作用氣孔限制分析中的一些問題[J]. 植物生理學通訊,1997,33(4):241-244.
[14]張永霞,李國旗,張 琦,等. 不同遮陰條件下羅布麻光合特性的初步研究[J]. 西北植物學報,2007,27(12):2555-2558.
[15]張 哲,黃淑萍,杜桂娟,等. 遮陰對4種豆科牧草光合特性的影響[J]. 草業科學,2013,30(1):44-51.
[16]Garnier E,Cortez J,Billès G,et al. Plant functional markers capture ecosystem properties during secondary succession[J]. Ecology,2004,85(9):2630-2637.
[17]朱萬澤,王金錫,薛建輝,等. 四川榿木光合生理特性研究[J]. 西南林學院學報,2001,21(4):196-204.
[18]宋 洋,廖 亮,劉 濤,等. 不同遮陰水平下香榧苗期光合作用及氮分配的響應機制[J]. 林業科學,2016,52(5):55-63.
[19]孫 磊,章 鐵,李宏開,等. 柿樹光合速率日變化及其影響因子的研究[J]. 北方果樹,2006(2):4-6.
[20]陳翠琴,呂洪飛,黃四娣,等. 三白草科2種植物光合作用和葉綠素熒光特性的比較研究[J]. 浙江農業學報,2011,23(4):725-730.
[21]杜占池,楊宗貴. 冰草葉片光合速率與生態因子的關系[J]. 草地學報,2000,8(3):155-163.
[22]Boardman N K. Comparative photosynthesis of sun and shade plants[J]. Annual Review of Plant Physiology,1977,28(1):355-377.王薇薇,祖艷俠,吳永成,等. 鹽脅迫對豇豆幼苗離子分布的影響[J].
