5-羥色胺和多巴胺在初孵揚子鱷小腦的定位與表達
2019-08-21 08:31:54任士佳陳梅芳劉再群周際海
生物學雜志 2019年4期
任士佳, 陳梅芳, 劉再群, 周際海
(安徽師范大學 生命科學學院 安徽省重要生物資源與利用重點實驗室 生物環境與生態安全省級重點實驗室, 蕪湖 241000)
5-羥色胺(5-hydroxytryptamine, 5-HT),因其最先是從人的血清素中發現的,具有緊縮血管的功能,故又稱血清素(serotonin)。5-HT是一種吲哚衍生物,分子式為C10H12N2O。現已證實,5-HT作為一種“經典”的神經遞質,普遍分布于脊椎動物的中樞神經系統中,參與介導各類活動諸如學習、記憶、痛覺傳導、睡眠、攝食、行為等的調節。另有研究表明,5-HT在變色龍種群間的信息交流過程中也發揮作用[1]。目前,在各類爬行動物如暹羅鱷[2]、尼羅鱷[3]、日本水龜[4]、平原巨蜥[5]、毒蝰[6]的中樞神經系統已有一些關于5-HT分布的報道。有學者認為暹羅鱷小腦內5-HT含量及濃度并不高,但幼體要高于成體[2]。在尼羅鱷小腦內可見小腦深核的5-HT免疫陽性[3]。在日本水龜的小腦,5-HT能纖維可見于顆粒層,但未見于分子層,且小腦核周圍也可見一些5-HT能纖維[4]。在平原巨蜥小腦核旁側可見5-HT免疫陽性神經細胞,分子層、顆粒層以及小腦核附近也可見5-HT免疫陽性神經纖維[5]。在毒蝰小腦皮質的顆粒層有大量的5-HT能纖維[6]。
多巴胺(dopamine, DA)也是一種“經典”的神經遞質,分子式為C8H11O2N。在之前的很長一段時間,人們一直以為多巴胺在中樞神經系統中僅僅是作為NE的前體物質發揮作用,直到1975年瑞典科學家Carlsson確定DA在腦內還扮演了信息傳遞者的角色,人們才意識到DA在神經信號傳導過程中也發揮著重要作用。現已證明,DA在運動控制、聯系學習、覺醒、執行能力等方面都發揮了重要的作用。人腦DA的缺乏與一些神經精神疾病,如精神分裂癥、帕金森癥等精神疾病的病發有著非常密切的干系[7]。目前對于DA在爬行動物如紅耳龜[8]、彩龜[9]、球蟒[10]、大壁虎[11]的中樞神經系統的分布已有一些相關研究。尚未見DA在鱷類動物中樞神經系統的相關報道。
揚子鱷(Alligatorsinensis)屬爬行綱鱷目短吻鱷科短吻鱷屬,是中國特有的一種珍稀淡水鱷,為國家一級保護動物,野生種群數量較少,主要分布于長江流域。它們體型較大、結構較進步,被譽為爬行動物中的“活化石”。現在,對揚子鱷的研究主要集中在揚子鱷的消化系統、呼吸系統和運動系統的組織學研究以及揚子鱷胚胎學、生態學、人工飼養和繁殖、保護和利用等方面[12],但關于揚子鱷中樞神經系統的研究并不是很多。對于揚子鱷小腦的報道,僅見于劉再群等[13]對初孵揚子鱷小腦皮質生長抑素的免疫組化研究。
本文運用Nissl染色方法研究了初孵揚子鱷小腦的組織學結構,運用免疫組織化學方法研究了5-HT和DA在初孵揚子鱷小腦的表達和定位情況,以期為日后更深層的研究提供一些理論基礎。
1 材料與方法
1.1 材料
初孵揚子鱷3只,長約15 cm,取自安徽省宣城市揚子鱷繁殖研究中心。乙醚麻醉后迅速從顱腔剝取整腦組織,腦重約0.24~0.36 g。上述行為均得到了中國林業部門和安徽師范大學動物倫理委員會的批準。用Bouin′s液固定48 h后,常規脫水、透明,石蠟包埋,萊卡切片機連續切片(厚約7 μm),于45 ℃清水展片并固定于含粘片劑的潔凈載玻片上,然后于37 ℃烘箱內烘干。
1.2 Nissl染色方法
干燥后的切片每隔5張間距選取一張,將切片放入二甲苯中脫蠟后,100% (I)、100% (II)、95% (I)、95% (II)、80%和70%梯度酒精和蒸餾水復水,再將切片置于甲苯胺藍染液中染色10 min,隨后置于0.25%冰乙酸分色3 s,蒸餾水清洗多余液體,放入烘箱中烘干后于二甲苯中透明,最后用中性樹膠進行封片。
1.3 免疫組織化學染色方法
SABC法:切片入二甲苯(I)和二甲苯(II)脫蠟,再分別入100% (I)、100% (II)、95% (I)、95% (II)、80%和70%酒精以及蒸餾水復水,30% H2O2和蒸餾水1∶ 9混合后滴加于組織上,室溫下放置5~10 min以滅活內源性酶,然后用蒸餾水沖洗3次;將切片浸入枸櫞酸鹽緩沖液(pH 6.0)熱修復抗原;冷卻至室溫后再用PBS(pH 7.2~7.6)洗2 min×3次;滴加5% BSA封閉液,室溫下封閉20 min,甩去多余的液體(勿洗);滴加一抗(5-HT, DA;1∶ 100稀釋),4 ℃冰箱中過夜。次日,PBS連續沖洗3次,每次2 min;滴加生物素化二抗,37 ℃恒溫箱中放置30 min后取出,PBS連續洗2 min×3次;再滴加SABC,37 ℃恒溫箱中放置30 min,PBS洗5 min×4次。DAB(取1 mL蒸餾水,往其中各加一滴A、B、C混勻)顯色,室溫下放置5~30 min,顯微鏡下控制反應時間,蒸餾水沖洗;蘇木素輕度復染,切片入梯度酒精脫水,再入二甲苯透明,最后用水溶性封片劑封片。抗兔5-HT多克隆抗體(BA-0121-1)、抗兔DA多克隆抗體(BA-1454)、鏈霉素親和素-生物素-過氧化物酶復合物(Streptavidin-Biotin-Peroxidase Complex,SABC)免疫組織化學試劑盒(SA 1029),均購自武漢博士德生物工程有限公司。
1.4 觀察
在Olympus BX61型顯微鏡和Motic BA600-4型全自動掃描顯微鏡下觀察、拍照。
2 結果
2.1 初孵揚子鱷小腦
初孵揚子鱷中樞神經系統長約4.5 cm,從前往后逐次可分成端腦、間腦、中腦、小腦、延髓和脊髓,通常將中腦和延髓合稱為腦干(圖 1)。初孵揚子鱷小腦位于中腦之后,延腦的背前側(圖 1-A、C),呈橢圓形,且橫徑大于縱徑,其與腦干共同圍成第四腦室(the fourth ventricle, V4)。

A和a: 背面觀; B和b: 腹面觀; C和c: 側面觀。 1: 嗅球; 2: 大腦; 3: 中腦; 4: 小腦; 5: 延髓; 6: 脊髓; 7: 漏斗
圖1初孵揚子鱷中樞神經系統
Figure 1 The central nervous system of the new-hatching Yangtze alligator
橫切面上,可見小腦尾端嵌入腦干上端,圍成第四腦室背部(圖 2-A)。往頭端,小腦在第四腦室背部逐漸變寬(圖 2-B),兩側與下面的腦干融合(圖 2-C)。再往頭端,小腦繼續變寬,且小腦中部嵌入第四腦室(圖 2-D);背部的小腦與腹部的腦干圍成的第四腦室逐漸變大,第四腦室的正中溝和外側溝明顯可見(圖 2-E、F),再繼續往頭端,小腦與腦干分離,第四腦室逐漸變小且僅被腦干封閉(圖 2-G、H、I)。縱切面上,可見小腦室(圖 2-J),小腦與腦干形成V4并通過脈絡叢來封閉V4,封閉不完全(圖 2-J)。
小腦皮質表面并不光滑,有皺褶,可見溝回分化(圖 3-A)。在小腦髓質內,埋藏了兩個小腦核團,分別是外側小腦核(the nucleus cerebelli lateralis, NCL)(圖 3-B)和內側小腦核(the nucleus cerebelli medialis, NCM)(圖 3-C)。外側小腦核細胞分布稀疏且胞體較小;內側小腦核細胞分布較密集且胞體較大。還觀察到在小腦皮質外有與其相聯系的血管叢(vascular plexus, VP)(圖3-D)。此外,髓質內還可見大量的血管(圖 3-E)。

A: 橫切面上小腦尾端嵌入腦干上端,圍成第四腦室(V4)背部; B: 橫切面上小腦在第四腦室背部逐漸變寬; C: 橫切面上小腦兩側與下面的腦干融合; D: 橫切面上小腦繼續變寬且小腦中部嵌入第四腦室; E、F: 背部的小腦與腹部的腦干圍成的第四腦室逐漸變大; G-I: 小腦與腦干分離,第四腦室逐漸變小且僅被腦干封閉; J: 縱切面上小腦與腦干形成V4并通過脈絡叢(CP)來封閉V4(插圖示脈絡叢)。標尺: 500 μm; 插圖標尺: 100 μm。VC: 小腦室
圖2初孵揚子鱷小腦(CBe)同腦干(BS)的相對位置
Figure 2 The relative position between the cerebellum (CBe) and brainstem (BS) of the new-hatching Yangtze alligator
2.2 5-HT在初孵揚子鱷小腦的分布
冠狀切面上,小腦皮質可明顯分為4層,從外向內為外顆粒層(external granular layer, EGL)、分子層(molecular layer, ML)、浦肯野細胞層(Purkinje cell layer, PCL)和顆粒層(granular layer, GL)(圖 4-A、B、C)。其中浦肯野細胞可見明顯多細胞層(圖 4-C)。髓質可見小腦核(圖 4-D)。矢狀切面上,可見第四腦室由頂部的小腦和下部的腦干圍成,第四腦室后部由脈絡叢(choroid plexus, CP)不緊密封閉(圖 5-A),構成第四腦室頂部的小腦皮質部位可見明顯的外顆粒層(圖 5-B),小腦髓質內部可見小腦室(ventriculus cerebelli, VC)(圖 5-D)。

矢狀切面上A: 小腦皮質表面并不光滑(標尺: 200 μm); B: 外側小腦核(標尺: 20 μm); C: 內側小腦核(標尺: 20 μm); D: 小腦皮質外血管叢(VP,標尺: 50 μm); E: 小腦髓質內的血管(箭頭示,標尺:50 μm)。GL: 顆粒層; ML: 分子層; PCL: 浦肯野細胞層
圖3初孵揚子鱷小腦結構
Figure 3 The cerebellar structure of the new-hatching Yangtze alligator
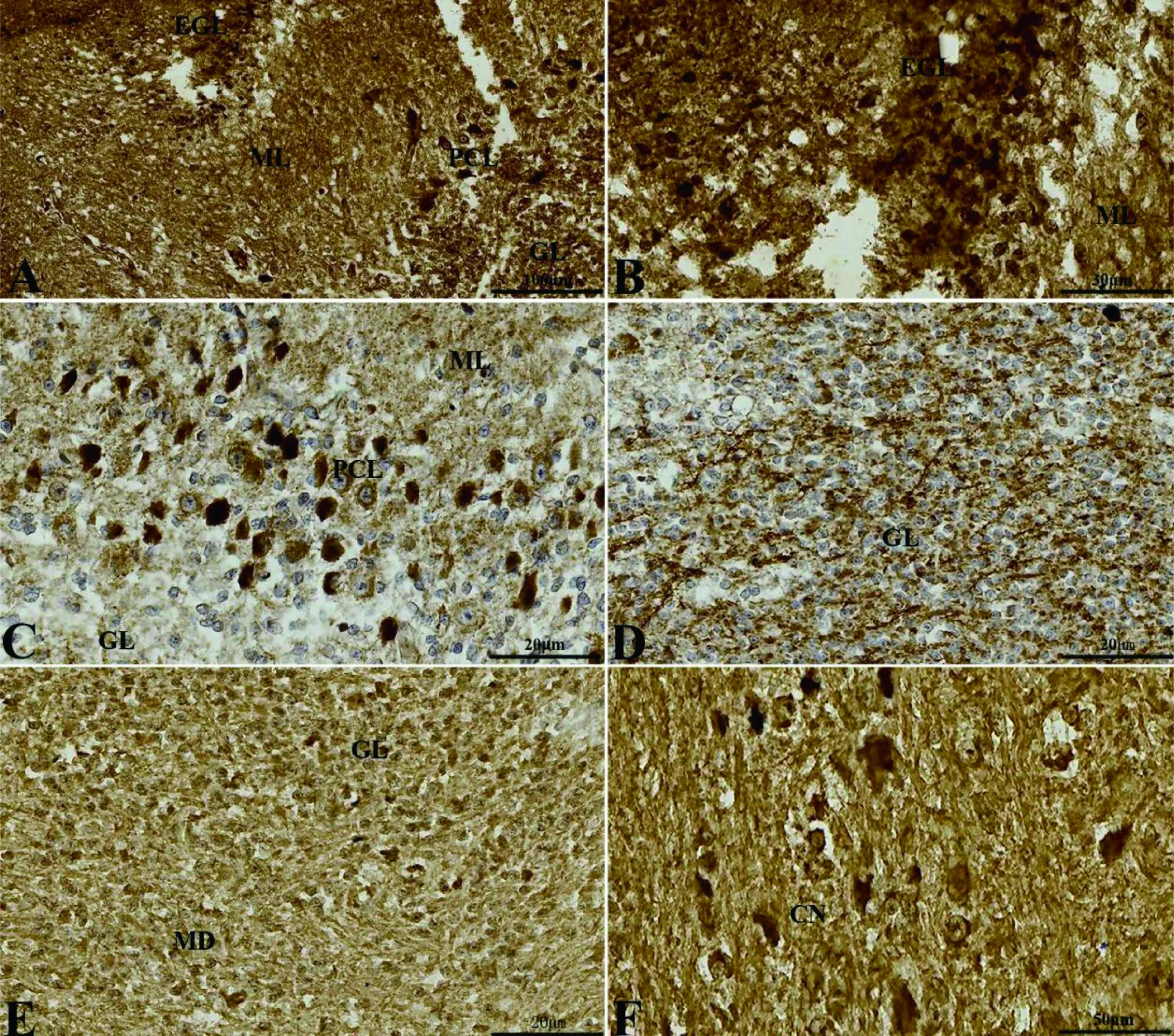
A: 小腦皮質分層(標尺: 100 μm); B: 外顆粒層 (EGL,標尺: 30 μm); C: 浦肯野細胞(PCL,標尺: 20 μm); D: 顆粒層(GL,標尺: 20 μm); E: 顆粒層與髓質(MD)的邊界(標尺: 20 μm); F: 髓質核團(CN,標尺: 50 μm)。ML: 分子層
圖4冠狀切面上5-HT在初孵揚子鱷小腦的分布
Figure 4 The distribution of the 5-HT in the cerebellum of the new-hatching Yangtze alligator in transversesections

A: 初孵揚子鱷小腦縱切(標尺: 300 μm); B: 外顆粒層(EGL,標尺: 20 μm); C: 浦肯野細胞(PCL,標尺: 20 μm); E: 小腦室(VC,標尺: 30 μm); E: 脈絡叢(CP,標尺: 80 μm)。ML: 分子層; GL: 顆粒層
圖5矢狀切面上5-HT在初孵揚子鱷小腦(CBe)的分布
Figure 5 The distribution of the 5-HT in the cerebellum (CBe) of the new-hatching Yangtze alligator in sagittal sections
冠狀切面上,外顆粒層細胞的細胞質為5-HT陽性(圖 4-B);浦肯野細胞顯示細胞質5-HT陽性,細胞核5-HT陰性(圖 4-C);分子層和顆粒層均顯示5-HT神經纖維陽性且陽性較強(圖 4-B、C、D)。髓質內側核團細胞體5-HT陽性(圖 4-F)。矢狀切面上,可見外顆粒層細胞5-HT陽性明顯(圖 5-B);分子層和顆粒層神經纖維5-HT陽性(圖 5-C);小腦室的室管膜細胞呈5-HT陰性(圖 5-D);脈絡叢細胞呈5-HT淺陽性(圖 5-E)。
2.3 DA在初孵揚子鱷小腦的分布
冠狀切面上,小腦皮質可明顯分為4層,從外向內為外顆粒層(external granular layer, EGL)、分子層(molecular layer, ML)、浦肯野細胞層(Purkinje cell layer, PCL)和顆粒層(granular layer, GL),見圖 6-A、B、C。其中浦肯野細胞可見明顯多細胞層(圖 6-C)。小腦髓質和皮質較易區分(圖 6-E),髓質可見小腦核(圖 6-F)。矢狀切面上,可見第四腦室由頂部的小腦和下部的腦干圍成,第四腦室后部由脈絡叢(choroid plexus, CP)不緊密封閉(圖 7-A),構成第四腦室頂部的小腦皮質部位可見明顯的外顆粒層(圖 7-B),小腦髓質內部可見小腦室(ventriculus cerebelli, VC),見圖 7-D。

A: 小腦皮質分層(標尺: 100 μm); B: 外顆粒層(EGL,標尺: 30 μm); C: 浦肯野細胞層(PCL,標尺: 20 μm); D: 小腦皮質與髓質(MD)的邊界(標尺: 50 μm); E: 髓質核團(CN,標尺: 30 μm); J: 第四腦室(V4)腦脊液物質(標尺: 20 μm)。ML: 分子層; GL: 顆粒層
圖6冠狀切面上DA在初孵揚子鱷小腦的分布
Figure 6 The distribution of the DA in the cerebellum of the new-hatching Yangtze alligator in transversesections

A: 初孵揚子鱷小腦縱切(標尺: 300 μm); B: 外顆粒層(EGL,標尺: 20 μm); C: 浦肯野細胞(PCL,標尺: 20 μm); D: 小腦室(VC,標尺: 30 μm); E: 脈絡叢(CP,標尺: 80 μm)。ML: 分子層; GL: 顆粒層
圖7矢狀切面上DA在初孵揚子鱷小腦(CBe)的分布
Figure 7 The distribution of the DA in the cerebellum (CBe) of the new-hatching Yangtze alligator in sagittal sections
冠狀切面上,外顆粒層細胞有微弱的陽性(圖 6-B);浦肯野細胞顯示細胞質DA陽性,細胞核DA陰性(圖 6-C);分子層和顆粒層均顯示DA神經纖維陽性(圖 6-B、C、D)。髓質內有較多DA免疫陽性纖維分布密集(圖 6-D),髓質內側核團細胞質DA陽性,細胞核陰性(圖 6-E)。第四腦腦室腦脊液物質也顯示DA免疫陽性(圖 6-F)。矢狀切面上,可見外顆粒層纖維DA陽性明顯(圖 7-B);分子層和顆粒層神經纖維DA陽性(圖 7-C);小腦室的室管膜細胞呈DA陰性(圖 7-D);脈絡叢細胞呈DA強陽性(圖 7-E)。
3 討論
由于爬行動物運動的復雜性,其小腦較兩棲動物更為發達,水生爬行動物的小腦就更加發達了;鱷的小腦已有分化成中央蚓部和兩側小腦鬈的趨向[14]。小腦在保持身體平衡、調節肌張力和協調肌群運動等方面都有著重要意義。劉再群等[13]曾對初孵揚子鱷小腦的生長抑素的分布進行了研究,而本文首次系統闡述了初孵揚子鱷小腦的組織學結構特征以及5-羥色胺和多巴胺在初孵揚子鱷小腦的分布情況。
3.1 小腦皮質分層
在爬行綱動物中,Larsell[15]曾研究了鱷目部分動物的小腦,他認為鱷類的小腦皮質可劃分成典型的3層,即分子層、浦肯野細胞層和顆粒層。報道表明其他一些爬行動物如紅耳龜[16]、中國石龍子[17]、尼羅鱷[18]的小腦皮質同樣可分為3層。成年揚子鱷的小腦皮質同樣可分3層,并不存在外顆粒層[12]。但是,本研究結果顯示初孵揚子鱷小腦皮質可分為4層,從外向內依次是外顆粒層、分子層、浦肯野細胞層和顆粒層。對小鼠小腦皮質的發生的研究表明,在小腦的組織生成過程中,來自原始神經上皮的一些神經前體細胞(也稱成神經細胞)會遷移到小腦的表面形成外顆粒層,在這層內側的那些已經停止增殖的成神經細胞則發展成小腦皮質顆粒細胞的前身,它們隨后遷移回到正在發生的白質中發展成內顆粒層[19]。我們推測,初孵時期的揚子鱷的小腦皮質確可分為4層,但伴隨著小腦發育成熟,小腦皮質的外顆粒層會逐漸消退。本實驗觀測到的正是初孵揚子鱷小腦外顆粒層殘余,它們會隨著小腦的發育成熟而逐漸退化直至消失。
3.2 浦肯野細胞層
浦肯野細胞的胞體呈梨形,所以也可稱之為梨狀神經細胞。浦肯野細胞層便是由排列整齊的浦肯野細胞的胞體構成。對老鼠小腦內浦肯野細胞的研究表明,浦肯野細胞可通過控制特定運動的區域的時相來協調多關節運動,且與四肢、軀干的協調有關[20]。長久以來,人們一直認為在爬行動物的小腦皮質中浦肯野細胞層是由單層胞體組成的[15, 17-18]。然而,我們在此注意到初孵揚子鱷小腦皮質的浦肯野細胞層其實是由多層浦肯野細胞胞體排列組成的。宋海燕等[21]曾在未發育成熟的皖西白鵝的小腦皮質觀察到部分浦肯野的多層排列且伴隨著小腦皮質的發育成熟多層浦肯野細胞最終逐漸退化成單層。有學者認為只有那些能夠與其他神經細胞建立正常突觸聯系的浦肯野細胞才能夠存活下來,而不能成功建立突觸聯系的細胞則會凋亡[22]。所以我們推測,在初孵揚子鱷小腦內,可能存在某種化學物質,它調控著浦肯野細胞的發育及突觸聯系的建立并最終使其成為均一的單細胞層,而那些沒能夠與其他神經細胞構建突觸聯系的浦肯野細胞便會出現凋亡,不能存活下來。
3.3 小腦室
陳壁輝等[12]認為成年揚子鱷小腦無空腔腦室,但是,本研究中,我們發現初孵揚子鱷小腦可見腦室,且小腦室與第四腦室相通。我們推測,揚子鱷的小腦室可能隨著小腦的發育增大而逐漸與第四腦室融合,不易區分。
3.4 5-羥色胺在小腦的分布
目前,關于5-HT免疫陽性神經細胞及纖維在爬行動物小腦的分布,僅有少量報道。有研究表明,幼年暹羅鱷(1個月)小腦內5-HT含量及濃度比成年鱷5-HT含量及濃度要高;此外,暹羅鱷小腦內的5-HT含量及濃度雖不高,但總體上要高于北美游蛇,可是其中樞神經系統其他區域5-HT含量及濃度低于其他爬行動物[2]。本研究中,我們觀察到初孵揚子鱷小腦的5-HT免疫陽性較強。鑒于小腦是運動也是平衡中樞,而5-HT涉及行為等各種活動的調控,這提示初孵揚子鱷小腦內5-HT可能參與揚子鱷運動的調節。在平原巨蜥[5]的小腦內,其小腦核側邊可觀察到5-HT免疫陽性神經細胞,且橫切面上小腦皮質可見分子層的免疫陽性纖維,小腦核周邊同樣可見免疫陽性纖維。我們注意到,在初孵揚子鱷小腦髓質的小腦核附近可見5-HT免疫陽性神經細胞及纖維。Challet等[6]發現蝰蛇小腦皮質的顆粒層內存在大量的5-HT能纖維。本研究中同樣觀察到初孵揚子鱷小腦的顆粒層內可見5-HT免疫陽性纖維。這些均提示在爬行綱的不同動物之間,5-HT在小腦的分布有一定的類似之處。有學者發現盡管錦龜小腦皮質浦肯野細胞層表現出對外源5-HT較強的反應且小腦皮質中可見5-HT能免疫陽性纖維,但是,其小腦內5-HT總體含量并不高[23]。我們發現,初孵揚子鱷小腦的5-HT免疫陽性較強,且不止見于浦肯野層。對彩龜腦干和脊髓5-HT分布的研究表明,小腦核會受到5-HT能神經纖維支配,且小腦皮質顆粒層可見些許背腹側行5-HT免疫陽性神經纖維,但是小腦皮質分子層未見5-HT免疫陽性纖維[24]。在初孵揚子鱷的小腦內,觀察到小腦皮質分子層和顆粒層均可見5-HT免疫陽性纖維,這表明在不同爬行動物的小腦,5-HT的分布也是存在一定差異的。
3.5 多巴胺在小腦的分布
目前未見DA在爬行動物小腦分布的相關報道。但是,在其他脊椎動物中已有一些小腦的DA免疫陽性神經細胞及纖維的分布的報道。歐洲鰻魚的小腦內僅包含稀疏的DA免疫陽性神經分布,小腦幾乎不接受DA能神經纖維的支配,DA能纖維稀疏的分布于小腦體側邊隆起的顆粒層內的顆粒細胞,在小腦聯合處也可見一些DA能纖維,這些纖維會形成一纖維束到達第四腦室上方的浦肯野細胞[25]。但是在本研究中,我們觀察到浦肯野細胞及髓質內側核團細胞均呈DA免疫陽性,分子層和顆粒層可見大量DA神經纖維陽性且髓質內也有較多DA免疫陽性纖維分布細胞質DA陽性,第四腦腦室腦脊液物質也顯示DA免疫陽性。考慮到小腦有維持平衡、協調運動的作用,揚子鱷是一種冬眠動物,推測DA在揚子鱷打洞等活動中發揮著重要的作用。對湖蛙和蠑螈小腦的DA的免疫組化分布的研究表明湖蛙和蠑螈小腦的腹側顆粒層區域可見少量的DA免疫陽性纖維[26]。本研究中,我們不僅在初孵揚子鱷的小腦皮質的顆粒層觀察到DA免疫陽性纖維,還在小腦其他各部分均觀察到DA免疫陽性。DA在運動控制、執行能力等方面都發揮了重要的作用,推測揚子鱷小腦DA的廣泛分布可能與其運動的復雜性有著密切的關系。在鳥綱動物中,灰斑鳩的小腦腳腹側可見DA免疫陽性纖維[27]。本研究中我們同樣觀察到在初孵揚子鱷的小腦內有豐富的DA免疫陽性分布。這提示DA在不同動物小腦的分布有一定的連續性。在哺乳綱動物中,對大鼠小腦內DA能神經分布的研究結果顯示整個大鼠小腦均可見DA能神經纖維分布,其中在小腦皮質的分子層分布最多,但是,未見免疫陽性神經細胞;免疫陽性神經纖維較細[28]。在此,我們在初孵揚子鱷的整個小腦均觀察到DA免疫陽性纖維且在顆粒層和髓質區域分布較為密集,同時,還觀察到浦肯野細胞也會表達DA免疫陽性。此外,還觀察到第四腦室腦脊液物質和脈絡叢細胞也呈DA免疫陽性,推測DA在揚子鱷幼體能產生一定的生理功能。
