油菜農桿菌介導轉化體系的優化
2019-08-17 06:48:28張姍姍耿思宇徐培林王景雪
山西農業科學 2019年8期
張姍姍,耿思宇,徐培林,王景雪
(1.山西大學生命科學學院,山西太原030006;2.中山大學教育部基因工程重點實驗室,廣東廣州510275)
油菜是我國重要的經濟作物,油菜籽油是我國主要的食用油之一,油菜籽榨油后的餅粕是較好的動物飼料,但是在生產中由于菜籽油中含有一定的芥子苷等有害物質,限制了油菜餅粕的利用。因此,油菜的遺傳改良尤為重要。在油菜的遺傳改良中,建立農桿菌介導的油菜高效轉化體系是進行油菜重要功能基因解析、遺傳改良的必要條件。
農桿菌介導轉化法是油菜轉基因研究中最常用的轉化方法。以往的研究發現,帶柄子葉[1]和下胚軸[2-3]是最常用的外植體;共培養時間對轉化效率有顯著影響,且共培養時間與基因型有關[4-7]。外植體與農桿菌共培養的時間過短或過長,都不利于獲得較高的轉化率。尤其是當外植體與農桿菌共培養時間過長時,外植體會出現嚴重褐化甚至死亡,顯著地降低了轉化率。與其他植物相比,油菜屬于難于進行遺傳轉化的物種,其在油菜遺傳轉化中存在著轉化率低,轉化過程中外植體易褐化死亡等問題。為解決這些問題,研究者對油菜遺傳轉化條件進行了優化。
有研究發現,農桿菌侵染外植體前,對外植體進行預培養,向分化培養基中加入AgNO3等都可以提高油菜的轉化率[8-9]。即使是難再生的Brassica rapa,通過向培養基中加入乙烯抑制劑如氨乙氧基乙烯基甘氨酸(aminoethoxyvinylglycine)和硝酸銀等,也獲得了再生苗[10-13]。一些研究表明,硝酸銀對于苗充分再生是專性的[14-15],尤其當選擇標記為卡那霉素時[16]。但是,AgNO3的濃度也不能過高,過高的Ag2+會對植物產生毒害。以往研究表明,在共培養前,外植體在愈傷誘導培養基上進行一定時間的預培養能夠顯著增加轉化率[17-20]。GONG 等[21]研究發現,AgNO3會誘導芥菜細胞中再生相關基因的表達。有研究表明,48 h 是農桿菌介導轉化體系的最佳共培養時間[22]。李韓帥等[23]研究發現,農桿菌菌液OD600為0.9~1.2 是玉米農桿菌介導轉化體系的最適濃度。
為了更加系統地對農桿菌介導的油菜基因轉化體系進行優化,本試驗全方位、系統地研究了農桿菌介導轉化中預培養時間、農桿菌侵染濃度、培養基中添加AgNO3的濃度和共培養基pH 值等對農桿菌轉化中抗性芽苗分化的影響,以期獲得優化的農桿菌介導的油菜轉化體系。
1 材料和方法
1.1 試驗材料和試劑
供試甘藍型油菜(Brassica napus L.)為湘油15 號,其種子由湖南農業大學油料研究所提供。
供試質粒為pGRA1300,質粒圖譜如圖1 所示。質粒pGRA1300 的宿主菌為農桿菌(Agrobacterium tumefaciens)菌株LBA4404;培養基中的無機鹽和維生素均為分析純級;固化劑瓊脂粉購于鼎國生物工程有限公司;用于植物培養的氨基酸和植物激素均購于Sigma 公司。
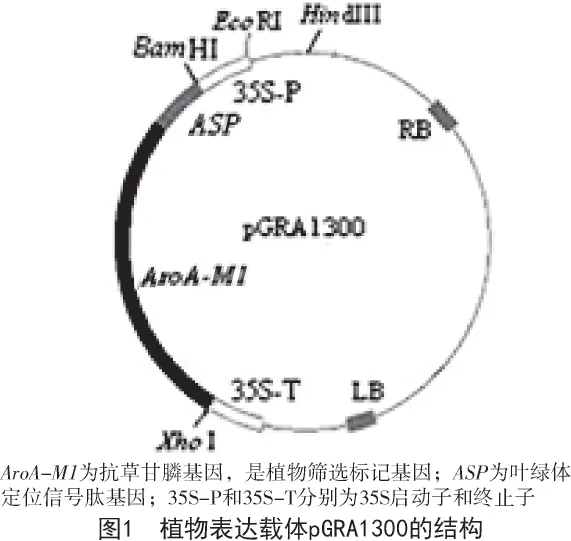
1.2 試驗方法
以湘油15 號為試驗材料,對影響農桿菌轉化的各種因素進行單因素分析。除了所研究因素不同外,其他操作均按下述農桿菌轉化程序進行。
1.2.1 轉化方法 將5~6 d 苗齡的無菌苗下胚軸切成約0.5~1.0 cm 的切段或帶0.3 cm 左右子葉柄的子葉切下,置于MS+2 mg/L 6-BA 培養基上分別進行2 d 的預培養,然后用OD600為0.4 的菌液侵染5 min。之后用無菌濾紙吸干外植體上多余的菌液。將外植體放置在MS+2 mg/L 6-BA 培養基上共培養2 d。然后將外植體放入含有300 mg/L Cef 的無菌水中沖洗2~3 次。洗掉附著在表面的菌體,放入MS+3 mg/L 6-BA+0.2 mg/L GA3+20 μmol/L AgNO3+8 mg/L 草甘膦+300 mg/L Cef 培養基中誘導愈傷組織分化和芽苗再生。20 d 后用原培養基繼代一次。外植體分化出小芽后,轉入新的篩選培養基中使小芽繼續生長。待分化出的小芽苗長到3 cm 左右時,將小芽苗從外植體上切下,放入生根培養基中誘導生根。
1.2.2 農桿菌轉化條件的優化 (1)預培養時間。將下胚軸或子葉外植體,在MS+2 mg/L 6-BA 培養基上分別進行0,1,2,3,4 d 的預培養。(2)農桿菌轉化菌液濃度。將預培養過的下胚軸和子葉外植體,分別用OD600為0.2,0.4,0.6,0.8,1.0 的菌液侵染5 min。(3)誘導培養基中添加AgNO3的濃度。將轉化過的外植體,分別放在加有AgNO3濃度為0,10,20,30,40 μmol/L 的MS+3 mg/L 6-BA+0.2 mg/L GA3+20 μmol/L AgNO3+8 mg/L 草甘膦+300 mg/L Cef 培養基上誘導愈傷組織和芽苗分化。(4)共培養基pH 值。將感染過的下胚軸或子葉外植體,分別置于pH 值為5.0,5.2,5.4,5.8 培養基上共培養2 d。之后,按照上述轉化程序進行轉化。轉化后40 d 時,統計不同處理條件下外植體的出愈率和抗性芽苗分化率。
所有的試驗處理均為3 次重復,所有的外植體培養均為(25±1)℃,光照周期為16 h 光照/8 h 黑暗。轉化后篩選劑為草甘膦,抗性芽苗分化階段篩選壓為8 mg/L 草甘膦。
2 結果與分析
2.1 外植體預培養對抗性芽苗分化的影響
子葉和下胚軸外植體在不同預培養時間下的出愈率和抗性芽苗分化率列于表1。不同預培養時間下外植體愈傷組織生長和抗性芽苗分化情況如圖2、圖3 所示。
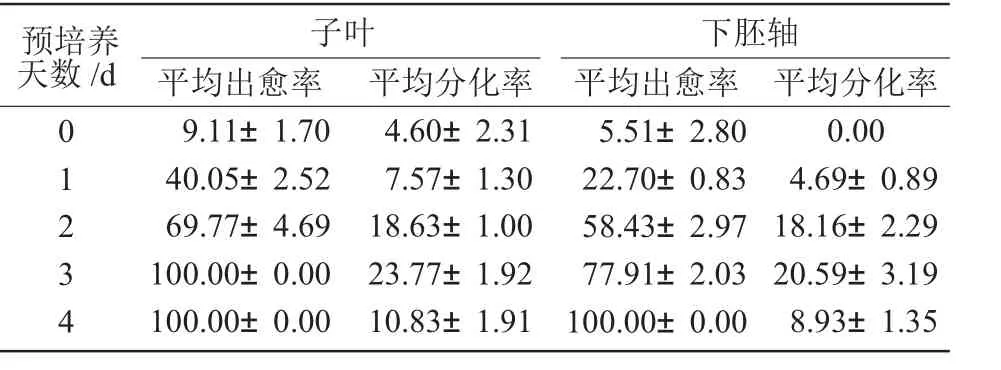
表1 預培養時間對抗性芽苗分化的影響 %
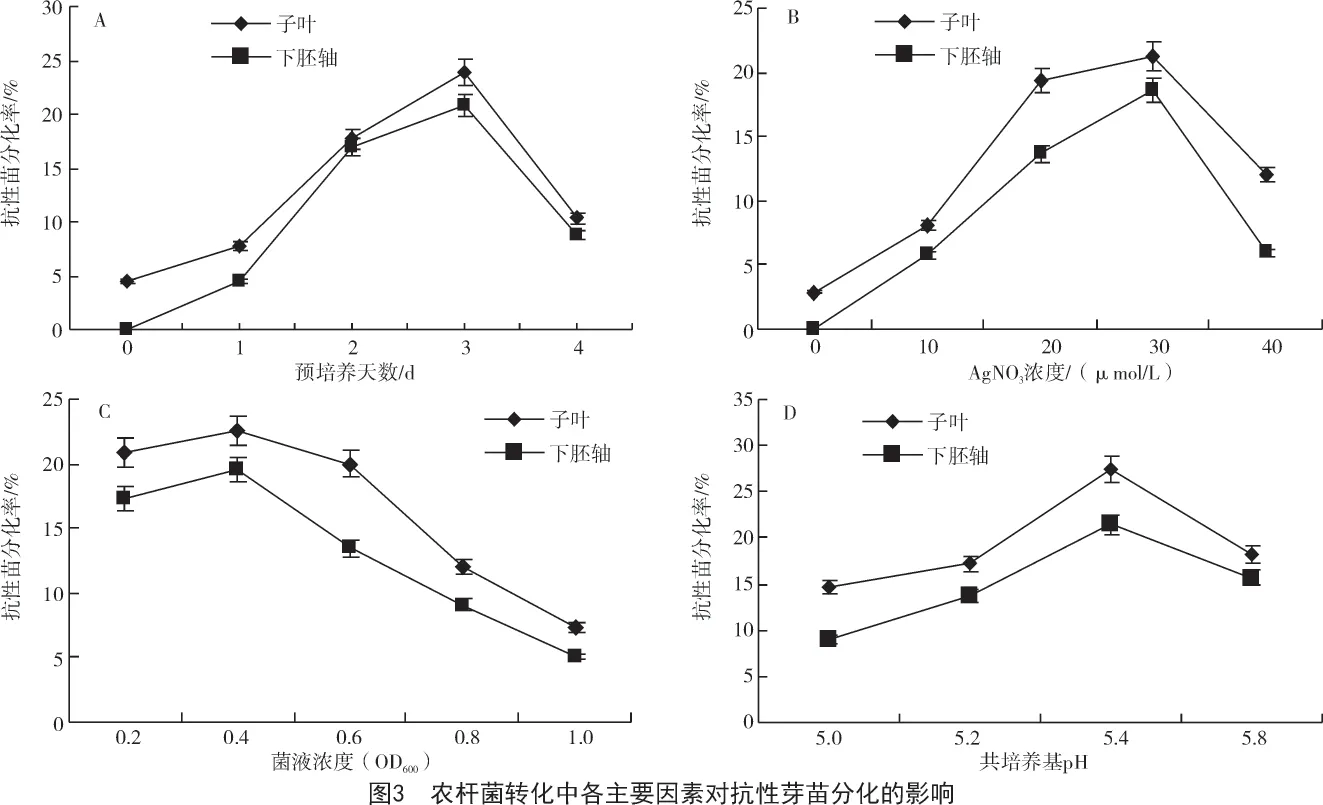
從表1 和圖2、圖3-A 可以看出,預培養對于高效率轉化非常重要,預培養3 d 無論是對子葉外植體還是對下胚軸外植體,其出愈率和抗性芽苗分化率都是最高的,子葉和下胚軸的抗性芽苗最高分化率分別為23.77%和20.59%。因此,外植體在農桿菌侵染和共培養前進行3 d 預培養有利于獲得較高的轉化效率。
2.2 不同AgNO3 濃度對抗性芽苗分化的影響
外植體經與農桿菌共培養后,置于附加有不同濃度AgNO3的分化培養基上培養42~56 d 后,在外植體上誘導出抗性芽苗;當添加0~30 μmol/L AgNO3時,抗性芽苗分化率隨著AgNO3濃度的增加而提高;但是當AgNO3濃度超過30 μmol/L 時,芽苗分化率會有所降低(圖3-B)。說明向培養基中添加AgNO3雖然可以提高抗性芽苗分化率,但是過高的AgNO3會對植物抗性芽苗的分化產生不利影響。在本試驗條件下,在基因轉化中AgNO3的最佳使用濃度為30 μmol/L。此時,添加AgNO3的處理分別比不添加的處理,子葉和下胚軸的抗性芽苗分化率分別提高19.34,17.96 百分點(圖3-B)。
2.3 農桿菌侵染的菌液OD600 對抗性芽苗分化的影響
通過對油菜農桿菌轉化后外植體生長發育的觀察發現,造成油菜基因轉化中轉化率低的主要原因是在油菜外植體經過農桿菌侵染和與農桿菌共培養后外植體上會出現嚴重的褐化現象。而且外植體褐化的程度與農桿菌侵染時農桿菌菌液濃度有關。在農桿菌侵染時間為5 min 時,用不同濃度的農桿菌菌液轉化后,外植體的出愈率和抗性芽苗的分化率列于表2。由表2 可知,農桿菌菌液濃度對轉化率有顯著影響,當農桿菌菌液OD600為1.0 時,子葉外植體的出愈率和抗性芽苗的分化率分別比菌液OD600為0.4 時下降34.69,15.38 百分點;下胚軸外植體的出愈率和抗性芽苗的分化率分別比菌液OD600為0.4 時下降26.15,13.39 百分點。從圖3-C可以看出,過高和過低的菌液濃度都不利于獲得高轉化效率,在本試驗中,無論子葉還是下胚軸,芽苗的最大分化率均是在農桿菌菌液OD600為0.4 時得到的,此時子葉和下胚軸的分化率分別為23.00%和18.95%。因此,在農桿菌介導的油菜基因轉化中,當侵染時間為5 min 時,最佳的農桿菌菌液OD600為0.4。
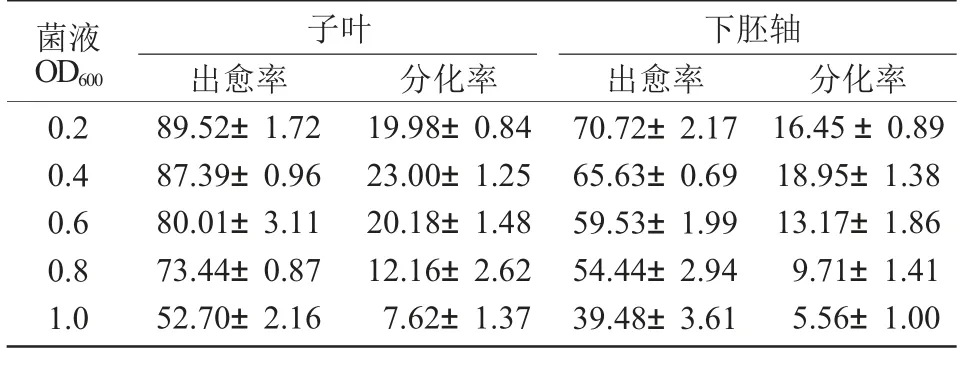
表2 農桿菌侵染濃度對抗性芽苗分化的影響 %
2.4 共培養基pH 值對抗性芽苗分化的影響
從圖3-D 可以看出,共培養基pH 值5.4 時,抗性芽苗的分化率最高;共培養基pH 值為5.0 和5.2時,外植體上褐化比較嚴重,可能是由于較低的共培養基pH 值有利于農桿菌的生長。觀察發現,共培養基pH 值為5.4 時,外植體上愈傷組織的生長狀態優于其他3 種培養基上的愈傷組織,愈傷組織致密性較好;子葉和下胚軸外植體在共培養基pH 值為5.4 時,抗性芽苗分化率分別比共培養基pH 值為5.8 時提高9.97,6.18 百分點。
3 結論與討論
本研究結果發現,外植體在農桿菌侵染前進行一定時間的預培養,農桿菌侵染及共培養后向誘導培養基中加入一定量AgNO3,可以提高油菜轉化率,這與之前的研究結果相一致[24-26]。本研究還發現,優化農桿菌侵染的菌液濃度和共培養培養基的pH 值也可以提高油菜的基因轉化率。有研究表明,植物進行離體培養時會產生乙烯,乙烯的積累會造成外植體的褐化。過高濃度的農桿菌侵染菌液也是造成外植體褐化的原因之一。AgNO3是乙烯的抑制劑,因此向農桿菌侵染后誘導培養基中添加30 μmol/L AgNO3,以及在農桿菌侵染時采用較低濃度的菌液侵染,可以減少油菜轉基因中外植體離體培養時乙烯產生量,減少外植體褐化,促進抗性芽苗的分化[27],農桿菌菌液濃度以OD600為0.4 時較好,這與何業華等[28]的研究結果相似。共培養基pH 值在油菜轉化中的作用,前人很少研究。楊廣東等[29]在大白菜上的研究認為,較低的共培養基pH 值(5.0)比較高的共培養基pH 值(5.8)誘芽率提高近2 倍。STACHEL 等[30]研究認為,pH 值在5.8 以上會使根癌農桿菌vir 基因處于不活化的狀態。因此,較低的共培養基pH 值有利于vir 基因的誘導。TAKASAKI等[31]研究認為,pH 值為5.2 的共培養基比pH 值為5.8 的共培養基更有利于獲得較高的轉化率。本研究結果認為,共培養基pH 值為5.4 有利于油菜農桿菌介導轉化后抗性芽苗的分化。不同植物的最適預培養時間不同,任偉等[32]研究表明,紫花苜蓿在農桿菌侵染前,預培養3 d 效果最好。
本試驗結果表明,優化的油菜農桿菌介導轉化體系為:外植體在農桿菌侵染前進行2~3 d 預培養、培養基中添加30 μmol/L AgNO3、農桿菌侵染的菌液OD600為0.4、侵染5 min、共培養基的pH 值為5.4。
