常壓室溫等離子體誘變選育木聚糖酶高產菌及其酶學性質研究
2019-07-30 03:16:04朱慧霞魯旭峰汪水玲馬曉靜姚日生
中國釀造 2019年7期
關鍵詞:生長
朱慧霞,方 楨,魯旭峰,汪水玲,馬曉靜,姚日生,2*
(1.合肥工業大學 食品與生物工程學院,安徽 合肥 230009;2.農產品生物化工教育部工程研究中心,安徽 合肥 230009)
木聚糖酶是一種重要的半纖維素水解酶系,能將木聚糖降解為木寡糖和木糖[1],已被用于生物燃料乙醇、食品、醫藥以及動物飼料等領域[2-3]。PARAB P等[4]利用芽孢桿菌(Bacillussp.)產生的木聚糖酶酶解海藻,得到可發酵的還原糖;DHIMAN S S等[5]利用嗜熱脂肪芽孢桿菌(Bacillus stearothermophilus)所產木聚糖酶處理柑橘果汁,通過降解其中果膠、淀粉和半纖維素等大分子多糖,獲得了高度澄清的柑橘果汁;KHAMBHATY Y等[6]將木聚糖酶用于紙漿預處理,能有效提高紙漿的白度。
木聚糖酶來源廣泛,有微生物、甲殼動物、植物、昆蟲、種子等。其中微生物主要包括真菌(如黑曲霉、煙曲霉、米曲霉、木霉、青霉等)、細菌(如地衣芽孢桿菌、黃熱芽孢桿菌、嗜熱雙胞菌、枯草芽孢桿菌、雙歧桿菌等)和放線菌[7-9]。一般認為微生物是生產商品木聚糖酶的最佳來源之一[10]。鄭麗麗等[11]從土壤中篩得一株黑曲霉,其所產木聚糖酶酶活為78.34 U/mL;KUMAR V等[12]從廢棄木材中篩得野生菌疏棉狀嗜熱霉菌,其所產木聚糖酶酶活為61.09 U/mL;吳仁智等[13]從土壤中篩得野生日本曲霉菌,其所產木聚糖酶酶活為26.26U/mL。由于野生菌所產木聚糖酶酶活較低,而對工業化生產而言,木聚糖酶酶活高不僅能提高生產效率、亦能有效節約能源[14],因此可采用一些方法提高野生菌的產酶能力,誘變育種是其中一種有效方法。
常壓室溫等離子體(atmospheric and room temperature plasma,ARTP)技術是近些年發展起來的一種新型微生物誘變育種方法,ARTP富含的活性粒子可以改變細胞壁和細胞膜的物理化學性質,并造成組織損傷和遺傳物質損傷,微生物細胞被迫啟動容錯水平高的“SOS修復機制”、出現較多錯誤傾向修復,引起微生物的基因突變[15-16]。ARTP誘變技術具有突變率高、成本低、環境友好等優點[17],已成為誘變領域的研究熱點。
本研究以黑曲霉(Aspergillus niger)FXY為出發菌株,通過ARTP技術進行誘變處理,旨在找到一株木聚糖酶高產突變株,并對突變菌株的酶活提高原因、所產木聚糖酶的酶學性質進行研究,為木聚糖酶的工業化生產和應用范圍擴大提供依據,對促進我國酶制劑工業的發展具有現實意義。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株與試劑
黑曲霉(Aspergillus niger)FXY:本實驗室保藏。
木聚糖(純度為99%):美國Sigma-Aldrich公司;3,5-二硝基水楊酸(3,5-dinitrosalicylic acid,DNS)(分析純):廈門海標科技有限公司;葡萄糖、MgSO4·7H2O、KH2PO4(均為分析純):國藥集團化學試劑有限公司;蛋白胨、酵母粉(均為生化試劑):北京奧博星生物技術有限責任公司。
1.1.2 培養基
馬鈴薯葡萄糖瓊脂(potatodextroseagar,PDA)培養基:稱取200g馬鈴薯切成小塊,加水煮爛(煮沸20~30min,能被玻璃棒戳破即可),用8層紗布過濾,加入15~20g瓊脂,繼續加熱攪拌混勻,待瓊脂溶解完后,加入葡萄糖20g,攪拌均勻,稍冷卻后再補足水分至1000mL,121℃高壓滅菌20min。
種子培養基:葡萄糖20.0 g/L,蛋白胨20.0 g/L,酵母粉10.0 g/L,pH自然,121℃高壓滅菌20 min。
初篩培養基:木聚糖10.0 g/L,酵母粉3.0 g/L,KH2PO42.0 g/L,MgSO4·7H2O 2.0 g/L,瓊脂 20.0 g/L,pH自然,121 ℃高壓滅菌20 min。
發酵產酶培養基:麩皮20.0 g/L,葡萄糖5.0 g/L,酵母粉5.0 g/L,MgSO4·7H2O 1.0 g/L,KH2PO41.0 g/L,pH自然,121℃高壓滅菌20 min。
1.2 儀器與設備
ARTP-Ⅱ型誘變育種儀:無錫源清天木生物科技有限公司;DHP-9052型電熱恒溫培養箱:上海一恒科技有限公司;LDZX-40SC型電熱壓力蒸汽滅菌器:上海申安醫療器械廠;SW-CJ-2FD型雙人單面超凈工作臺:蘇州凈化設備有限公司;HQL-300A型柜式恒溫冷凍搖床:中國科學院武漢科學儀器廠;WFI800-D3B型紫外可見分光光度計:北京瑞利分析儀器廠。
1.3 方法
1.3.1 木聚糖酶酶活的測定
木聚糖酶的酶活用DNS法[18]測定。木聚糖酶酶活定義:在一定條件下(50.0℃、pH 5.0),每分鐘酶解木聚糖產生1.0 μmol木糖所需的酶量為1個酶活單位(U)。發酵所產木聚糖酶的酶活是以每毫升發酵液所含酶活單位表示,即為U/mL。
1.3.2 孢子懸浮液的制備
將保藏的黑曲霉FXY接入種子培養基,于30.0℃、180r/min活化48.0 h后轉接至固體培養基,之后于30.0℃培養96.0 h。用生理鹽水將不同培養時間的黑曲霉孢子洗脫至盛有玻璃珠的三角瓶中,于220 r/min充分振蕩使其活化并分散。之后經無菌孢子過濾器過濾后用生理鹽水將孢子懸浮液稀釋至106~108個/mL。
1.3.3 ARTP誘變
將10.0 μL不同培養時間的黑曲霉FXY孢子懸浮液均勻涂布于無菌金屬載片上,并分別暴露于ARTP噴射器下1.0~12.0 min。每次處理后,將載片轉移至裝有1.0 mL無菌水的無菌管中,振蕩洗脫后進行稀釋涂布,用未經ARTP處理的菌株作為對照,于30.0℃培養72.0h后進行菌落計數,分別用公式(1)、(2)計算致死率、正突變率,并繪制致死率曲線、正突變率曲線。將酶活提高超過對照菌株20.0%及以上的菌株定義為正突變菌株。

1.3.4 初篩
根據誘變的致死率曲線,選取適當的誘變條件進行誘變處理,將處理后的孢子懸浮液稀釋涂布至初篩培養基上,于30.0℃、180 r/min培養72.0 h。產酶菌株會在初篩培養基上產生透明圈,測定透明圈與菌落直徑的大小,選取透明圈直徑(H)與菌落直徑(C)比值(即H/C值)較大的菌落進行后續的復篩實驗。
1.3.5 復篩
將初篩得到的菌落接種至種子培養基中培養48.0 h,再以2.0%的接種量轉接入發酵產酶培養基中,于30.0℃、180r/min培養96.0h后離心收集發酵液,取上清液適當稀釋后測其木聚糖酶酶活,與菌株FXY在相同條件下發酵所得粗酶液的酶活進行比較。
1.3.6 誘變菌株的遺傳穩定性實驗
將誘變所得優勢菌株進行連續傳代培養,并將各代菌株轉接至發酵產酶培養基,測定酶活,比較各代菌株產酶變化情況,以檢測誘變菌株的遺傳穩定性。
1.3.7 生長曲線的測定
將孢子懸浮液以接種量2.0%接入裝液量為50mL/250mL種子培養基中,于30.0℃、180r/min培養。每隔4.0h取出發酵液離心,棄去上清液,并將固體放入105.0℃烘箱中干燥至恒質量。通過測定干細胞質量繪制生長曲線。
1.3.8 產酶曲線的測定
將已培養48.0 h的種子液以2.0%的接種量接入發酵培養基中,于30.0℃、180 r/min培養,每隔12.0 h取樣,測定發酵液中木聚糖酶酶活,并用考馬斯亮藍法[19]測定發酵液中的蛋白含量。
1.3.9 木聚糖酶酶學性質
最適溫度的確定:將發酵液于8000r/min離心20.0min,上清液即為粗酶液。將粗酶液進行適當稀釋,分別于35.0℃、40.0℃、45.0℃、50.0℃、55.0℃、60.0℃進行酶解反應,測定酶活。將木聚糖酶酶活的最高值定義為100.0%,以計算不同溫度條件下的相對酶活。
熱穩定性實驗:將粗酶液進行適當稀釋,分別在上述溫度保溫30.0~180.0 min后測定酶活。將各溫度未保溫的木聚糖酶酶活最高值定義為100.0%,以計算在不同溫度條件下保溫不同時間的相對酶活。
最適反應pH值的確定:將粗酶液進行適當稀釋,分別在檸檬酸-檸檬酸鈉緩沖溶液(pH值為3.0~4.5)、磷酸鹽緩沖溶液(pH值為5.0~8.0)、甘氨酸-氫氧化鈉緩沖溶液(pH值為8.5~10.0)、磷酸氫二鈉-氫氧化鈉緩沖溶液(pH值為10.5~11.0)條件下反應,測定酶活。將木聚糖酶酶活的最高值定義為100.0%,以計算不同pH值條件下的相對酶活。
pH穩定性實驗:將粗酶液進行適當稀釋,分別在上述pH值孵育60min后測定酶活。將各pH值未孵育的木聚糖酶酶活的最高值定義為100.0%,以計算在不同pH值條件下孵育60 min后的相對酶活。
2 結果與分析
2.1 誘變條件的篩選
對不同培養時間的黑曲霉FXY進行ARTP誘變,其致死率曲線如圖1所示。由圖1可知,菌株FXY培養時間越短,ARTP處理相同時間所對應的致死率越高;培養時間越長,ARTP處理相同時間所對應的致死率越低。這是因為菌株FXY培養時間越長,孢子細胞壁越厚,等離子體中的活性粒子進入細胞受到的阻力會變大,致使活性粒子不易穿透細胞,所以致死率降低。培養4 d的菌株FXY所制備的孢子懸浮液經誘變處理后,致死曲線出現了“馬鞍形”,即致死率呈現先升高、后下降、再升高的趨勢,是因為ARTP誘變過程中產生了“質量沉積效應”和“電荷交換效應”,在這兩種效應的刺激下,可激活菌株體內的應急反應機制(SOS修復)[20-21],該機制是一種自我修復方式、能夠降低菌株的致死率,而隨著ARTP處理時間進一步延長,受損的菌株細胞不能完全進行自我修復,所以菌株的致死率又會升高[22]。
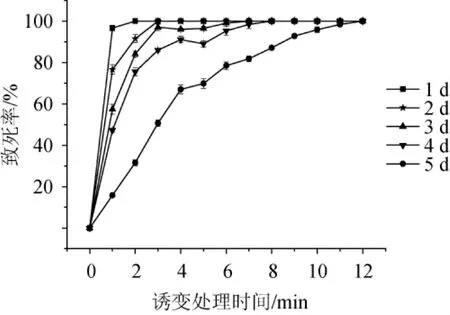
圖1 黑曲霉FXY進行常壓室溫等離子體誘變的致死率曲線Fig.1 Lethality curve ofAspergillus nigerFXY mutated by atmospheric and room temperature plasma
根據現代誘變理論,誘變致死率在95.0%時有較高的正突變率[23],因此對不同培養時間致死率最接近95.0%的菌株進行產酶篩選,其正突變率結果如圖2所示,培養4 d的菌株所對應的正突變率最高,達到了45.6%,因此確定ARTP誘變處理時菌株FXY的培養時間為4 d,誘變處理時間為5.0 min。
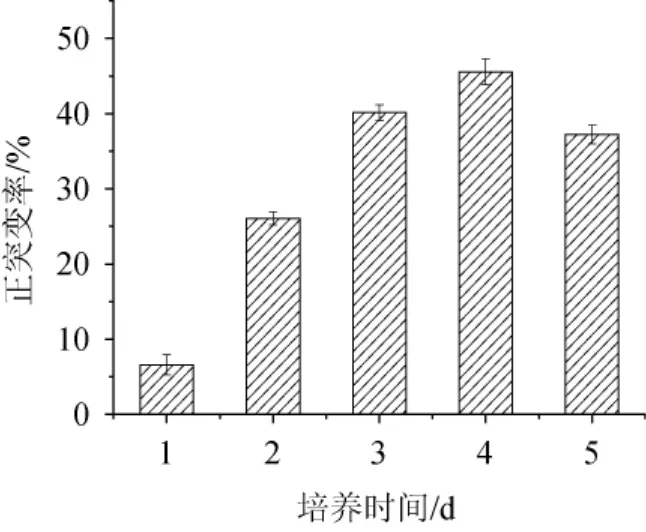
圖2 黑曲霉FXY進行常壓室溫等離子體誘變的正突變率結果Fig.2 Positive mutation rate results ofAspergillus nigerFXY mutated by atmospheric and room temperature plasma
2.2 誘變菌株的篩選結果
出發菌株黑曲霉FXY的H/C值為1.4,通過比較初篩所得各菌株H/C值大小,從中挑選了49株H/C值超過3.0的菌株,對這49株菌進行產酶發酵并測定酶活,結果如圖3所示,與出發菌株(79.42 U/mL)相比,共有42株菌的酶活提高了20.0%及以上,其中編號為ARTP-19、ARTP-49、ARTP-80、ARTP-82的4株菌的酶活提高了100.0%及以上,故后續對這4株優勢菌株進行遺傳穩定性實驗。
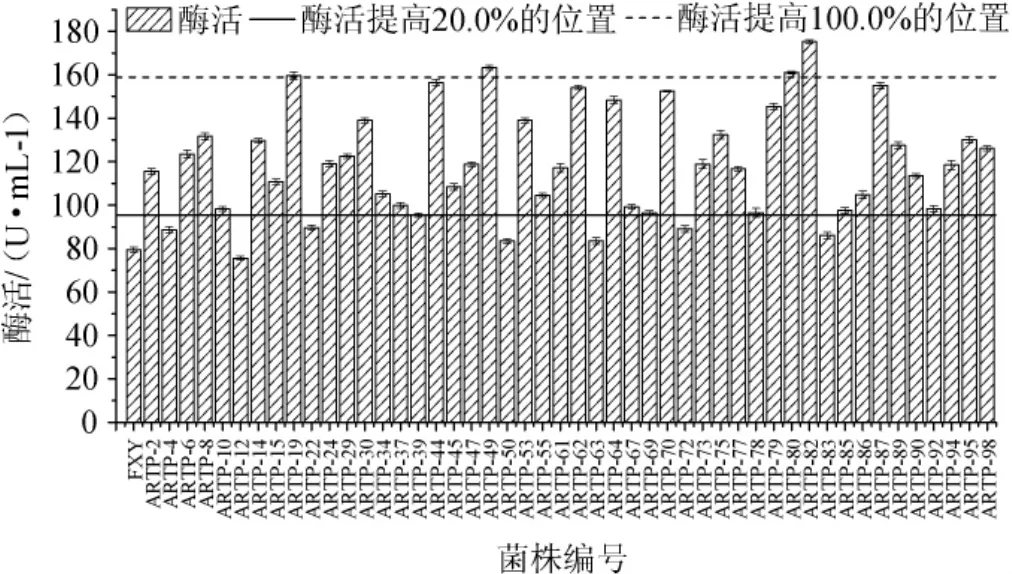
圖3 常壓室溫等離子體誘變菌株所產木聚糖酶酶活Fig.3 Activities of xylanase produced by atmospheric and roomies temperature plasma mutagenized strains
2.3 誘變優勢菌株的遺傳穩定性實驗
將編號為ARTP-19、ARTP-49、ARTP-80、ARTP-82這4株優勢菌株進行連續傳代培養,各菌株分別連續傳代5次,并將各代菌株轉接至發酵產酶培養基中,培養96 h后測定發酵液中木聚糖酶酶活,結果如表1所示。由表1可知,經過5次連續傳代培養,這4株正突變菌株均能穩定遺傳,其中編號為ARTP-82的菌株所產木聚糖酶酶活最高,達到了175.33 U/mL,較出發菌株(79.42 U/mL)提高了120.8%,因此選用菌株ARTP-82進行后續研究。
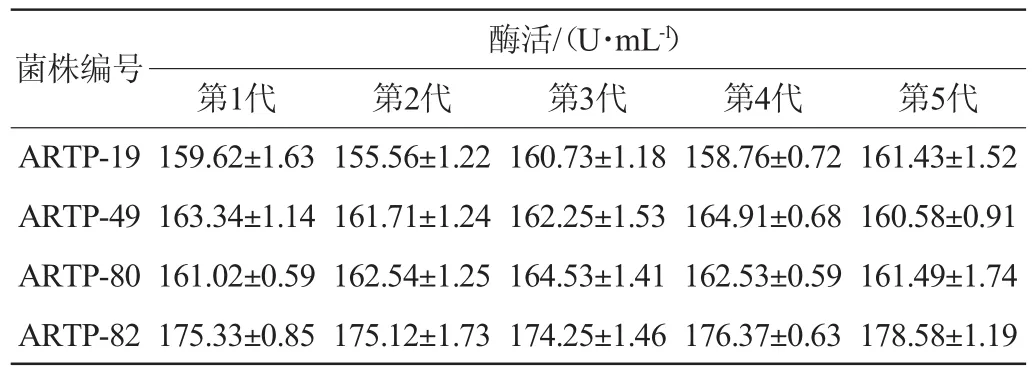
表1 常壓室溫等離子體誘變優勢菌株的遺傳穩定性實驗Table 1 Hereditary stability experiments of atmospheric and room temperature plasma mutagenic dominant strains
2.4 生長曲線的測定
采用干重法測定黑曲霉FXY和菌株ARTP-82的生長曲線,結果如圖4所示。由圖4可知,0~12 h這兩株菌生長均較慢,處于“延滯期”,菌體干質量增加少。12 h后,菌株都已適應生長環境,生長速率加快、菌體干重值迅速上升,此時菌株處于“對數生長期”。從36 h開始,這兩株菌的生長速率幾乎都不變,處于生長的“平穩期”,因此菌體干質量都基本保持不變。比較這兩株菌的生長曲線,發現這兩株菌在12h后菌體干質量就有差異,在24.0h時,菌株ARTP-82的生物量6.1 g/L比菌株FXY(7.5 g/L)低23.0%,差異達到最大;在36 h時,菌株生長達到平穩期,ARTP-82的生物量9.6 g/L比菌株FXY(10.2 g/L)低5.9%。可見ARTP誘變確實改變了黑曲霉的生長并對其產木聚糖酶也造成了影響,后續實驗也驗證了這一推斷。
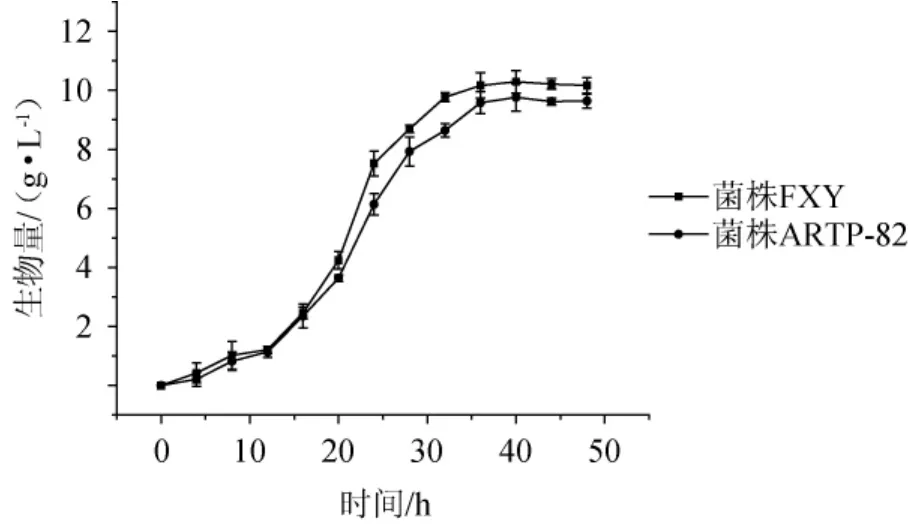
圖4 菌株FXY和菌株ARTP-82的生長曲線Fig.4 Growth curves of strain FXY and strain ARTP-82
2.5 發酵產酶曲線
菌株FXY和菌株ARTP-82的發酵產酶曲線如圖5所示,發酵前期,由于菌株主要利用營養物質進行生長,因此產酶較少,蛋白量也較少。當發酵時間達到36 h后,菌株開始大量產酶,蛋白含量亦隨之增加。當發酵至96 h時,木聚糖酶酶活達到最高,分別為79.33 U/mL和174.32 U/mL。隨著發酵時間再延長,酶活有所下降,可能是因為菌株的代謝產物對酶有抑制作用,而此時蛋白含量不變,這是因為菌株停止產酶,而代謝產物不影響蛋白含量。對比菌株FXY和菌株ARTP-82,在發酵108 h后,兩株菌的發酵液中蛋白含量基本保持不變,菌株ARTP-82蛋白含量維持在6.8 mg/mL,比菌株FXY(5.5 mg/mL)提高了23.6%,結合菌株的生長曲線可知,菌株ARTP-82木聚糖酶酶活的增加,與其生物量增加無關、與酶比活力增加有關。
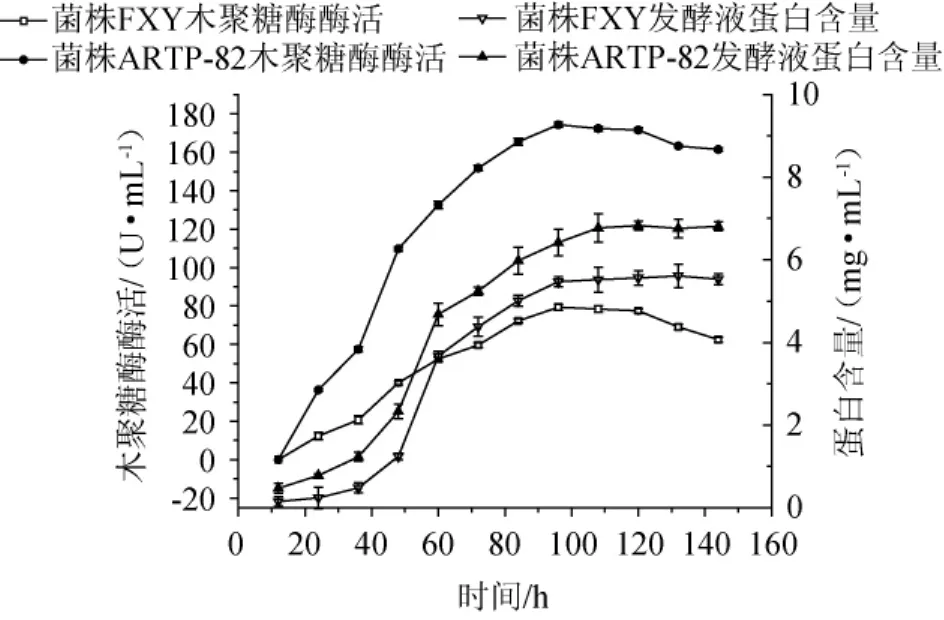
圖5 菌株FXY和菌株ARTP-82的發酵產酶曲線Fig.5 Enzyme production curves by fermentation of strain FXY andstrain ARTP-82
2.6 木聚糖酶的最適反應溫度及熱穩定性
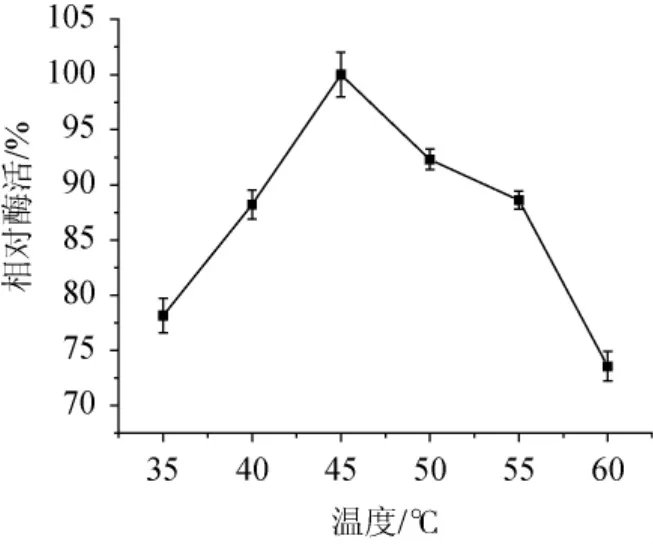
圖6 菌株ARTP-82產木聚糖酶的最適反應溫度Fig.6 Optimum reaction temperature of xylanase produced by strain ARTP-82
不同溫度條件下木聚糖酶相對酶活如圖6所示,木聚糖酶在40~55℃時能保持80%以上的相對酶活,最適反應溫度為45℃,相對酶活為100%。溫度較低時,由于酶解反應速率較慢、產生的還原糖少,用DNS法測定酶活,顯示相對酶活較低;而溫度過高會導致部分木聚糖酶失活,因此相對酶活也會降低。木聚糖酶的熱穩定性實驗結果如圖7所示,該木聚糖酶在35~45℃條件下保溫30~180 min,酶活穩定。當溫度>45℃,隨著保溫時間的延長,木聚糖酶的相對酶活會隨之降低。在50℃、保溫60.0 min后木聚糖酶相對酶活下降了8.5%,保溫180min后相對酶活下降了28.8%。在60℃保溫60min后木聚糖酶的相對酶活就下降了21.5%,而保溫180 min后,相對酶活只有15.7%。這說明溫度越高,保溫時間越長,木聚糖酶越容易失活,相對酶活也就越低,熱穩定性越差。
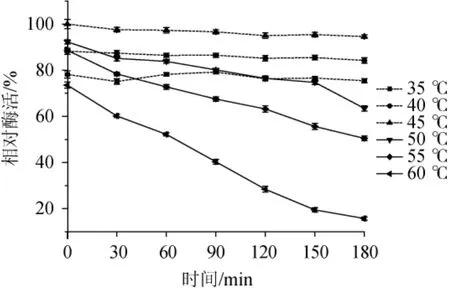
圖7 菌株ARTP-82產木聚糖酶的熱穩定性Fig.7 Thermal stability of xylanase produced by strain ARTP-82
2.7 木聚糖酶的最適反應pH值及pH穩定性
不同pH值條件下木聚糖酶相對酶活如圖8所示,木聚糖酶在pH值為3~8之間能保持85.0%以上的相對酶活,最適反應pH值為7,反應環境偏酸和偏堿都會使木聚糖酶部分失活,因此在pH值較低和較高的情況下,木聚糖酶的相對酶活都會下降。在pH值為3~5時,木聚糖酶的相對酶活在85.0%以上,而pH值為9~11時,木聚糖酶的相對酶活均<75.0%,這說明該木聚糖酶較適合在酸性環境中使用。
木聚糖酶的pH穩定性結果如圖9所示,該木聚糖酶在pH值為7時孵育60 min后,其相對酶活為98.5%,說明,該木聚糖酶在中性環境中較穩定。而在偏酸和偏堿的環境中,將該木聚糖酶孵育60 min后,相對酶活均會下降,并且在堿性環境中酶活下降幅度比在酸性環境中大,這也充分說明了該木聚糖酶對酸的耐受性比對堿的耐受性強。
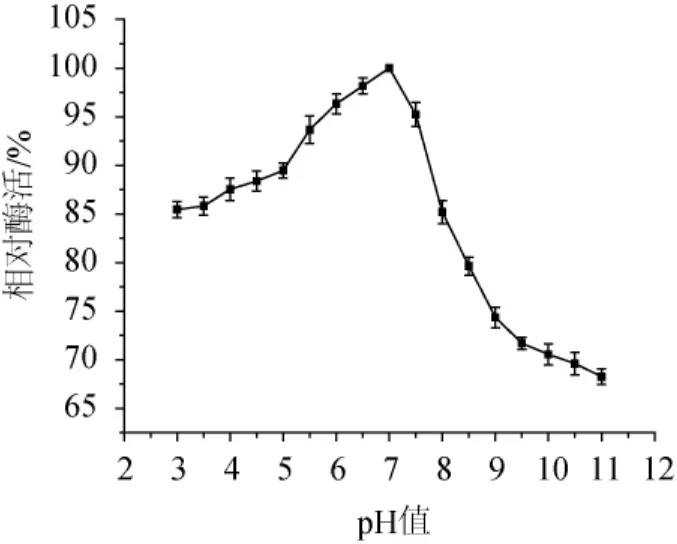
圖8 菌株ARTP-82產木聚糖酶的最適反應pH值Fig.8 Optimum reaction pH of xylanase produced by strain ARTP-82
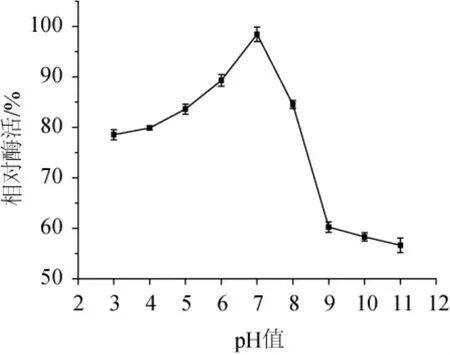
圖9 菌株ARTP-82產木聚糖酶的pH穩定性Fig.9 pH stability of xylanase produced by strain ARTP-82
3 結論
本研究對黑曲霉FXY進行ARTP誘變,分別對出發菌株的培養時間和誘變處理時間進行了考察,結果表明,ARTP誘變的最優條件為:菌株培養時間4 d,誘變處理時間5.0 min,在該條件下,篩得的優勢且能穩定遺傳的菌株ARTP-82所產木聚糖酶的酶活為175.33 U/mL,較出發菌株提高了120.8%,這表明黑曲霉對ARTP敏感,誘變效果好。本研究證實了ARTP誘變技術的突變率高,對選育木聚糖酶高產菌的可行性和有效性。為了探究誘變菌株木聚糖酶酶活提高的原因,對優勢菌株ARTP-82和出發菌株FXY的生長和發酵產酶情況作了對比研究,并得出結論,誘變菌株酶活之所以提高,不是因為菌株生物量的增加,而是與酶比活增加有關。通過對優勢菌株ARTP-82所產木聚糖酶酶學性質的研究得知該木聚糖酶的最適反應溫度為45℃、最適反應pH值為7,并且在該溫度和pH值條件下,酶的穩定性良好。本研究可為其他菌株的ARTP誘變提供有益的參考,對木聚糖酶的工業化生產也具有一定的借鑒意義。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
