抗草甘膦轉(zhuǎn)基因大豆AG5601對根際微生物群落功能多樣性的影響
2019-07-29 02:50:30張卓劉茂炎王培黃文坤劉二明彭煥彭德良
生物技術(shù)通報 2019年7期
張卓 劉茂炎 王培 黃文坤 劉二明 彭煥 彭德良
(1. 中國農(nóng)業(yè)科學院植物保護研究所植物病蟲害生物學國家重點實驗室,北京 100193;2. 湖南省植物保護研究所 園藝作物病蟲害治理湖南省重點實驗室,長沙 410125;3. 湖南農(nóng)業(yè)大學植物保護學院,長沙 410128)
自1994年美國獲準抗草甘膦除草劑轉(zhuǎn)基因大豆商業(yè)化種植以來,種植面積逐年增加,2017年全球種植面積達9 410萬hm2,占所有轉(zhuǎn)基因作物種植面積的50%[1]。隨著轉(zhuǎn)基因作物商業(yè)化生產(chǎn)步驟的加快,轉(zhuǎn)基因作物的風險評價備受關(guān)注[2-5]。轉(zhuǎn)基因作物對土壤生態(tài)環(huán)境和微生物群落結(jié)構(gòu)、功能及多樣性的影響是其安全性評價的一個重要內(nèi)容[6],土壤-微生物-作物三者互作的特殊微生態(tài)系統(tǒng)組成根際環(huán)境,根際環(huán)境是作物根系營養(yǎng)成分吸收、新陳代謝和生長的場所,根際微生物廣泛存在大豆根際生境中,并對大豆生長產(chǎn)生多方面的影響[7]。轉(zhuǎn)基因作物種植后,其表達的外源基因產(chǎn)物進入到土壤后,可能與土壤微生物發(fā)生作用從而影響其活動和功能[8]。由于農(nóng)田中土壤微生物群落的組成和多樣性與作物的根際分泌物有關(guān),轉(zhuǎn)基因與非轉(zhuǎn)基因作物的根際分泌物的變化導致其對土壤微生物群落的影響有所不同[9]。因此,轉(zhuǎn)基因作物對土壤微生物的多樣性、結(jié)構(gòu)和功能的影響是作為評價轉(zhuǎn)基因作物安全的重要指標之一[10]。此外,轉(zhuǎn)基因作物殘茬對土壤生態(tài)系統(tǒng)、土壤特異生物功能團、土壤生物多樣性也可能產(chǎn)生一定的影響[11]。
BIOLOG是描述微生物群落功能多樣性變化的重要指標,它能代表實際土壤微生物群體對碳源利用的動力學特征[12]。研究發(fā)現(xiàn)轉(zhuǎn)基因大豆和轉(zhuǎn)基因玉米根際微生物群落功能多樣性與受體的相比無顯著性差異[13-15],但轉(zhuǎn)Bt基因水稻SHK601與受體水稻相比對根際微生物群落功能有短暫影響[16]。不同耐草甘膦轉(zhuǎn)基因大豆對根際土壤固氮細菌多樣性的影響研究發(fā)現(xiàn),耐草甘膦轉(zhuǎn)基因大豆M88、GTS40-3-2 和非轉(zhuǎn)基因大豆中黃13 根際土壤固氮細菌多樣性指數(shù)和均勻度指數(shù)無顯著差異;抗蟲耐草甘膦轉(zhuǎn)基因大豆ZB 根際土壤固氮細菌多樣性指數(shù)顯著高于非轉(zhuǎn)基因大豆中黃13,均勻度指數(shù)無顯著差異[15]。轉(zhuǎn)G10-EPSPS基因抗草甘膦大豆SHZD32-01和配施草甘膦處理后,顯著影響結(jié)莢期根際土壤細菌和根瘤菌的多樣性,但其影響隨大豆的生長而逐漸消失[17]。王麗娟等[18]研究發(fā)現(xiàn),轉(zhuǎn)基因大豆PAT和ALS根際土壤微生物活性均高于相應(yīng)親本,兩種轉(zhuǎn)基因大豆根際土壤微生物群落物種均一度和優(yōu)勢度指數(shù)較對應(yīng)親本均無顯著差異,主成分分析發(fā)現(xiàn),非轉(zhuǎn)基因親本大豆ALS1 以及中黃13 土壤微生物群落碳源利用類型相似,僅轉(zhuǎn)基因大豆ALS 土壤微生物碳源利用類型表現(xiàn)出差異。
不同轉(zhuǎn)基因作物品種對土壤微生物群落的影響有一定的差異,基于轉(zhuǎn)基因作物安全性評價的個案原則開展相關(guān)研究是其安全性評價的必要步驟。本研究以抗草甘膦轉(zhuǎn)基因大豆AG5601、親本SNK500及常規(guī)大豆中黃13為材料,采用BIOLOG-ECO技術(shù),對成熟期和收割60 d后的根際殘茬根際微生物群落功能多樣性的差異進行比較分析。其結(jié)果有助于解析抗草甘膦轉(zhuǎn)基因大豆AG5601對根際微生物群落功能的影響及其對碳源利用的情況,為抗草甘膦轉(zhuǎn)基因大豆安全性評價提供參考。
1 材料與方法
1.1 材料
1.1.1 供試大豆品種 抗草甘膦轉(zhuǎn)基因大豆AG5601和親本非轉(zhuǎn)基因大豆SNK500由美國孟山都公司提供,當?shù)爻R?guī)品種中黃13由中國農(nóng)業(yè)科學院廊坊科研中試驗基地提供。
1.1.2 試驗地點和氣候條件 本試驗于2016年在河北省廊坊市安次區(qū)炊莊中國農(nóng)業(yè)科學院廊坊科研中試驗基地進行,試驗期間5-10月的月平均氣溫為20.6、23.7、27.4、25.0、19.4及14.5℃,平均降雨量 為 50.0 mm、67.9 mm、75.2 mm、75.2 mm、74.3mm及68.0 mm。
參照轉(zhuǎn)基因植物及其產(chǎn)品環(huán)境安全檢測 耐除草劑大豆第4部分:“生物多樣性影響”(中華人民共和國農(nóng)業(yè)部公告第2031號公告),試驗設(shè)3個處理分別為抗草甘膦轉(zhuǎn)基因大豆AG5601、親本大豆SNK500和當?shù)爻R?guī)品種中黃13,每個處理4次重復,共12個小區(qū)。小區(qū)采用隨機排列,小區(qū)間設(shè)有1.5 m寬的空白隔離帶,小區(qū)面積約150 m2(長11.5 m,寬13.0 m)。2016年5月25日播種,試驗小區(qū)常規(guī)管理,未施用化肥和農(nóng)藥,成熟期土壤的部分化學和生物學性狀如表1所示。
1.2 方法
1.2.1 土壤樣品采集 于大豆成熟期和收割后60 d殘茬根部取樣,每小區(qū)隨機取10株,用小鏟子挖出根部,輕輕抖掉根系外圍土,用毛刷輕刷黏附在根表面的土壤作為根際土,用無菌的封口袋密封并充分混勻,過50目篩,作為1個土壤樣品。

表1 供試土壤的化學和生物學性狀
1.2.2 BIOLOG-ECO分析 土壤樣品在25℃條件下活化24 h,取3 g土放入27 mL 的0.85 mol/L NaC1溶液中,振蕩30 min后,吸取3 mL上清,加入27 mL NaC1溶液,混勻后再吸取3 mL上清,加入27 mL NaC1溶液;最終的稀釋比例為1∶1 000。向ECO板的各孔中加入150 μL的稀釋液,每個土樣3次重復,將接種好的微孔板放在保濕的容器中,并放入25℃的恒溫培養(yǎng)箱中。分別于0、12、24、36、48、60、72、84、96、108、120、132、144、156、168、180、192、204、216、228和 240 h在 ELxS08 Biolog微孔板讀數(shù)儀(BIO-TEK Instruments INC,USA)上進行讀數(shù)并記錄。
1.2.3 數(shù)據(jù)分析 利用Excel對ECO板結(jié)果進行統(tǒng)計分析。單孔的平均顏色變化率(AWCD)計算方法:

式中,C為處理每個碳源孔的光密度值;R為對照孔的光密度值;n為培養(yǎng)基碳源種類數(shù)。
土壤微生物功能多樣性采用BIOLOG 培養(yǎng)72 h的數(shù)據(jù)進行分析,以Simpson,Shannon和 McIntosh3種指數(shù)來表征土壤微生物群落功能多樣性。
Shannon指數(shù)H 用來估算群落多樣性的高低,計算公式如下:

其中pi為第i孔的相對吸光值(Ci-R)與整個平板相對吸光值總和的比率
Simpson指數(shù)D,用于評估某些常見種的優(yōu)勢度,計算公式如下:

其中ni為第i孔的相對吸光值(Ci-R),N為相對吸光值總和;
Simpson指示用1/D值表示,
McIntosh指數(shù)U,反應(yīng)群落物種均一性,

ni同上。
采用SPSS16.0軟件進行方差分析和主成分分析,土壤理化性質(zhì)和微生物多樣性指數(shù)的顯著性分析采用單因素方差分析,對微生物碳源利用進行主成分分析。

圖1 AG5601,SNK500和中黃13根際微生物在BIOLOG-ECO板上的平均顏色變化率
2 結(jié)果
2.1 不同處理根際土壤微生物群落平均吸光值的變化
由圖1可以看出,AG5601成熟期的根際微生物群落AWCD在整個培養(yǎng)期內(nèi)均顯著高于親本SNK500,培養(yǎng)72 h 后高于常規(guī)品種中黃13,但差異不顯著;說明AG5601根際微生物的生理活性顯著高于親本SNK500,但與中黃13根際微生物的生理活性相比無顯著性差異。殘茬期,AG5601、SNK500和中黃13大豆根際微生物群落AWCD隨著培養(yǎng)時間的延長,微生物利用碳源量逐漸增加,非轉(zhuǎn)基因大豆SNK500和中黃13大豆根際微生物群落利用碳源量在144 h 后略高于AG5601,在整個培養(yǎng)過程中無顯著性差異。
在成熟期,AG5601與親本SNK500根際土壤微生物群落在McIntosh指數(shù)和Simpson指數(shù)差異顯著,AG5601、SNK500和中黃13根際微生物群落在Shannon指數(shù)無顯著性差異。AG5601與SNK500殘茬的根際土壤微生物群落的Shannon指數(shù)、McIntosh指數(shù)和Simpson指數(shù)均無顯著差異,但與中黃13在McIntosh指數(shù)上差異顯著(表2)。
2.2 微生物群落功能主成分分析
主成分分析發(fā)現(xiàn),AG5601、SNK500和中黃13成熟期的根際微生物群落在第1主成分上的得分沒有顯著性差異,AG5601在第2主成分上的得分與中黃13差異顯著,但與親本SNK500不顯著(圖2)。第1、2主成分貢獻率分別為81.88%和10.28%,累積貢獻率為92.16%,說明第1主成分是變異的主要來源,起主要參考作用。從31種碳源在2個主成分上的載荷值可知,除2-羥基苯甲酸外,其余30種碳源均集中在第1主成分上,決定了主成分1的分異,而影響主成分2的主要碳源主要是2-羥基苯甲酸、丙酮酸甲酯、α-環(huán)糊精、L-蘇氨酸、D-葡萄胺酸、甘油(表3)。綜合第1、2主成分分析結(jié)果:被AG5601和SNK500根際微生物群落特異利用的碳源有:D-半乳糖酸-γ-內(nèi)酯、L-精氨酸、D-木糖、L-天冬酰胺酸、I-赤蘚糖醇、α-環(huán)糊精、L-蘇氨酸、D-葡萄胺酸、衣康酸、葡萄糖-L-谷氨酸、葡萄糖- L-磷酸、α-丁酮酸、a-D-乳糖。被中黃13特異利用的碳源有:β-甲基-D-葡萄苷、丙酮酸甲酯、D-半乳糖醛酸、土溫-40、2-羥基苯甲酸、L-苯甲基丙氨酸、土溫-80、D-甘露醇、4-羥基苯甲酸、L-絲氨酸、N-乙酰基-D-葡萄胺、γ-羥基丁酸、肝糖、D-纖維二糖、苯基乙胺、甘油、D-蘋果酸、腐胺。說明在AG5601和SNK500成熟期,根際微生物在碳源的利用上保持一致。
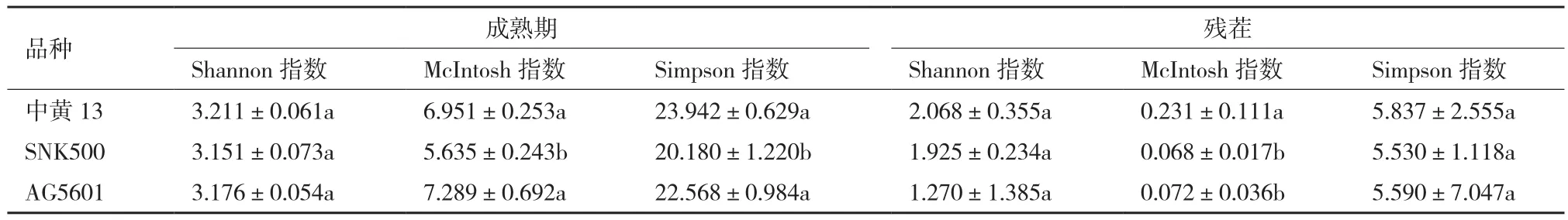
表2 轉(zhuǎn)基因與非轉(zhuǎn)基因大豆土壤微生物群落功能多樣性

圖2 土壤微生物碳源利用類型主成分分析(72 h)
主成分分析發(fā)現(xiàn),AG5601、SNK500和中黃13殘茬的根際微生物群落在第1主成分上的得分基本一致,AG5601在第2主成分上的得分與SNK500差異不顯著,與中黃13差異顯著(圖2)。第1、2主成分貢獻率分別為65.92%和25.21%,累積貢獻率為91.13%,說明第1主成分是變異的主要來源,起主要參考作用。從31種碳源在2個主成分上的載荷值可見(表4),除D-半乳糖醛酸、2-羥基苯甲酸、D-甘露醇、4-羥基苯甲酸、N-乙酰基-D-葡萄胺、D-葡萄胺酸不被根際微生物利用外,其他的25種碳源均被根際微生物利用(表4)。再從25種碳源在2個主成分上的載荷值可以看出,除L-天冬酰胺酸、腐胺外,其余23種碳源均集中在第1主成分上,決定了主成分1的分異,而影響主成分2的主要碳源主要是L-天冬酰胺酸、腐胺、L-精氨酸、葡萄糖-L-谷氨酸、苯基乙胺。綜合第1、2主成分結(jié)果,被AG5601和SNK500根際微生物群落特異利用的碳源有:L-精氨酸、丙酮酸甲酯、D-木糖、L-天冬酰胺酸、i-赤蘚糖醇、L-苯甲基丙氨酸、L-絲氨酸、γ-羥基丁酸、L-蘇氨酸、衣康酸、葡萄糖-1-磷酸、α-丁酮酸、苯基乙胺、甘油。被中黃13根際微生物群落特異利用的碳源有:β-甲基-D-葡萄苷、D-半乳糖酸-γ-內(nèi)酯、土溫-40、土溫-80、α-環(huán)糊精、肝糖、葡萄糖-L-谷氨酸、D-纖維二糖、α-D-乳糖、D-蘋果酸、腐胺。其他碳源則可被3種大豆的根際微生物群落共同利用或均不利用。說明AG5601和SNK500殘茬的根際微生物群落在碳源的利用上亦保持一致。

表3 ECO板上31種碳源在第一和第二主成分上的載荷值
3 討論
土壤根際微生物群落碳源的利用量代表著土壤中可培養(yǎng)微生物群落的數(shù)量。土壤微生物的數(shù)量是對土壤生態(tài)條件的綜合反應(yīng),土壤微生物中的細菌、真菌、放線菌具有不同的生活習性,它們的數(shù)量變化和在土壤微生物中所占的比例對其生活的土壤生態(tài)條件具有一定的指示意義[19]。根際是植物與土壤生態(tài)系統(tǒng)物質(zhì)交換的活躍界面,植物同化大氣中的CO2,將部分光合產(chǎn)物轉(zhuǎn)運至地下,促進土壤微生物的生長和新陳代謝;土壤中的微生物則將土壤中的有機態(tài)養(yǎng)分轉(zhuǎn)化成無機形態(tài),供給植物吸收利用[20]。

表4 轉(zhuǎn)基因大豆殘茬期ECO板上31種碳源在第一和第二主成分上的載荷值
近年來隨著新轉(zhuǎn)基因作物的出現(xiàn),轉(zhuǎn)基因作物的種植對根際微生物群落結(jié)構(gòu)和功能的影響受國內(nèi)外學者的普遍關(guān)注[4-6]。土壤微生物作為評價轉(zhuǎn)基因作物安全的重要指標,其群落結(jié)構(gòu)多樣性直觀地反映轉(zhuǎn)基因作物對土壤生態(tài)環(huán)境的影響[10]。已有研究結(jié)果表明,抗草甘膦轉(zhuǎn)基因大豆與常規(guī)大豆根系分泌物相比,表明前者根系分泌物含有的氨基酸和碳水化合物的量更多,進而對根際微生物產(chǎn)生一定影響[21]。本研究結(jié)果表明,AG5601與親本大豆SNK500成熟期的根際微生物群落的光密度差異顯著,而與中黃13根際微生物光密度不顯著,AG5601、SNK500和中黃13殘茬的根際微生物群落的光密度差異無顯著性差異。和以往的研究結(jié)果相比,不同時期轉(zhuǎn)基因作物根系對土壤微生物的影響有所不同。呂曉波等[21]經(jīng)過連續(xù)3年的大田和盆栽試驗,發(fā)現(xiàn)轉(zhuǎn)基因大豆與親本相比,在不同生育期根際土壤細菌和放線菌均表現(xiàn)降低趨勢。葉飛等[23]研究發(fā)現(xiàn),在棉花的整個生育期土壤微生物群落的光密度差異不顯著,花鈴期轉(zhuǎn)基因棉花根際土壤微生物群落的光密度均低于對照。
研究根際微生物群落對不同種類碳源利用率的差異,能夠更全面地了解微生物群落代謝功能特征。本研究結(jié)果表明,在成熟期AG5601與SNK500根際土壤微生物McIntosh指數(shù)、Simpson多樣性指數(shù)均顯著高于中黃13,而AG5601、SNK500和中黃13殘茬的根際土壤微生物多樣性差異不顯著。王麗娟等[18]研究表明,轉(zhuǎn)基因大豆ALS 顯著提高了根際土壤微生物的物種香農(nóng)-維納指數(shù)。但并對土壤微生物群落的物種均一度和優(yōu)勢度未產(chǎn)生顯著影響。轉(zhuǎn)基因大豆PAT 的種植對根際土壤微生物的物種香農(nóng)-維納指數(shù)、物種均一度和優(yōu)勢度指數(shù)均未產(chǎn)生顯著影響。梁晉剛等[14]研究表明,轉(zhuǎn)基因高蛋氨酸大豆及其受體大豆在豐富度指數(shù)、多樣性指數(shù)和均勻度指數(shù)上均沒有顯著性差異。轉(zhuǎn)基因抗蟲棉SGK321 及其親本棉石遠321不同生長時期其根際土壤微生物群落功能多樣性的變化研究表明,播種后30 d后,轉(zhuǎn)雙價棉根際土壤微生物群落豐富度指數(shù)和優(yōu)勢度指數(shù)顯著高于親本常規(guī)棉,而 90 d后,轉(zhuǎn)基因抗蟲棉根際土壤微生物群落豐富度指數(shù)和優(yōu)勢度指數(shù)則顯著低于親本常規(guī)棉[24]。以上結(jié)果表明,土壤微生物群落豐富度和優(yōu)勢度隨轉(zhuǎn)基因作物生長時期的不同而異,但轉(zhuǎn)基因作物的對土壤微生物多樣性無顯著影響。
Garland 等[25]研究結(jié)果表明,主成分分析結(jié)果中各處理在空間上位置與微生物在碳源的利用能力密切相關(guān)。本研究主成分分析結(jié)果表明,AG5601、SNK500和中黃13根際微生物群落在第1主成分上的得分基本一致,說明轉(zhuǎn)基因大豆AG5601和非轉(zhuǎn)基因大豆SNK500和中黃13根際在碳源的利用上亦保持一致。王麗娟等[18]研究表明,轉(zhuǎn)基因大豆PAT、親本PAT1、非轉(zhuǎn)基因親本大豆ALS1 以及中黃13 土壤微生物群落碳源利用類型相似,僅轉(zhuǎn)基因大豆ALS 土壤微生物碳源利用類型表現(xiàn)出差異。本研究結(jié)果表明,轉(zhuǎn)基因大豆AG5601種植后,對于微生物多樣性無顯著性影響,因此,轉(zhuǎn)基因大豆和常規(guī)品種種植后,根際土壤中微生物碳源利用上趨于穩(wěn)定。
近些年來用新方法分析土壤微生物群落取得了很大的進步,但是這些方法對土壤微生物群落的全面了解具有局限性[26]。而底物碳源的多樣性為微生物的生長提供了廣泛的選擇,從而使大多數(shù)可培養(yǎng)微生物均可在有31種碳源的微孔板上進行生命活動,并使微孔板發(fā)生顏色變化。土壤生態(tài)環(huán)境中微生物活性及群落多樣性受許多環(huán)境因子的影響,如土壤質(zhì)地結(jié)構(gòu)、土壤肥力、碳源、pH值、水分及植被類別等。而植物根系分泌的有機化合物是影響不同植物根際微生物多樣性的關(guān)鍵性因素[27]。由于土壤微生物受到的影響因子太多、太復雜,因此轉(zhuǎn)基因大豆影響土壤微生物的內(nèi)在機制及多種評價指標有待于進一步的研究。
4 結(jié)論
本研究發(fā)現(xiàn),雖在成熟期AG5601根際微生物群落利用碳源高于SNK500,但AG5601殘茬的根際微生物群落的光密度低于SNK500和中黃13,說明轉(zhuǎn)基因大豆AG5601根系對土壤微生物群落活性的影響是短暫的。主成分分析發(fā)現(xiàn),AG5601與SNK500在成熟期及收獲后的殘茬根際土壤微生物的碳源利用模式無差別,綜上所述,與對照相比,抗草甘膦轉(zhuǎn)基因大豆AG5601對根際微生物群落功能多樣性無顯著性影響。
猜你喜歡
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
音樂探索(2022年2期)2022-05-30 21:01:37
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
鄭州大學學報(醫(yī)學版)(2015年2期)2015-02-27 14:50:46
