飼糧α-亞麻酸水平對意大利蜜蜂工蜂幼蟲生理機能的影響
2019-07-29 03:24:56于靜張衛星馬蘭婷胥保華
中國農業科學 2019年13期
關鍵詞:水平
于靜,張衛星,馬蘭婷,胥保華
飼糧-亞麻酸水平對意大利蜜蜂工蜂幼蟲生理機能的影響
于靜,張衛星,馬蘭婷,胥保華
(山東農業大學動物科技學院,山東泰安 271018)
【】探究飼糧中-亞麻酸的添加水平對意大利蜜蜂()工蜂幼蟲抗氧化活性和免疫能力的影響。移取1日齡意大利蜜蜂工蜂幼蟲1 200只,隨機分為5組,每組5個重復,每個重復48只;其中1組為對照組,飼喂不添加-亞麻酸的基礎飼糧,4組為處理組,分別飼喂-亞麻酸添加水平為0.02%、0.04%、0.06%和0.08%的飼糧。按照室內蜜蜂幼蟲飼養方法,將1日齡幼蟲用移蟲針移至溫度適宜的加入200 μL飼糧的24孔細胞培養板內,培養板置于恒溫培養箱中(溫度33℃,相對濕度55%),試驗期間每天更換飼糧。飼養至第6天末或第7天初,幼蟲開始有直立或排便現象時,將幼蟲轉移至提前鋪好滅菌紙的24孔細胞培養板內準備化蛹。從飼養第1天開始,每天檢查并記錄幼蟲和蛹的死亡數量,并將死亡個體及時移除,直至未死亡的蛹全部羽化新蜂,記錄成功化蛹和羽化新蜂個體數量,統計幼蟲化蛹率和羽化率。各組分別取5、6和7日齡幼蟲測定抗氧化、免疫、脂質代謝指標及相關基因表達量。飼糧中-亞麻酸的添加水平為0.02%和0.04%時,化蛹率和羽化率顯著高于與其他處理組(<0.05),而工蜂幼蟲血淋巴中甘油三酯(TG)、總膽固醇(TC)和低密度脂蛋白(LDL)含量顯著低于對照組,高密度脂蛋白(HDL)的含量卻顯著高于對照組(<0.05)。飼糧中-亞麻酸的添加水平為0.04%時,工蜂幼蟲超氧化物歧化酶(T-SOD)的活性較對照組顯著增加,丙二醛(MDA)的含量顯著降低(<0.05)。飼糧中-亞麻酸的添加水平為0.02%、0.04%和0.06%時,6日齡工蜂幼蟲的溶菌酶(lysozyme)和酚氧化酶(PO)活性顯著高于對照組(<0.05)。飼糧中-亞麻酸添加水平為0.04%時,6日齡工蜂幼蟲脂肪酸合成酶(FAS)和乙酰輔酶A羧化酶(ACC)活性顯著低于對照組(<0.05)。飼糧中-亞麻酸添加水平為0.04%時,5和7日齡工蜂幼蟲和相對表達量顯著高于對照組,但飼糧中-亞麻酸添加水平為0.08%時,相對表達量會顯著降低(<0.05)。-亞麻酸對意大利蜜蜂工蜂幼蟲抗氧化活性和免疫能力有一定影響,幼蟲飼糧中-亞麻酸適宜添加水平為0.02%—0.04%。
意大利蜜蜂;工蜂;-亞麻酸;添加水平;抗氧化活性;免疫
0 引言
【研究意義】脂肪酸是細胞膜的主要成分,對細胞膜的功能起著重要的作用。脂肪酸也是蜜蜂繁殖和發育所必需的,是蜜蜂冬季能量和脂肪體發育的來源[1]。-亞麻酸是動物體內一種必需的不飽和脂肪酸,它是所有n-3多不飽和脂肪酸的前體物質,在體內可轉化為長鏈的n-3多不飽和脂肪酸DHA和EPA,從而發揮與DHA和EPA相似的生理功能,對機體生長發育和正常代謝起重要作用[2]。-亞麻酸能抑制脂肪酸合成酶、COA-羧化酶、二酰甘油乙酰轉移酶和3-羥基-3-甲基戊二酰輔酶A還原酶(3-hydroxy-3-methyl glutarylcoenzyme A reductase,HMGR)等活性,并可加強線粒體中的-氧化,減少甘油三酯和膽固醇的合成,從而起到降血脂作用。-亞麻酸通過抑制ω-6系PUFA的代謝,減少前列腺素PGE2、前列腺環素PGI2、血栓素TXA2的合成,從而產生抗炎、抗過敏、抗血栓等作用。蜜蜂幼蟲的主要脂肪酸來源是蜂王漿,蜂王漿中-亞麻酸含量極少,而-亞麻酸在體內不能合成,必須從食物中獲得,因此探究幼蟲飼糧中-亞麻酸適宜添加水平十分必要。【前人研究進展】大量研究表明,與哺乳動物相似,-亞麻酸等ω-3脂肪酸對蜜蜂的認知功能有著至關重要的作用[1],也有研究認為蜜蜂腸道中高濃度的亞麻酸和亞油酸具有抵抗真菌和細菌感染的作用[3]。Arien等[4]研究了-亞麻酸等ω-3脂肪酸缺乏對蜜蜂的營養影響,發現飼喂低-亞麻酸飼糧的蜜蜂嗅覺和觸覺聯想學習能力明顯下降;李興鵬等[5]研究發現,含有-亞麻酸等不飽和脂肪酸的人工飼料更適合蠋蝽()老齡若蟲生長發育,提高成蟲獲得率和種群增長率;姚思宇等[6]以不同劑量的-亞麻酸給小鼠連續灌胃30—35 d,發現小鼠的抗體生成細胞數和血清溶血素水平顯著提高,說明-亞麻酸具有增強小鼠免疫力的作用;朱保忠等[7]研究發現,-亞麻酸具有抗氧化和延長壽命的作用,而且-亞麻酸與抗氧化劑聯用能強化這種作用;馬蘭婷[8]研究表明,代用花粉中-亞麻酸水平能夠影響蜜蜂血淋巴中脂質含量、蜂體脂肪酶活性及蜂體相關基因表達水平等,但在代用花粉中添加過高或過低的-亞麻酸都會降低意大利蜜蜂()的壽命。【本研究切入點】目前,關于-亞麻酸對意大利蜜蜂營養作用的研究較少,而其幼蟲對-亞麻酸的營養需要量未見報道。【擬解決的關鍵問題】以人工飼養的意大利蜜蜂工蜂幼蟲為研究對象,通過探究幼蟲飼糧中不同-亞麻酸水平對其生長發育、抗氧化活性和免疫指標的影響,明確工蜂幼蟲對-亞麻酸的適宜需要量,為制定蜜蜂飼養標準提供依據。
1 材料與方法
試驗于2018—2019年在山東農業大學動物科技學院完成。
1.1 材料
意大利蜜蜂工蜂幼蟲取自山東農業大學試驗蜂群。對照組飼糧參照Vandenberg等[9]的配方配制,在基礎飼糧中分別添加0.02%、0.04%、0.06%、0.08%的-亞麻酸,制成4種試驗組飼糧(表1)。

表1 試驗飼糧組成
主要試劑:Total RNA Kit II試劑盒購自OMEGA公司,熒光定量試劑盒購自TransGen公司,Transcript First Strand cDNA Synthesis Kit試劑盒購自Roche公司,昆蟲酶聯免疫吸附試驗(ELISA)試劑盒購自科諾迪生物科技有限公司,-亞麻酸(濃度80%以上)購自江蘇天凱生物有限公司。
1.2 試驗方法
首先用蜂王隔離器將蜂王固定在一張巢脾上產卵,12 h內產完足夠多的卵時,標記產卵區,釋放蜂王,經過3 d,工蜂卵孵化為小幼蟲,即1日齡幼蟲。試驗選取1 200只1日齡意大利蜜蜂工蜂幼蟲,600只用于幼蟲化蛹率和羽化率指標測定,另外600只用于生理生化指標測定。試驗幼蟲均按單因素隨機設計分為5組,每組5個重復,每個重復48只。將1日齡幼蟲用移蟲針移至溫度適宜的加入200 μL飼糧的24孔細胞培養板內,培養板置于恒溫培養箱中(溫度33℃,相對濕度55%),試驗期間每天更換飼糧。飼養至第6天末或第7天初,幼蟲開始有直立或排便現象時,將幼蟲轉移至提前鋪好滅菌紙的24孔細胞培養板內準備化蛹,具體飼養方法參照王穎[10]。
1.3 測定指標與方法
1.3.1 化蛹率和羽化率測定 從飼養第1天開始,每天檢查并記錄幼蟲的死亡數量,并將死亡個體及時移除,直至未死亡的幼蟲全部化蛹并羽化成新蜂,記錄成功化蛹個體數量和羽化新蜂個體數量,統計幼蟲化蛹率和羽化率。
1.3.2 粗脂肪含量測定 幼蟲粗脂肪含量測定采用王少梅等[11]氯仿-甲醇抽提法。分別取5、7日齡工蜂幼蟲樣品10只,放在提前烘干稱重的稱量瓶中,置于65℃恒溫干燥箱中12 h,將烘干樣品置于10 ml離心管中,加入2 ml氯仿-甲醇(2﹕1)混合液,用全自動樣品快速研磨儀勻漿3 min,靜置24 h。向離心管中加入2 ml氯仿-甲醇(2﹕1)混合液,3 000 r/min離心10 min,將上清移至另一離心管中,向殘渣中加入2 ml氯仿-甲醇(2﹕1)混合液,同樣條件離心,將上清轉移另一離心管中;向上清離心管中加入1.2 ml 1.6% CaCl2溶液,搖勻,靜置1 h,同樣條件離心,吸去上層液;將1 ml 2% CaCl2-氯仿-甲醇(3﹕8﹕4)混合液上層液沿壁緩緩加入離心管中,同樣條件離心,吸去上層液;下層液轉入已烘干稱重的稱量瓶中,70℃恒溫烘干稱重,稱量瓶兩次差值即為粗脂重。
1.3.3 幼蟲血淋巴脂質含量測定 采用體積為40 μl的毛細血管吸取7日齡工蜂幼蟲血淋巴,用吸耳球將血淋巴吹入已經加有少許苯基硫脲的1.5 ml離心管中,試驗過程中離心管放在冰盒上,以防血淋巴被氧化,樣品放在-80℃保存。測定時,4℃條件下13 000 r/min離心5 min,取100 μl上清備用,采用日立7020型全自動生化分析儀測定幼蟲血淋巴中甘油三酯、膽固醇、高密度脂蛋白和低密度脂蛋白含量。
1.3.4 幼蟲抗氧化指標和脂質代謝、免疫相關酶活性測定 每個重復隨機選取1只6日齡工蜂幼蟲,按1﹕9質量體積比加入一定量生理鹽水于2 ml離心管中進行組織勻漿,以配制成10%勻漿液,在4℃條件下,13 000 r/min離心10 min,取100 μl上清備用。采用昆蟲酶聯免疫吸附試驗(ELISA)測定丙二醛(MDA)含量、總超氧化物歧化酶(T-SOD)、乙酰輔酶A羧化酶(ACC)、脂肪酸合成酶(FAS)、酚氧化酶(PO)和溶菌酶(lysozyme)活性。
1.3.5 免疫相關基因表達量的測定 使用Total RNA Kit II試劑盒提取5和7日齡工蜂幼蟲總RNA,并用超微量分光光度計檢測其濃度和純度。根據Transcript First Strand cDNA Synthesis Kit試劑盒,將總RNA反轉錄為cDNA,-20℃保存備用。內參基因選擇表達穩定的,委托生工生物工程(上海)股份有限公司完成定量引物的設計與合成,引物信息見表2。按照實時熒光定量試劑盒操作指南,用美國ABI7500系統檢測目的基因的相對表達量,反應體系為20 μL。反應條件:94℃30 s;94℃ 5 s,60℃34 s,40個循環。

表2 目的基因引物序列
1.4 數據分析
數據采用SAS 9.2軟件進行單因素方差分析(one-way ANOVA)和Turkey檢驗進行比較分析,<0.05為差異顯著,<0.01為差異極顯著,試驗數據均以“mean±SD”的形式表示。
2 結果
2.1 飼糧α-亞麻酸添加水平對意大利蜜蜂工蜂幼蟲化蛹率和羽化率的影響
飼糧中-亞麻酸的添加水平為0.02%和0.04%時,化蛹率和羽化率顯著高于其他處理組;-亞麻酸的添加水平為0.06%和0.08%時,化蛹率和羽化率均顯著低于對照組(<0.05)(表3)。
2.2 飼糧中α-亞麻酸添加水平對7日齡意大利蜜蜂工蜂幼蟲血淋巴生化指標的影響
飼糧中-亞麻酸添加水平為0.04%時,血淋巴中總蛋白(total protein,TP)含量顯著低于對照組,而飼糧中添加-亞麻酸會顯著降低工蜂幼蟲血淋巴中甘油三酯(triglyceride,TG)和總膽固醇(total cholesterol,TC)的含量(<0.05)。與對照組相比,飼糧中-亞麻酸添加水平為0.02%、0.04%和0.06%時,高密度脂蛋白(high density lipoprotein,HDL)的含量顯著升高,而低密度脂蛋白(low density lipoprotein,LDL)的含量顯著降低(<0.05)(表4)。

表3 飼糧中α-亞麻酸添加水平對工蜂幼蟲化蛹率和羽化率的影響
同行數據后不同小寫字母表示差異顯著(<0.05)。下同
Data with different lowercases in the same row indicate significant difference (<0.05). The same as below

表4 飼糧中α-亞麻酸添加水平對工蜂幼蟲血淋巴生化指標的影響
2.3 飼糧中α-亞麻酸添加水平對5、7日齡意大利蜜蜂工蜂幼蟲粗脂含量的影響
-亞麻酸添加水平為0.02%、0.06%和0.08%時,5日齡工蜂幼蟲蟲體粗脂含量顯著高于對照組;-亞麻酸添加水平為0.06%和0.08%時,7日齡工蜂幼蟲蟲體粗脂含量顯著高于對照組(<0.05)(表5)。
2.4 飼糧中α-亞麻酸添加水平對6日齡意大利蜜蜂工蜂幼蟲抗氧化指標的影響
與對照組相比,當飼糧中-亞麻酸添加水平為0.02%和0.04%時,6日齡工蜂幼蟲的丙二醛(MDA)含量顯著降低(<0.05)(圖1-A);飼糧中-亞麻酸添加水平為0.04%時,6日齡工蜂幼蟲超氧化物歧化酶(SOD)活性顯著高于對照組(<0.05)(圖1-B)。

表5 飼糧中α-亞麻酸添加水平對工蜂幼蟲粗脂含量的影響(干基,%)

柱上不同小寫字母表示差異顯著(P<0.05)。下同Different lowercases on the bars indicate significant difference (P<0.05). The same as below
2.5 飼糧中α-亞麻酸添加水平對意大利蜜蜂工蜂幼蟲脂質代謝相關酶活性的影響
飼糧中-亞麻酸添加水平為0.02%和0.04%時,意大利蜜蜂工蜂幼蟲體內乙酰輔酶A羧化酶(ACC)活性顯著低于對照組(<0.05)(圖2-A)。飼糧中-亞麻酸添加水平為0.02%時,工蜂幼蟲體內脂肪酸合成酶(FAS)活性與對照組無顯著差異(>0.05),但添加水平為0.04%、0.06%和0.08%時,FAS活性顯著低于對照組(<0.05)(圖2-B)。
2.6 飼糧中α-亞麻酸添加水平對意大利蜜蜂工蜂幼蟲免疫能力的影響
飼糧中-亞麻酸添加水平為0.02%、0.04%和0.06%時,6日齡工蜂幼蟲的溶菌酶(lysozyme)和酚氧化酶(PO)活性顯著高于對照組(<0.05)(圖3)。
飼糧中-亞麻酸添加水平為0.02%和0.04%時,5日齡工蜂幼蟲和相對表達量顯著高于對照組(<0.05),但當飼糧中-亞麻酸添加水平為0.06%和0.08%時,相對表達量會明顯降低(圖4-A)。與對照組相比,當飼糧中-亞麻酸添加水平為0.04%和0.06%時,7日齡工蜂幼蟲和相對表達量顯著增加,而當飼糧中-亞麻酸添加水平為0.08%時,e和相對表達量卻顯著降低(<0.05)(圖4-B)。

圖2 飼糧中α-亞麻酸添加水平對6日齡工蜂幼蟲脂質代謝相關酶活性的影響

圖3 6日齡工蜂幼蟲溶菌酶和酚氧化酶活性
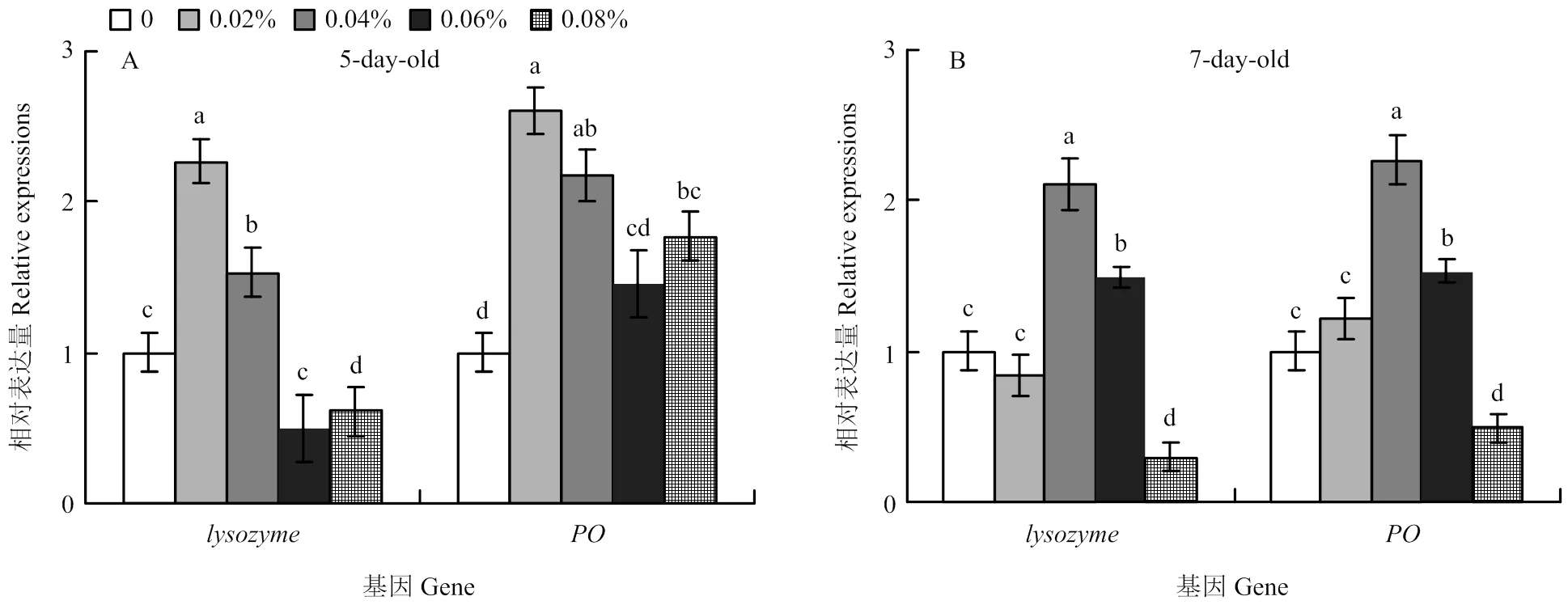
圖4 酚氧化酶和溶菌酶基因相對表達量
3 討論
3.1 飼糧中α-亞麻酸添加水平對意大利蜜蜂工蜂幼蟲化蛹率和羽化率的影響
-亞麻酸作為n-3系列不飽和脂肪酸,對昆蟲尤其蜜蜂生長性能的研究極少,但在畜禽上的研究較多。潘瑜等[12]研究表明,隨著亞麻油替代魚油水平的升高,鯉魚生長性能呈現出先升高而后降低的趨勢。也有大量研究表明,添加一定量的亞麻油對肉雞和蛋雞的生產性能無顯著影響[13-15]。杜海濤等[16]研究表明,低水平的-亞麻酸添加量對肉兔生長性能影響不顯著,而高水平的-亞麻酸添加量顯著降低肉兔的生長性能。本試驗表明,低水平的-亞麻酸添加量對意大利蜜蜂工蜂幼蟲的化蛹率和羽化率無顯著影響,而當飼糧中-亞麻酸添加水平達到0.06%時會顯著降低幼蟲的化蛹率和羽化率。李興鵬等[5]采用兩種分別添加棕櫚油和亞麻酸的飼料飼喂蠋蝽,發現取食添加亞麻酸飼料的蠋蝽老齡幼蟲的死亡率顯著降低,成蟲羽化率和雌雄成蟲體重日均增長率亦顯著高于取食棕櫚油飼料的蠋蝽,而添加棕櫚酸的飼料更適合低齡若蟲。由此可見,-亞麻酸對畜禽乃至蜜蜂生長性能的影響仍存在爭議,這可能與-亞麻酸的添加量和動物物種及其所處的生理階段有關。
3.2 飼糧中α-亞麻酸添加水平對意大利蜜蜂工蜂幼蟲脂質代謝的影響
3.2.1 對7日齡工蜂幼蟲血淋巴生化指標的影響 昆蟲血淋巴是血細胞、間質液和血漿的混合物,為昆蟲新陳代謝中物質交換與儲存的場所,在免疫、創傷、愈合、防御等方面有著重要作用[17]。總膽固醇、甘油三酯、高密度脂蛋白、低密度脂蛋白含量是臨床上用來反映血脂代謝功能的常用指標[18]。-亞麻酸可以降低肝臟膽固醇合成過程中的關鍵酶3-羥基-3-甲基戊二酰輔酶A還原酶的活性,從而降低肝臟膽固醇的合成量,同時也能增加血液高密度脂蛋白含量[19-20]。大量動物試驗和臨床試驗表明,多不飽和脂肪酸(PUFA)可以降低甘油三酯、膽固醇和低密度脂蛋白水平,升高高密度脂蛋白水平。許繼取[21]研究表明,增加-亞麻酸攝入的高脂飲食的大鼠血漿中甘油三酯、血漿總膽固醇水平均明顯降低;MEDEIROS等[22]證明-亞麻酸可顯著降低血清總膽固醇、甘油三酯、低密度脂蛋白膽固醇(LDL-C)水平;姚林杰等[18]研究表明,飼料中添加1.08%-亞麻酸能顯著降低團頭魴血清總膽固醇和甘油三酯含量,而高密度脂蛋白含量則顯著升高。
本研究表明,飼糧中添加-亞麻酸顯著降低意大利蜜蜂工蜂幼蟲血淋巴甘油三酯和總膽固醇的含量,0.02%和0.04%組總膽固醇含量顯著低于其他試驗組,-亞麻酸添加水平為0.02%、0.04%和0.06%時會顯著降低低密度脂蛋白含量,升高高密度脂蛋白的含量,0.04%組低密度脂蛋白含量顯著低于其他試驗組,而高密度脂蛋白的含量顯著高于其他組,這與上述研究結果相吻合,說明在適宜的添加量下,-亞麻酸具有降血脂的作用。
3.2.2 對6日齡工蜂幼蟲脂質代謝相關酶活性的影響-亞麻酸作為結構物質和代謝調控物質,發揮著重要的生理作用,而-亞麻酸的某些生理作用是通過調節相關酶的活性來實現的。-亞麻酸的降血脂作用一方面是通過對代謝率的調節來實現,另一方面則是通過抑制有關脂肪和甘油合成酶系來實現的。Mersmann等[23]通過在成年豬飼料中添加脂肪酸的研究發現,PUFA對脂肪酸的合成速度以及FAS和ACC的活性具有抑制作用;Toussant等[24]分別用富含PUFA和飽和脂肪酸的飼糧飼喂大鼠,結果發現飼喂富含PUFA飼糧的大鼠肝臟FAS和ACC活性顯著低于飼喂富含飽和脂肪酸飼糧的大鼠;Blake等[25]研究發現,飼喂PUFA的大鼠肝臟中FAS mRNA豐度是飼喂飽和脂肪酸大鼠的6%。
大量的研究結果表明,PUFA對脂肪合成酶系有抑制作用,這與本試驗結果相一致,飼糧中-亞麻酸添加水平為0.04%時,會顯著抑制FAS和ACC的活性,從而起到降血脂的作用。
3.3 飼糧中α-亞麻酸添加水平對意大利蜜蜂工蜂幼蟲抗氧化能力的影響
自由基通過其強氧化作用對核酸進行氧化和交聯,使DNA發生斷裂、突變,對機體的抗氧化系統和大分子產生損傷,損傷的積累導致細胞衰老或死亡[7]。超氧化物歧化酶(SOD)是生物體內重要的抗氧化酶,它能清除超氧陰離子自由基,對抗與阻斷因氧自由基對細胞造成的損害,并及時修復受損細胞,是體內抗氧化能力的總體體現。丙二醛(MDA)是膜脂過氧化的重要產物,其含量間接地反映機體細胞受自由基攻擊的嚴重程度[26]。葛雙雙等[27]研究表明,余甘子核仁油具有較強的抗氧化作用,而其主要的抗氧化活性物質為-亞麻酸,隨著余甘子核仁油-亞麻酸濃度的升高,對DPPH自由基和ABTS+·清除率也逐漸增大,抗氧化能力也逐漸增強;朱保忠等[7]研究表明,-亞麻酸能顯著提高動物血清SOD、谷胱甘肽過氧化物酶水平,降低MDA含量,提高動物抗自由基能力并延長壽命;張利華等[28]研究表明,飲食補充-亞麻酸可顯著增加糖尿病大鼠SOD和CAT活性,降低MDA含量,提高糖尿病大鼠的抗氧化能力。本研究表明,當-亞麻酸添加水平為0.02%和0.04%時顯著降低工蜂幼蟲MDA含量,而0.04%水平-亞麻酸能顯著增加SOD的活性。這與上述報道一致,說明飼糧中添加適宜的-亞麻酸能夠顯著提高幼蟲的抗氧化能力。
3.4 飼糧中α-亞麻酸添加水平對意大利蜜蜂工蜂幼蟲免疫能力的影響
蜜蜂血細胞總數被認為是間接衡量細胞免疫能力的指標[29],血細胞數目與包囊反應、酚氧化酶活力和擬寄生抗性具有正相關性,一般認為,血細胞數目減少是免疫能力降低的體現[30-32]。體液免疫會激活信號通路引起抗菌肽(antimicrobial peptide)和溶菌酶的合成,并激發酚氧化酶原級聯系統,它們能協同作用阻擋或殺死入侵微生物[33]。姚思宇等[6]研究表明,-亞麻酸能明顯促進小鼠的單核-巨噬細胞和腹腔巨噬細胞的吞噬能力,提高小鼠NK細胞的活性,最終能增強小鼠的免疫力;雙金等[34]研究表明,奶牛日糧中添加富含-亞麻酸的添加劑可明顯改善或增強奶牛的體液免疫和細胞免疫功能;Lessard等[35]通過分別飼喂奶牛富含n-6和n-3多不飽和脂肪酸的日糧,發現喂食富含n-3 PUFA飼糧的奶牛與喂食富含n-6 PUFA飼糧的奶牛相比,淋巴細胞增殖反應顯著增強。溶菌酶是巨噬細胞活化時釋放的一種水解酶C,可水解溶壁微球菌、巨大芽孢桿菌及黃色八疊球菌等革蘭氏陽性菌細胞壁上的糖苷鍵,使細菌溶解,溶菌酶含量可以評價動物機體的免疫狀態。陳士勇等[36]研究表明,日糧中添加PUFA可以改善產蛋雞體液免疫水平,提高抗體效價,但攝入過高的PUFA會顯著降低產蛋雞溶菌酶含量,從而降低了機體免疫反應。添加過高或過低的不飽和脂肪酸都會使免疫功能有所下降,只有適量的供給才能提高機體的免疫功能。
本研究表明,當飼糧中-亞麻酸的添加水平為0.02%、0.04%和0.06%時,可以顯著提高幼蟲溶菌酶和酚氧化酶活性。0.02%添加水平的-亞麻酸飼糧可以顯著提高5日齡幼蟲和相對表達量,而0.04%添加水平的-亞麻酸飼糧可以提高7日齡幼蟲和相對表達量,但當飼糧中-亞麻酸添加水平為0.08%時,和相對表達量卻顯著降低。由此可以說明,適宜的-亞麻酸可以增強蜜蜂的免疫力,但如果添加量過高則會造成蜜蜂的免疫力下降。
4 結論
意大利蜜蜂工蜂幼蟲飼糧中-亞麻酸添加水平對幼蟲的化蛹率、羽化率以及抗氧化、免疫能力和脂質代謝均有一定程度的影響;意大利蜜蜂工蜂幼蟲飼糧中-亞麻酸的添加水平為0.02%—0.04%時,幼蟲的化蛹率和羽化率有所提高,且有利于增加其抗氧化和免疫能力,為較為適宜的-亞麻酸添加水平。
[1] MANNING R. Fatty acids in pollen: a review of their importance for honey bees., 2001, 82(2): 60-75.
[2] 吳俏槿, 杜冰, 蔡尤林, 梁鉆好, 林志光, 邱國亮, 董立軍.-亞麻酸的生理功能及開發研究進展. 食品工業科技, 2016, 37(10): 386-390.
WU Q J, DU B, CAI Y L, LIANG Z H, LIN Z G, QIU G L, DONG L J. Research development of alpha-linolenic acid., 2016, 37(10): 386-390. (in Chinese)
[3] FELDLAUFER M F, KNOX D A, LUSBY W R, SHIMANUKI H. Antimicrobial activity of fatty acids against, the causative agent of American foulbrood disease., 1993, 24: 95-99.
[4] ARIEN Y, DAG A, ZARCHIN S, MASCI T, SHAFIR S. Omega-3 deficiency impairs honey bee learning., 2015, 112(51): 15761-15766.
[5] 李興鵬, 宋麗文, 陳越渠, 李亞男, 左彤彤, 武三安. 不同脂肪源人工飼料對蠋蝽生長發育及生殖力的影響. 林業科學, 2018, 54(6): 85-93.
LI X P, SONG L W, CHEN Y Q, LI Y N, ZUO T T, WU S A. Influence of different fatty acids in artificial diets on growth, development and fecundity of., 2018, 54(6): 85-93. (in Chinese)
[6] 姚思宇, 趙鵬, 李彬, 李風文, 馬宇燕, 覃輝艷, 張陸娟, 蘇愛榮, 梁慧莉.-亞麻酸對小鼠免疫功能影響的實驗研究. 中國熱帶醫學, 2007, 7(3): 334-349.
YAO S Y, ZHAO P, LI B, LI F W, MA Y Y, QIN H Y, ZHANG L J, SU A R, LIANG H L. Experimental study on the effect of linolenic acid on enhancement of mice’s immune function., 2007, 7(3): 334-349. (in Chinese)
[7] 朱保忠, 李琳.-亞麻酸與抗氧化劑聯用對果蠅壽命及小鼠抗氧化能力的影響. 中國組織工程研究與臨床康復, 2008, 12(7): 1264-1267.
ZHU B Z, LI L. Influence of alpha-linoleic acid plus anti-oxidant agent on drosophila life and mouse antioxidation ability., 2008, 12(7): 1264-1267. (in Chinese)
[8] 馬蘭婷. 代用花粉中-亞麻酸水平對蜜蜂采食量、群勢及脂質代謝的影響[D]. 泰安: 山東農業大學, 2013.
MA L T. Effect of dietary-linolenic acid level on intake, colony development and lipid metabolism of honey bee (L.)[D]. Taian: Shandong agricultural university, 2013. (in Chinese)
[9] VANDENBERG J D, SHIMANNUKI H. Technique for rearing worker honeybees in the laboratory., 1987, 26(2): 90-97.
[10] 王穎. 營養和空間因素對蜜蜂級型分化的影響[D]. 泰安: 山東農業大學, 2015.
WANG Y. Effects of nutritional factors and living space on the caste determination of honey bees[D]. Taian: Shandong agricultural university, 2015. (in Chinese)
[11] 王少梅, 陳少蓮, 崔奕波.用氯仿-甲醇抽提法測定魚體脂肪含量的研究. 水生生物學報, 1993, 17(2): 193-196.
WANG S M, CHEN S L, CUI Y B. Study on the procedures of chloroform-methanol extraction for the determination of lipid content of fish samples., 1993, 17(2): 193-196. (in Chinese)
[12] 潘瑜, 陳文燕, 林仕梅, 高啟平, 羅莉. 亞麻油替代魚油對鯉魚生長性能、肝胰臟脂質代謝及抗氧化能力的影響. 動物營養學報, 2014, 26(2): 420-426.
PAN Y, CHEN W Y, LIN S M, GAO Q P, LUO L. Effect of replacement of fish oil by linseed oil on growth performance, lipid metabolism and antioxidant ability in hepatopancreas of common carp () ., 2014, 26(2): 420-426. (in Chinese)
[13] 劉利曉. 富含n-3多不飽和脂肪酸雞肉的研究[D]. 武漢: 華中農業大學, 2007.
LIU L X. Study on producing of chicken meat enriched in n-3 polyunsaturated fatty acids[D]. Wuhan: Huazhong agricultural university, 2007. (in Chinese)
[14] 汪鯤. n-3多不飽和脂肪酸在蛋黃和組織中的富集規律及其對產蛋雞脂類代謝的影響[D]. 北京: 中國農業科學院, 2000.
WANG K. Enrichment of n-3 polyunsaturated fatty acid in egg yolk and tissue and its effect on lipid metabolism of laying hens[D]. Beijing: Chinese academy of agricultural sciences, 2000. (in Chinese)
[15] LOPEZ-Ferrer S, BAUCELLS M D, BARROETA A C, GALOBART J, GRASHORN M A. n-3 enrichment of chicken meat. 2. Use of precursors of long-chain polyunsaturated fatty acids: Linseed oil., 2001, 80(6): 753-761.
[16] 杜海濤, 王春陽, 王雪鵬, 麻名文, 李福昌. 日糧-亞麻酸水平對斷奶至2月齡肉兔生長性能、脂肪酸構成及肝臟相關基因mRNA表達的影響. 畜牧獸醫學報, 2011, 42(5): 671-678.
DU H T, WANG C Y, WANG X P, MA M W, LI F C. Effect of-linolenic acid in diets on growth performance, fatty acids composition of tissues andl mRNA expression of liver related genes in weaned to 2-month-old rabbits., 2011, 42(5): 671-678. (in Chinese)
[17] 張月月, 馬振剛. 一種高效收集蜜蜂血淋巴的新方法. 中國蜂業, 2017, 68(6): 64.
ZHANG Y Y, MA Z G. A new method for efficient collection of haemolymph of bees., 2017, 68(6): 64. (in Chinese)
[18] 姚林杰, 葉元土, 蔡春芳, 許凡, 劉猛, 劉漢超, 董嬌嬌, 陳科全, 黃雨薇.團頭魴幼魚飼料中-亞麻酸、亞油酸的適宜含量. 動物營養學報, 2015, 27(3): 766-774.
YAO L J, YE Y T, CAI C F, Xu F, LIU M, LIU H C, DONG J J, CHEN K Q, HUANG Y W. Optimal dietary-linolenic acid and linoleic acid contents of blunt snout bream () fingerlings., 2015, 27(3): 766-774. (in Chinese)
[19] SANGIOVANNI J P, CHEW E Y. The role of omega-3 long-chain polyunsaturated fatty acids in health and disease of the retina., 2005, 24(1): 87-138. (in Chinese)
[20] 雙金, 黎明, 敖力格日瑪, 侯先志, 閆素梅. 亞麻籽對肉羊血清脂蛋白和脂肪代謝相關生化指標的影響. 動物營養學報, 2014, 26(4): 918-929.
SHUANG J, LI M, AO L, HOU X Z, YAN S M. Effects of flaxseed on serum biochemical indices related to lipoprotein and fat metabolism of meat sheep., 2014, 26(4): 918-929. (in Chinese)
[21] 許繼取. 亞麻酸對高脂大鼠血脂影響及促進肝臟SR-BI表達機制研究[D]. 武漢: 華中科技大學, 2006.
Xu J Q. Effect of-linolenic acid on plasma lipid of rats with high fat diet and mechanism of increasing hepatocyte SR-BI expression[D]. Wuhan: Huazhong university of science and technology, 2006. (in Chinese)
[22] MEDEIROS D M, HAMPTON M, KURTZER K, PARELMAN M, AL-TAMIMI E, DROUILLARD J S. Feeding enriched omega-3 fatty acid beef to rats increases omega-3 fatty acid content of heart and liver membranes and decreases serum vascular cell adhesion molecule-1 and cholesterol levels., 2007, 27(5): 295-299.
[23] MERSMANN H J, HOUK J M, PHINNEY G, UNDERWOOD M C, BROWN L J. Lipogenesis byliver and adipose tissue preparations from neonatal swine., 1973, 224(5): 1123-1129.
[24] TOUSSANT M J, WILSON M D, CLARKE S D. Coordinate suppression of liver acetyl-CoA carboxylase and fatty acid synthetase by polyunsaturated fat., 1981, 111(1): 146-153.
[25] BLAKE W L, CLARKE S D. Suppression of rat hepatic fatty acid synthase and S14gene transcription by dietary polyunsaturated fat., 1990, 120(12): 1727-1729.
[26] ZHANG L D, LI H C, CHONG T, GAO M, YIN J, FU D L, DENG Q, WANG Z M. Prepubertal exposure to genistein alleviates Di-(2-ethylhexyl) phthalate induced testicular oxidative stress in adult rats., 2014, 2014: Article ID 598630.
[27] 葛雙雙, 張雯雯, 李坤, 徐涓, 劉蘭香, 鄭華, 張弘. 余甘子核仁油的體外抗氧化活性及其作用機理. 食品科學, 2017, 38(15): 127-134.
GE S S, ZHANG W W, LI K, XU J, LIU L X, ZHENG H, ZHANG H. antioxidant activity and mechanismofL. seed oil., 2017, 38(15): 127-134. (in Chinese)
[28] 張利華, 張薇, 韋廣洪, 楊沛, 劉軍, 牛曉琳.-亞麻酸對糖尿病大鼠炎癥介質和氧化應激的影響. 中國應用生理學雜志, 2012, 28(1): 64-67.
ZHANG L H, ZHANG W, WEI G H, YANG P, LIU J, NIU X L. Effect of alpha-linolenic acid on inflammation and oxidative stress in diabetic rats., 2012, 28(1): 64-67. (in Chinese)
[29] WILSON K, COTTER S C, REESON A F, PELL J K. Melanism and disease resistance in insects., 2001, 4(6): 637-649.
[30] RANTALA M J, KOSKIMAKI J, TASKINEN J, TYNKKYNEN K, SUHONEN J. Immunocompetence, developmental stability and wingspot size in the damselflyL.:, 2000, 267(1460): 2453-2457.
[31] COTTER S C, KRUUK L E B, WILSON K. Costs of resistance: genetic correlations and potential trade-offs in an insect immune system., 2004, 17(2): 421-429.
[32] KRAAIJEVELD A R, LIMENTANI E C, GODFRAY H C. Basis of the trade-off between parasitoid resistance and larval competitive ability in.:, 2001, 268(1464): 259-261.
[33] BEDICK J C, TUNAZ H, NOR ALIZA A R, PUTNAM S M, ELLIS M D, STANLEY D W. Eicosanoids act in nodulation reactions to bacterial infections in newly emerged adult honey bees,, but not in older foragers.:, 2001, 130(1): 107-117.
[34] 雙金, 金曙光, 包鵬云, 楊愛軍, 嘎爾迪. 探討富含-亞麻酸的添加劑對奶牛脂肪代謝及免疫功能的影響. 黑龍江畜牧獸醫, 2004(11): 16-18.
SHUANG J, JIN S G, BAO P Y, YANG A J, Ga R D. Investigation of the effects of alpha-linolenic acid-rich additives on fat metabolism and immune function of dairy cows., 2004(11): 16-18. (in Chinese)
[35] LESSARD M , GAGNON N, GODSON D L, PETIT H V. Influence of parturition and diets enriched in n-3 or n-6 polyunsaturated fatty acids on immune response of dairy cows during the transition period., 2004, 87(7): 2197-2210.
[36] 陳士勇, 咼于明, 夏兆剛, 袁建敏.不同類型多不飽和脂肪酸對產蛋雞免疫功能及肝臟脂質過氧化的影響. 營養學報, 2003, 25(4): 383-388.
CHEN S Y, guO Y M, XIA Z G, YUAN J M. Effects of different types of polyunsaturated fatty acids on humoral immune function and hepatic lipid peroxidation of laying hens., 2003, 25(4): 383-388. (in Chinese)
Effect of dietary-linolenic acid levels on physiological function ofworker Bee larvae
YU Jing, ZHANG WeiXing, MA LanTing, xu baoHua
(College of Animal Science and Technology, Shandong Agricultural University, Taian 271018, Shandong)
】The objective of this study is to explore the effect of dietary-linolenic acid levels on the antioxidant activity and immunity ofworker bee larvae.【】Twelve hundreds 1-day-old worker bee larvae were randomly divided into 5 group, with 5 replicates in each group and 48 worker bee larvae in each replicate. The control group was fed with basic diets without-linolenic acid, and the 4 treatment groups were fed experimental diets containing different amounts of-linolenic acid (0.02%, 0.04%, 0.06%, and 0.08%). According to the indoor feedingmethod, the 1-day-old worker bee larvae were moved to a 24-well cell culture plate with 200 μL diet at suitable temperature. The larvae were moved by worm-moving needle. The culture plate was placed in a constant temperature incubator (temperature 33℃, relative humidity 55%), the diet was changed daily during the test. Feeding until the end of the 6th day or the beginning of the 7th day, when the larvae started to stand upright or defecate, the larvae were moved to the 24-well cell culture plate covered with sterilized paper in advance to prepare for pupation. From the 1st day of feeding, the death number of larvae and pupae was checked and recorded every day, and the dead individuals were removed in time until all adult emerged. The number of undead pupae and emergence new bees was recorded, and pupation rate and emergence rate were calculated. The 5-, 6- and 7-day-old worker bee larvae were used to measure antioxidant, immune, lipid metabolism indexes and related gene expression.【】 When the addition level of-linolenic acid in the diet was 0.02% and 0.04%, the pupation rate and emergence rate were significantly higher than those in other treatment groups (<0.05), while the contents of triglyceride (TG), total cholesterol (TC) and low density lipoprotein (LDL) in the hemolymph of worker bee larvae were significantly lower than those of the control group, the content of high density lipoprotein (HDL) was significantly higher than that of the control group (<0.05). Compared with the control group, the activity of superoxide dismutase (SOD) was significantly increased, while the content of malondialdehyde (MDA) was significantly decreased when the addition level of-linolenic acid was 0.04% (<0.05). When the addition level of-linolenic acid was 0.02%, 0.04% and 0.06%, the activities of lysozyme and phenoloxidase (PO) of 6-day-old worker bee larvae were significantly higher than those in the control group (<0.05). When the addition level of-linolenic acid was 0.04%, the activities of fatty acid synthase (FAS) and acetyl CoA carboxylase (ACC) of 6-day-old worker bee larvae were significantly lower than those in the control group (<0.05). When the addition level of-linolenic acid was 0.04%, the relative expression levels ofandof 5- and 7-day-old worker bee larvae were significantly higher than those in the control group, but when the addition level of-linolenic acid was 0.08%, the relative expression level ofwas significantly decreased (<0.05). 【】Different dietary supplementation levels of-linolenic acid have significant effects on antioxidant activity and immunity ofworker bee larvae, and the appropriate addition level of-linolenic acid in larval diet is 0.02%-0.04%.
; worker bee;-linolenic acid; addition level; antioxidant activity; immunity
10.3864/j.issn.0578-1752.2019.13.015
2019-03-07;
2019-03-27
國家蜂產業技術體系建設專項資金(CARS-44)、山東省農業良種工程(南種北繁)(2017LZN006)、山東省“雙一流”學科建設經費(2016-2020)
于靜,E-mail:1391031996@qq.com。
胥保華,E-mail:bhxu@sdau.edu.cn
(責任編輯 岳梅)
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
