日中性草莓不同季節(jié)果實(shí)揮發(fā)性成分差異
2019-07-29 03:24:06董靜鐘傳飛王桂霞常琳琳孫健孫瑞張宏力李睿隗永青鄭書(shū)旗張運(yùn)濤
中國(guó)農(nóng)業(yè)科學(xué) 2019年13期
董靜,鐘傳飛,王桂霞,常琳琳,孫健,孫瑞,張宏力,李睿,隗永青,鄭書(shū)旗,張運(yùn)濤
日中性草莓不同季節(jié)果實(shí)揮發(fā)性成分差異
董靜,鐘傳飛,王桂霞,常琳琳,孫健,孫瑞,張宏力,李睿,隗永青,鄭書(shū)旗,張運(yùn)濤
(北京市林業(yè)果樹(shù)科學(xué)研究院/北京市草莓工程技術(shù)研究中心/農(nóng)業(yè)部華北地區(qū)園藝作物生物學(xué)與種質(zhì)創(chuàng)新重點(diǎn)實(shí)驗(yàn)室,北京 100093)
【】比較日中性草莓秋季和冬季果實(shí)揮發(fā)性成分,明確季節(jié)變化對(duì)揮發(fā)性成分構(gòu)成及香氣值的影響,為研究草莓香氣形成和開(kāi)發(fā)利用日中性品種提供理論依據(jù)。以日中性品種‘阿爾比’‘蒙特瑞’‘波特拉’和‘圣安德瑞斯’的秋季大拱棚栽培和冬季日光溫室栽培的成熟果實(shí)為試材,采用頂空固相微萃取和氣譜-質(zhì)譜聯(lián)用方法檢測(cè)揮發(fā)性成分并進(jìn)行定性、定量分析,計(jì)算揮發(fā)性成分的數(shù)量、含量、比例和香氣值,通過(guò)主成分分析對(duì)秋、冬果實(shí)香氣成分含量差異做進(jìn)一步分析,并依據(jù)香氣值>1的成分,利用聚類熱圖對(duì)供試樣品進(jìn)行分類。共檢測(cè)到88種揮發(fā)性成分,計(jì)算了57種成分的香氣值,其中有30種成分的香氣值>1。季節(jié)變化對(duì)各類揮發(fā)性成分?jǐn)?shù)量和總含量的影響因品種而異;冬季果實(shí)中酯類成分的含量及在總揮發(fā)性成分中的比例顯著高于秋季果實(shí),而萜烯類成分的比例則顯著低于秋季果實(shí)。主成分分析表明,‘波特拉’果實(shí)的揮發(fā)性成分含量受季節(jié)變化影響最小,而‘圣安德瑞斯’受季節(jié)影響最大。所有揮發(fā)性成分中,丁酸乙酯的平均香氣值最高,對(duì)果實(shí)香氣有重要影響;與秋季果實(shí)相比,冬季果實(shí)乙酸丁酯和丁酸甲酯的香氣值顯著升高;香氣值>1的萜烯類成分在冬季果實(shí)中的香氣值之和均低于秋季果實(shí)。丁酸乙酯、丁酸丁酯、4-甲氧基-2,5-二甲基-3()-呋喃酮(DMMF)、己酸乙酯、芳樟醇等成分在不同季節(jié)果實(shí)的香氣差異中起重要作用。通過(guò)熱圖分析可以將供試樣品分為3類,其中第I類樣品包括‘阿爾比’秋季果實(shí)、冬季果實(shí)和‘蒙特瑞’秋季果實(shí),它們的總香氣值和丁酸乙酯香氣值均高于其他樣品。季節(jié)變化主要影響果實(shí)的酯類和萜烯類成分,冬季果實(shí)的酯類含量、比例顯著高于秋季果實(shí),而萜烯類的比例則顯著低于秋季果實(shí)。4個(gè)品種中,‘波特拉’果實(shí)揮發(fā)性成分含量受季節(jié)影響最小,而‘阿爾比’果實(shí)香氣最濃,香氣受季節(jié)變化的影響最小,‘圣安德瑞斯’揮發(fā)性成分含量和香氣均受影響最大。
草莓;日中性品種;果實(shí)揮發(fā)性成分構(gòu)成;香氣值;酯類;萜烯類
0 引言
【研究意義】濃郁香氣是優(yōu)質(zhì)草莓的主要品質(zhì)性狀之一,也是消費(fèi)者挑選草莓的重要依據(jù)。中國(guó)是草莓生產(chǎn)和消費(fèi)大國(guó),栽培方式以促成栽培為主,2015年的栽培面積已超過(guò)15萬(wàn)公頃、產(chǎn)量達(dá)到400萬(wàn)噸以上[1]。目前,國(guó)內(nèi)草莓促成栽培采用的是短日照(Short-day)品種,主要供應(yīng)冬春鮮果市場(chǎng);而日中性(Day-neutral)品種的花芽分化不受日照長(zhǎng)度限制,既可用于冷涼地區(qū)夏秋草莓生產(chǎn),實(shí)現(xiàn)周年供應(yīng),也可以用于冬季促成栽培,向消費(fèi)者提供更多選擇[2-5]。生產(chǎn)者充分利用日中性草莓的這種特性,正嘗試通過(guò)設(shè)施栽培條件將夏秋草莓生產(chǎn)延長(zhǎng)至冬季,以提高經(jīng)濟(jì)效益,因此,需要加深對(duì)不同季節(jié)果實(shí)品質(zhì)性狀的了解。對(duì)日中性草莓的果實(shí)揮發(fā)性成分進(jìn)行研究,分析比較秋、冬季節(jié)香氣物質(zhì)的差異,對(duì)草莓香氣研究和日中性品種的開(kāi)發(fā)利用具有重要意義。【前人研究進(jìn)展】在草莓果實(shí)中已檢測(cè)到360余種揮發(fā)性成分,包括酯類、萜烯類、醇類、醛類、酮類、呋喃類等,這些成分組成了復(fù)雜的混合物[6-8],其中酯類已鑒定出130多種,占總揮發(fā)性成分的25%—90%[9],是種類最多的一類成分。己酸乙酯、丁酸乙酯、丁酸甲酯、己酸甲酯、4-甲氧基-2,5-二甲基-3()-呋喃酮(DMMF)、芳樟醇等成分的含量較高,在草莓香氣中起主導(dǎo)作用[10-14]。通常認(rèn)為不同的香氣物質(zhì)對(duì)果實(shí)香氣的貢獻(xiàn)不同,草莓的特征香氣由許多揮發(fā)物共同形成[15],不同類型的香氣可能與關(guān)鍵香氣成分在含量和相互比例方面的變化有關(guān)[8]。基因型是影響果實(shí)揮發(fā)性成分構(gòu)成的主要因素,果實(shí)成熟度、氣候條件、栽培方式、采后處理等因素也會(huì)使水果香氣發(fā)生改變[14,16-21]。例如,鳳梨草莓(×)和森林草莓()果實(shí)酯類構(gòu)成存在種間差異,導(dǎo)致二者的香氣類型不同[22];有關(guān)葡萄的研究表明,露地栽培葡萄的單萜類化合物的種類及含量均高于溫室栽培,具有更濃郁的香氣[18, 23]。【本研究切入點(diǎn)】國(guó)內(nèi)日中性草莓的栽培面積不斷擴(kuò)大,栽培方式也從單一的夏秋避雨栽培或冬春促成栽培,開(kāi)始逐漸向兩者結(jié)合、延長(zhǎng)采收期的方向發(fā)展。在主栽品種中,‘阿爾比’的果實(shí)香氣最濃,‘蒙特瑞’‘圣安德瑞斯’次之,‘波特拉’香氣較淡;而與冬季相比,同一品種秋季采收的果實(shí)更有“草莓味”。但目前國(guó)內(nèi)未見(jiàn)有關(guān)日中性草莓香氣的報(bào)道,國(guó)外研究也較少,而且多以春季采收的露地栽培果實(shí)為試材[24-25],尚缺乏有關(guān)季節(jié)變化對(duì)果實(shí)香氣影響的系統(tǒng)報(bào)道。【擬解決的關(guān)鍵問(wèn)題】以4個(gè)日中性品種在夏秋大拱棚栽培和冬季日光溫室栽培的果實(shí)為試材,分析比較揮發(fā)性成分種類、構(gòu)成及香氣值在不同季節(jié)果實(shí)中的差異,為研究草莓香氣形成和開(kāi)發(fā)利用日中性品種提供理論依據(jù)。
1 材料與方法
1.1 供試品種
以‘阿爾比’‘蒙特瑞’‘波特拉’和‘圣安德瑞斯’4個(gè)日中性品種為研究對(duì)象,分別選取夏秋大拱棚避雨栽培和冬季日光溫室促成栽培的果實(shí)用于揮發(fā)性成分檢測(cè)。供試植株均購(gòu)自拉森峽谷農(nóng)業(yè)發(fā)展(北京)有限責(zé)任公司,夏秋栽培的植株于2016年4月19日定植在承德市農(nóng)林科學(xué)院隆化基地的大拱棚內(nèi),生長(zhǎng)季正常管理,9月9日取樣;促成栽培的植株于2016年9月10日定植在北京市林業(yè)果樹(shù)科學(xué)研究院日光溫室內(nèi),2017年2月21日取樣。每個(gè)品種隨機(jī)選取正常成熟果實(shí)0.5 kg,3次重復(fù),采摘后立即放入塑料方盒中,加冰袋運(yùn)回實(shí)驗(yàn)室,-80℃條件下保存。
1.2 揮發(fā)性成分檢測(cè)
委托農(nóng)業(yè)部蔬菜種子質(zhì)量監(jiān)督檢驗(yàn)測(cè)試中心對(duì)供試樣品的揮發(fā)性成分進(jìn)行定性、定量測(cè)定。
采用頂空固相微萃取法(SPME)進(jìn)行樣品預(yù)處理。取樣前先將固相微萃取頭在氣相色譜進(jìn)樣口老化2 h,老化溫度250℃。采用四分法取樣打漿,將8 g勻漿迅速轉(zhuǎn)入15 mL樣品瓶中,加蓋封口。將老化好的萃取頭插入樣品瓶頂空部分,35℃水浴條件下萃取25 min,然后將萃取頭抽出,插入氣譜-質(zhì)譜聯(lián)用儀(GC-MS),于250℃解吸1 min,進(jìn)行GC-MS檢測(cè)分析。
島津GCMS-GC2010(Shimadzu Co.,Kyoto,Japan)氣譜-質(zhì)譜聯(lián)用儀測(cè)定果實(shí)揮發(fā)性成分。控制條件為:毛細(xì)管柱(DB-5MS):30 m×0.25 mm;膜厚度0.25 μm;載氣為氦氣;流速1 mL·min-1;程序升溫:40℃保持2 min,以8℃·min-1升溫到220℃,保持6 min。進(jìn)樣口溫度250℃;EI離子源電子能量70 eV,質(zhì)量范圍30—550 aum。
1.3 數(shù)據(jù)分析
未知化合物質(zhì)譜圖經(jīng)計(jì)算機(jī)檢索,同時(shí)于NIST library和Wiley library兩個(gè)質(zhì)譜庫(kù)相匹配,并結(jié)合人工圖譜解析及資料分析對(duì)各材料的總離子流圖進(jìn)行處理,采用峰面積歸一化法計(jì)算各成分在總揮發(fā)物中所占比例。混標(biāo)法制備標(biāo)準(zhǔn)曲線進(jìn)行定量分析,有標(biāo)樣的直接計(jì)算含量;沒(méi)有標(biāo)樣的根據(jù)化學(xué)結(jié)構(gòu)相似、碳原子數(shù)相近的原則進(jìn)行近似定量。
表1中序號(hào)為1、2、4、5、7、8、9、10、12、17、18、19、20、24、26、34、39、41、42、43、44、46、47、48、49、57、60、67、68、69、70、71、74、76、77、79、80、81、82、88的40種成分購(gòu)有標(biāo)樣,所有標(biāo)樣均購(gòu)自Sigma-Aldrich(St. Louis,MO,USA)。
分別利用SAS 8.01軟件對(duì)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)分析、Canoco for Windows 4.5軟件進(jìn)行主成分分析、R語(yǔ)言制作聚類熱圖。
2 結(jié)果
2.1 檢測(cè)到的果實(shí)揮發(fā)性成分
在供試的所有樣品中,共檢測(cè)到88種揮發(fā)性成分(表1),包括48種酯、10種萜烯、6種酮、9種醇、3種內(nèi)酯、1種呋喃、7種醛、4種酸,其中酯類成分?jǐn)?shù)量最多,占54.5%,是各種果味的主要來(lái)源,其次是萜烯類和醇類,分別占11.4%和10.2%,萜烯類物質(zhì)多數(shù)具有花香和甜味,醇類則提供了青味和草本氣息。秋季草莓果實(shí)中的揮發(fā)性成分?jǐn)?shù)量為80種,冬季果實(shí)中數(shù)量略低,為73種;不同季節(jié)果實(shí)中種類差別最大的揮發(fā)性成分為酯類,有13種含量較低的酯只在一個(gè)季節(jié)的果實(shí)中檢測(cè)到。乙酸己酯、乙酸-2-己烯酯、乙酸苯甲酯、丁酸甲酯、丁酸丁酯、丁酸-2-己烯酯、己酸甲酯、己酸乙酯、芳樟醇、D-檸檬烯、橙花叔醇、己醇、反式-2-己烯醇、辛醇、DMMF、反式-2-己烯醛、苯甲醛等17種成分在所有樣品中都能檢測(cè)到,是共有成分。其中,丁酸甲酯在所有揮發(fā)性成分中平均含量最高,而且在冬季果實(shí)中的含量均顯著高于秋季,約是秋季果實(shí)的2—3倍;丁酸丁酯在冬季果實(shí)中的平均含量是秋季的3.5倍;秋季果實(shí)中芳樟醇、橙花叔醇、D-檸檬烯、反式-2-己烯醇、辛醇、DMMF的平均含量明顯高于冬季果實(shí),而己醇、苯甲醛的含量均顯著高于冬季果實(shí)。
2.2 果實(shí)揮發(fā)性成分構(gòu)成分析
果實(shí)揮發(fā)性成分構(gòu)成主要包括各類揮發(fā)性成分的數(shù)量、含量及其在總揮發(fā)性成分中所占比例等[22],本研究中,供試日中性草莓的果實(shí)揮發(fā)性成分構(gòu)成在不同季節(jié)、不同品種間存在顯著差異(表2)。
‘阿爾比’‘蒙特瑞’在秋季時(shí)果實(shí)揮發(fā)性成分?jǐn)?shù)量顯著高于冬季,而‘波特拉’和‘圣安德瑞斯’在冬季時(shí)果實(shí)揮發(fā)性成分?jǐn)?shù)量會(huì)增多;在各類成分中,酯類數(shù)量變化最大,這也是引起總揮發(fā)物數(shù)量發(fā)生變化的主要原因。‘阿爾比’秋季果實(shí)的酯類、醇類數(shù)量顯著高于冬季果實(shí),內(nèi)酯數(shù)量顯著低于冬季果實(shí),其他變化不顯著;‘蒙特瑞’秋季果實(shí)中,酯類、內(nèi)酯類和酸類成分?jǐn)?shù)量顯著高于冬季果實(shí);‘波特拉’秋季果實(shí)的酯類、酸類數(shù)量顯著低于冬季果實(shí),萜烯類、內(nèi)酯類、醛類數(shù)量則顯著高于冬季果實(shí);‘圣安德瑞斯’秋季果實(shí)中只有酯類數(shù)量顯著低于冬季果實(shí),而萜烯類、醛類數(shù)量顯著高于冬季果實(shí)。

表1 日中性草莓中檢測(cè)到的揮發(fā)性成分及其氣味閾值和氣味描述

續(xù)表1 Continued table 1

續(xù)表1 Continued table 1

續(xù)表1 Continued table 1
ND表示“未檢測(cè)到”
ND means ‘not detected’
‘阿爾比’和‘波特拉’冬季果實(shí)的揮發(fā)性成分總含量顯著高于秋季果實(shí),‘圣安德瑞斯’在冬季時(shí)揮發(fā)性成分含量顯著低于秋季,而‘蒙特瑞’差異不顯著。在所有供試品種中,秋季果實(shí)的酯類含量均顯著低于冬季果實(shí),而萜烯類含量則高于冬季果實(shí),其中‘阿爾比’‘蒙特瑞’和‘圣安德瑞斯’差異顯著。另外,‘阿爾比’秋季果實(shí)的酮類、內(nèi)酯類、醛類、酸類含量分別顯著低于冬季果實(shí),醇類含量顯著高于冬季果實(shí);‘蒙特瑞’秋季果實(shí)的呋喃類、醛類、酸類含量均顯著高于冬季果實(shí);‘波特拉’秋季果實(shí)的醇類含量顯著高于冬季,酸類則顯著低于冬季;‘圣安德瑞斯’秋季果實(shí)的酮類、醇類、內(nèi)酯類、呋喃類、醛類含量顯著高于冬季,而酸類含量則顯著低于冬季果實(shí)。
供試4個(gè)品種的酯類成分在總揮發(fā)性成分中的比例都是秋季顯著低于冬季;萜烯類、醇類、呋喃類與酯類相反,秋季果實(shí)中的比例顯著高于冬季;‘波特拉’和‘圣安德瑞斯’秋季果實(shí)的酮類比例顯著高于冬季;除了‘圣安德瑞斯’秋季果實(shí)中內(nèi)酯類的比例顯著高于冬季,其他品種無(wú)顯著變化;在醛類所占比例方面,‘蒙特瑞’‘波特拉’‘圣安德瑞斯’均為秋季果實(shí)顯著高于冬季果實(shí);只有‘蒙特瑞’秋季果實(shí)的酸類比例顯著高于冬季,其他3個(gè)品種冬季果實(shí)酸類比例則顯著增加。
2.3 揮發(fā)性成分的主成分分析
為進(jìn)一步比較供試8個(gè)樣品的果實(shí)揮發(fā)性成分差異,對(duì)檢測(cè)到的88種成分的含量進(jìn)行主成分分析,PC1和PC2的貢獻(xiàn)率分別為39.2%和31.8%(圖1)。在4個(gè)品種中,只有‘波特拉’的秋、冬季樣品分布于同一象限,2個(gè)樣品間距離最小,說(shuō)明‘波特拉’果實(shí)的揮發(fā)性成分及其含量受季節(jié)變化影響最小,而‘圣安德瑞斯’的樣品間距離最遠(yuǎn),果實(shí)揮發(fā)性成分含量受季節(jié)影響最大。不同品種的冬季果實(shí)均趨于PC1的負(fù)半軸分布,除‘波特拉’外,其他3個(gè)品種的秋季果實(shí)均趨于PC1的正半軸。88種揮發(fā)性成分在載荷圖中主要分布于PC2的正半軸,其中貢獻(xiàn)較大的包括丁酸乙酯、丁酸丁酯、芳樟醇、橙花叔醇、-金合歡烯、苯甲醛、丁酸-2-己烯酯、2-庚酮、2-甲基丙酸-1-甲基丁酯、2-庚醇、-癸內(nèi)酯、-十二內(nèi)酯、DMMF等,以上成分在供試品種不同季節(jié)果實(shí)的香氣物質(zhì)含量差異中起重要作用。
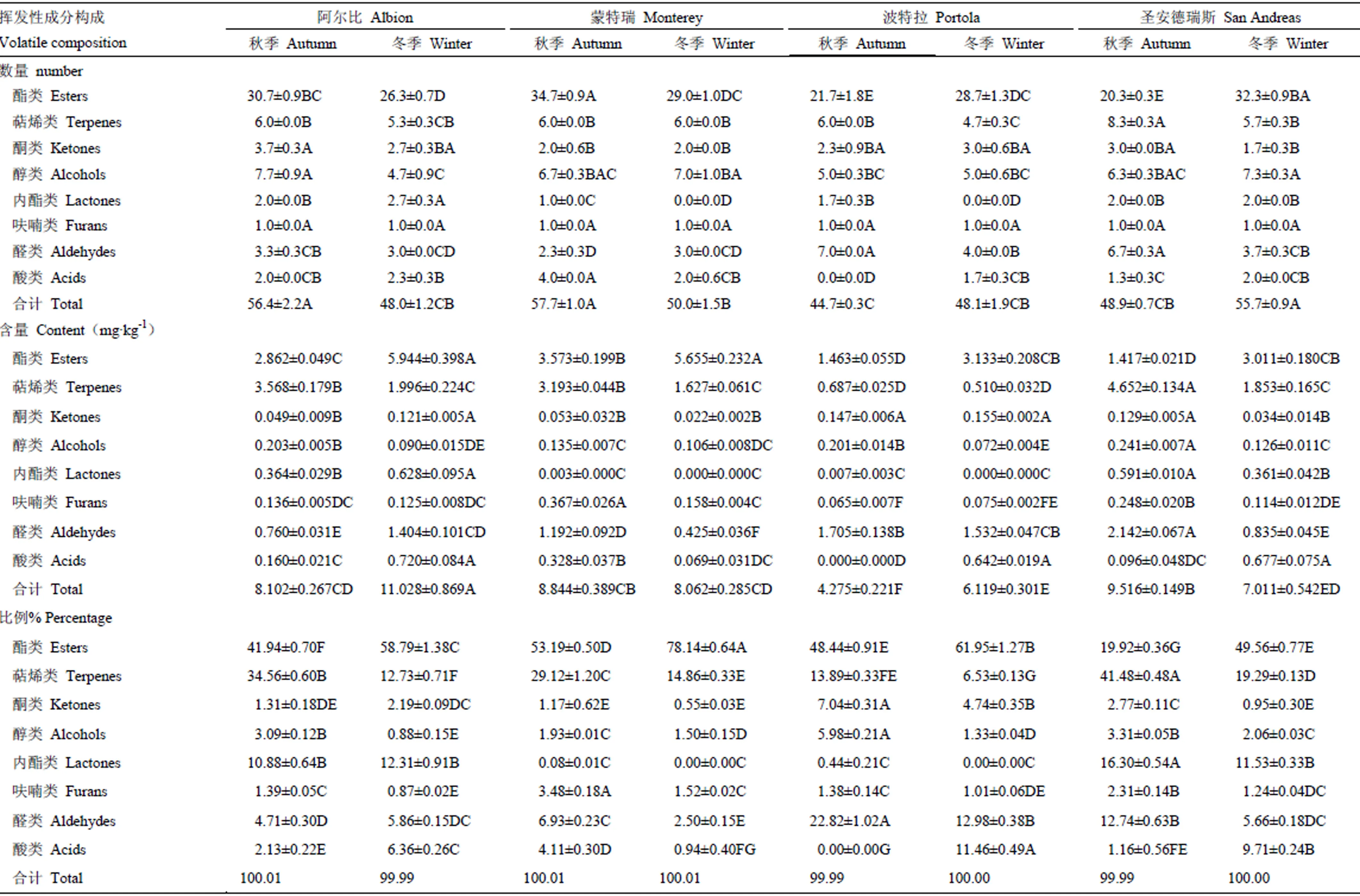
表2 不同季節(jié)日中性草莓果實(shí)揮發(fā)性成分構(gòu)成
每行的不同字母表示在<0.01水平差異極顯著。下同
Means with the different letter are significantly different at<0.01 in each row. The same as below
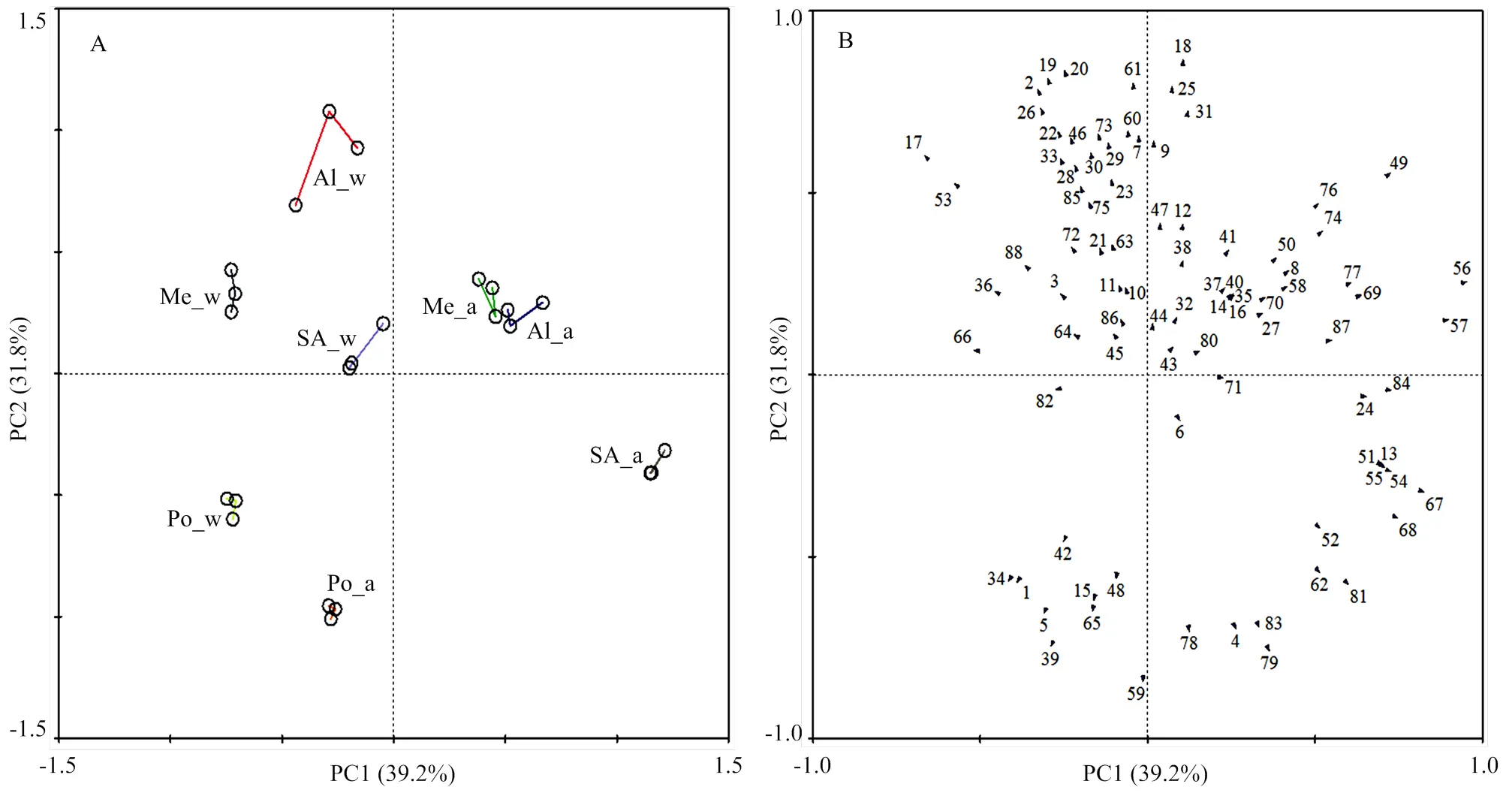
A:主成分得分圖;B:主成分載荷圖 A: PCA score plots; B: PCA loading plots
Al_a:阿爾比_秋;Al_w:阿爾比_冬;Me_a:蒙特瑞_秋;Me_w:蒙特瑞_冬;Po_a:波特拉_秋;Po_w:波特拉_冬;SA_a:圣安德瑞斯_秋;SA_w:圣安德瑞斯_冬。圖中數(shù)字對(duì)應(yīng)表1中的88種揮發(fā)性成分。下同
Al_a: Albion_autumn; Al_w: Albion_winter; Me_a: Menterey_autumn; Me_w: Menterey_winter; Po_a: Portola_autumn; Po_w: Portola_winter; SA_a: San Andreas_autumn; SA_w: San Andreas_winter. The numbers in this figure correspond to the 88 volatiles listed in table 1. The same as below
圖1 不同季節(jié)日中性草莓果實(shí)揮發(fā)性成分檢測(cè)含量的主成分分析
Fig. 1 Principal component analysis of detected content of volatiles in day-neutral strawberries harvested in autumn and winter
2.4 主要揮發(fā)性成分的香氣值分析
揮發(fā)性成分的香氣值是含量與香氣閾值之比,能反映該成分對(duì)香氣的貢獻(xiàn),而含量≥香氣閾值的成分才能被感覺(jué)器官感知。與低香氣值成分相比,很顯然,香氣值≥1的成分對(duì)香氣形成的貢獻(xiàn)更大[14,42]。本研究中有57種揮發(fā)性成分檢索到香氣閾值并計(jì)算了香氣值(表1、表3),其中有30種成分在至少1個(gè)樣品中的香氣值>1,下面就以這些香氣值>1的揮發(fā)性成分為具體分析對(duì)象。
乙酸己酯、丁酸甲酯、己酸甲酯、己酸乙酯、芳樟醇、橙花叔醇、DMMF、反式-2-己烯醛、壬醛9種成分在所有樣品中的香氣值均大于1,是供試品種果實(shí)香氣的主要組成成分。丁酸乙酯的平均香氣值最高,為261.50,其次為己酸乙酯214.17,芳樟醇166.08位列第三;乙酸丁酯、丁酸甲酯在4個(gè)供試品種冬季果實(shí)中的香氣值均顯著高于秋季果實(shí),而芳樟醇、橙花叔醇、反式-2-己烯醇、順式-2-庚烯醛則呈相反趨勢(shì),香氣值在其中3個(gè)品種的冬季果實(shí)中顯著降低。
這些成分的分布及其在不同季節(jié)間的變化趨勢(shì)因品種而異。丁酸乙酯、芳樟醇、己酸乙酯、反式-2-己烯醛、-癸內(nèi)酯、丁酸甲酯、乙酸己酯、壬醛等成分對(duì)‘阿爾比’果實(shí)香氣貢獻(xiàn)較大;‘阿爾比’秋季果實(shí)中,含有17種香氣值>1的成分,它們的香氣值之和為1 094.84,而冬季果實(shí)中則含有19種,香氣值之和為1 610.72,大大高于秋季;有14種成分在兩個(gè)季節(jié)的果實(shí)中香氣值均>1,其中芳樟醇和橙花叔醇秋季的香氣值顯著高于冬季,乙酸己酯、丁酸甲酯、丁酸乙酯、己酸乙酯、-癸內(nèi)酯等10種成分則顯著低于冬季;與秋季果實(shí)相比,冬季果實(shí)中酯類、醛類、內(nèi)酯類、酮類成分的香氣值分別升高,萜烯類、呋喃類的香氣值降低,其中丁酸乙酯、己酸乙酯的香氣值增加最多。
‘蒙特瑞’的秋季和冬季果實(shí)中各含有18種香氣值>1的成分,它們的香氣值之和分別為1 094.07和759.86,冬季略低;其中16種成分相同,丁酸乙酯、芳樟醇、己酸乙酯、乙酸己酯、反式-2-己烯醛、DMMF、壬醛、丁酸甲酯、橙花叔醇等對(duì)‘蒙特瑞’果實(shí)香氣形成起重要作用。乙酸己酯、乙酸辛酯、丁酸乙酯、己酸乙酯、橙花叔醇、DMMF、反式-2-己烯醛等成分在秋季的香氣值顯著高于冬季;酯類、萜烯類、醛類和呋喃類在冬季果實(shí)中的香氣值均低于秋季果實(shí)。

表3 不同季節(jié)日中性草莓果實(shí)的揮發(fā)性成分香氣值

續(xù)表3 Continued table 3

續(xù)表3 Continued table 3
和其他3個(gè)品種相比,‘波特拉’果實(shí)中香氣值>1的成分較少,秋季和冬季果實(shí)中分別含有17種和14種成分,總香氣值也較低,僅為461.17和610.13;其中冬季果實(shí)酯類香氣值之和是秋季的3.8倍,呋喃類香氣值比秋季略有增加,而萜烯類和醛類的香氣值則分別低于秋季果實(shí);癸醛是秋季果實(shí)中香氣值最高的成分,而己酸乙酯在冬季果實(shí)中的香氣值比秋季果實(shí)高316.67,占總香氣值的50%以上,是引起冬季果實(shí)香氣值增加的主要因素;總的來(lái)說(shuō),己酸乙酯、芳樟醇、反式-2-己烯醛、壬醛、己醛、3-甲基丁酸甲酯、乙酸己酯、丁酸甲酯等對(duì)‘波特拉’香氣形成作用較大。
‘圣安德瑞斯’果實(shí)中香氣值>1的成分較多,在秋季和冬季果實(shí)中數(shù)量分別為18種和21種,己酸乙酯、芳樟醇、癸醛、-癸內(nèi)酯、反式-2-己烯醛、壬醛、乙酸己酯等是‘圣安德瑞斯’的重要香氣成分;秋季‘圣安德瑞斯’香氣值>1成分的總香氣值為1 027.48,高于冬季的779.88,但秋季果實(shí)中未檢測(cè)到丁酸乙酯,而冬季果實(shí)含有丁酸乙酯,并且香氣值較高;酯類的數(shù)量和香氣值在冬季均高于秋季,萜烯類、醛類、內(nèi)酯類、呋喃類成分的香氣值則低于秋季果實(shí),其中己酸乙酯、芳樟醇、癸醛的香氣值顯著降低,是造成‘圣安德瑞斯’冬季總香氣值下降的主要原因。
2.5 30種香氣值>1成分的聚類熱圖分析
通過(guò)聚類熱圖分析,可以將供試樣品分為3大類(圖2)。第I類包括‘阿爾比’秋季果實(shí)、‘蒙特瑞’秋季果實(shí)和‘阿爾比’冬季果實(shí),這3個(gè)樣品的總香氣值和丁酸乙酯的香氣值均高于其他樣品,另外,芳樟醇、己酸乙酯、丁酸辛酯、丁酸丁酯、-十二內(nèi)酯、DMMF的香氣值也較高,戊醛、己酸甲酯、己醛、2-甲基丁酸甲酯的香氣值較低,癸醛香氣值為0;第II類包括‘圣安德瑞斯’冬季果實(shí)、‘蒙特瑞’冬季果實(shí)和‘波特拉’秋季果實(shí),芳樟醇、己酸乙酯、壬醛、乙酸-2-甲基丁酯、2-甲基丁酸甲酯香氣值較高,反式-2-己烯醛、DMMF、橙花叔醇香氣值較低;第III類只有‘波特拉’冬季果實(shí)和‘圣安德瑞斯’秋季果實(shí),和前兩類相比,這一類的己酸乙酯香氣值最高,壬醛、己酸甲酯、D-檸檬烯、己醛香氣值較高,乙酸己酯、乙酸葉醇酯、乙酸丁酯、丁酸丁酯、2-戊酮香氣值較低,丁酸乙酯香氣值為0。另外,從圖2也可以看出,‘阿爾比’秋、冬果實(shí)同屬第I類,香氣差異最小;‘圣安德瑞斯’的大多數(shù)成分在兩個(gè)季節(jié)果實(shí)中的香氣值差異大,因而香氣變化最大。

圖2 不同季節(jié)日中性草莓果實(shí)中30種揮發(fā)性成分香氣值的熱圖分析
3 討論
研究表明,光照、溫度是影響揮發(fā)性成分合成的主要環(huán)境因素,光強(qiáng)和光質(zhì)都能調(diào)控?fù)]發(fā)物的合成,溫度變化也能導(dǎo)致植物揮發(fā)性成分的差異[43-49]。提高光合有效輻射可以促進(jìn)多種落葉樹(shù)木合成和釋放甲醇[49];當(dāng)光強(qiáng)從65 μmol·m-2·s-1增強(qiáng)到295 μmol·m-2·s-1,冬小麥植株中的芳樟醇、-丁子香烯、乙酸苯甲酯、()-羅勒烯、()-3-己烯醛、()-2-己烯醛、()-3-己烯-1-醇、()-2-己烯-1-醇、乙酸己酯和乙酸-()-3-己烯-1-酯等成分的含量也隨之升高[50];較高的光量子通量密度(PFD,photon flux density)能增加山香屬植物中香檜烯、-蒎烯、-水芹烯、雙環(huán)大牻牛兒烯等萜烯類成分含量,而日光燈光譜、紅光、10﹕4紅藍(lán)光等不同光質(zhì)也會(huì)影響萜烯類合成,光質(zhì)不同造成的影響也不同[45];矮牽牛中2-苯乙醇的含量水平則在紅光和遠(yuǎn)紅光下升高[46]。溫度對(duì)不同植物揮發(fā)性成分的影響存在差異,隨著栽培溫度在22℃/15℃、25℃/18℃、28℃/20℃范圍內(nèi)升高,蓼()葉片中揮發(fā)性醛的相對(duì)含量降低,而萜烯、醇、烷烴的相對(duì)含量增高[51];與晝/夜溫度22℃/16℃相比,28℃/22℃使矮牽牛花的苯甲醛、苯甲醇、苯乙醇、苯甲酸苯甲酯、苯甲酸乙酯、苯甲酸甲酯、丁子香酚、異丁子香酚、苯乙醛等揮發(fā)性成分的含量降低,這可能與高溫下調(diào)了相關(guān)基因表達(dá)有關(guān)[43]。揮發(fā)性成分合成和色素合成間存在一定聯(lián)系,苯丙氨酸在L-苯丙氨酸脫氨酶(PAL)作用下的產(chǎn)物可以進(jìn)入苯丙烷類代謝途徑和花青素合成途徑,分別生成苯丙烷類揮發(fā)性成分和花青素類色素[43,52-53],而調(diào)控植物色素合成的主要外部因素也是光、溫條件[52,54-55]。因此,屬于R2R3-MYB家族的一些轉(zhuǎn)錄因子也能參與揮發(fā)性成分代謝的調(diào)控,例如擬南芥的、甜櫻桃的、蘋(píng)果的等,在矮牽牛和玫瑰中表達(dá),能激活苯丙烷類代謝途徑中的多個(gè)基因,促進(jìn)花青素和香氣物質(zhì)的合成[55-59]。高溫環(huán)境會(huì)使矮牽牛中苯丙烷類香氣成分的合成受抑制,這可能與苯丙烷類代謝途徑和莽草酸途徑中相關(guān)結(jié)構(gòu)基因的表達(dá)下調(diào)和負(fù)調(diào)控因子的表達(dá)量增加有關(guān),但在矮牽牛中過(guò)量表達(dá),即使在長(zhǎng)期高溫環(huán)境中,轉(zhuǎn)基因株系的揮發(fā)物水平也會(huì)保持不變甚至更高[43]。研究發(fā)現(xiàn),在月季中,miR156-調(diào)控模塊能調(diào)控花青素及大牻牛兒烯D和一些萜烯類揮發(fā)性成分的合成,隨著miR156下調(diào)表達(dá),形成MYB-bHLH- WD40蛋白復(fù)合體,激活了花青素合成酶基因和大牻牛兒烯D合成酶基因,而的表達(dá)下調(diào)會(huì)降低包括橙花叔醇合酶基因在內(nèi)的一些萜烯類合成酶基因的表達(dá),從而導(dǎo)致某些萜烯類成分的減少甚至缺失[60]。
季節(jié)變化、氣候差異以及不同栽培模式所導(dǎo)致光照、溫度等環(huán)境條件的變化,會(huì)直接影響果實(shí)香氣物質(zhì)合成。在中國(guó)釀酒葡萄產(chǎn)區(qū)中,河北昌黎夏季炎熱、半濕潤(rùn),而甘肅高臺(tái)夏季涼爽、干燥,兩地生產(chǎn)的‘赤霞珠’葡萄的揮發(fā)性成分存在差異[61],昌黎所產(chǎn)‘赤霞珠’含有更豐富的苯甲醇、苯乙醇、1-己醇、1-辛烯-3-醇等醇類物質(zhì),而己醛、庚醛、2-甲氧基-3-異丁基吡嗪和反式--大馬酮在高臺(tái)所產(chǎn)‘赤霞珠’中的含量水平更高。溫室栽培能提高鮮食葡萄果實(shí)中的綠葉香氣,但露地栽培條件下,對(duì)香氣貢獻(xiàn)較大、具有花香和甜香氣味的單萜化合物含量更高,這種差異可以在一定程度上解釋溫室葡萄和露地葡萄香氣的不同[18,62]。同樣,具有柑橘、花香等氣味的癸醛、橙花醛、香葉醛、-紫羅酮、金合歡基丙酮、香葉基丙酮等成分在露地番茄中的水平顯著高于大拱棚番茄[63]。研究發(fā)現(xiàn),光合有效輻射是影響露地草莓揮發(fā)性成分構(gòu)成的主要因素[24],光照強(qiáng)度能正向調(diào)控C13-降異戊二烯和單萜類物質(zhì)的合成,因而設(shè)施條件下的光照減弱影響了萜烯合成相關(guān)基因的表達(dá),降低了果實(shí)中單萜化合物的含量,特別是芳樟醇的含量[64-66],而光照減弱能促進(jìn)‘Delicious’蘋(píng)果酯類物質(zhì)的合成[67]。本研究中,4個(gè)供試品種秋季果實(shí)的酯類含量、比例均顯著低于冬季果實(shí),萜烯類含量高于冬季果實(shí),而萜烯類、醇類、呋喃類比例分別顯著高于冬季果實(shí),而且乙酸丁酯、丁酸甲酯等酯類成分和芳樟醇、橙花叔醇等萜烯類成分的香氣值變化也遵循這一規(guī)律,與前人研究結(jié)果基本相符,說(shuō)明秋季大拱棚內(nèi)較強(qiáng)的光照能促進(jìn)草莓萜烯類成分的合成,冬季日光溫室的弱光則有利于酯類合成。另外,有研究表明,低于15℃的氣溫有助于草莓香氣物質(zhì)的生成,Sanz等[68]對(duì)花后24—31 d的‘Camarosa’果實(shí)進(jìn)行離體培養(yǎng),發(fā)現(xiàn)25℃/10℃的晝/夜溫度最有利于香氣成分積累,但目前尚缺少溫度影響草莓揮發(fā)性成分構(gòu)成的報(bào)道。八倍體栽培草莓基因組測(cè)序已完成[69],有助于今后深入挖掘參與草莓香氣代謝及調(diào)控的基因,也有助于進(jìn)一步研究環(huán)境因素的變化對(duì)香氣形成的影響,這對(duì)人為調(diào)控草莓果實(shí)品質(zhì)具有重要價(jià)值。
根據(jù)香氣值,可以將果實(shí)的所有揮發(fā)性成分分為兩類,其中香氣值<1的成分的作用尚未明確,而香氣值≥1的成分可以通過(guò)感官感知,是與果實(shí)香氣表型相關(guān)的優(yōu)勢(shì)成分[8,70]。本研究中,有30種成分在至少1個(gè)樣品中香氣值>1,其中乙酸己酯、丁酸甲酯、丁酸乙酯、己酸甲酯、己酸乙酯、芳樟醇、DMMF、-癸內(nèi)酯、反式-2-己烯醛等成分在草莓中早有報(bào)道,與果實(shí)香氣密切相關(guān)[10,12,14,16,28,71-74]。果實(shí)的香氣特點(diǎn)主要由高香氣值成分決定[60],通過(guò)比較各樣品香氣值>1的成分以及進(jìn)行聚類熱圖分析,可以發(fā)現(xiàn),本研究中丁酸乙酯、己酸乙酯、丁酸丁酯、DMMF、芳樟醇等成分對(duì)不同季節(jié)果實(shí)香氣差異貢獻(xiàn)較大。而將樣品分類后,3類樣品的高香氣值成分各不相同:第I類的總香氣值最高,3個(gè)樣品丁酸乙酯的香氣值均遠(yuǎn)遠(yuǎn)高于其他樣品,芳樟醇和己酸乙酯的香氣值也比較高;第II類樣品的香氣值較高的成分為芳樟醇和己酸乙酯;第III類的特點(diǎn)是己酸乙酯的香氣值高于其他樣品,而丁酸乙酯香氣值為0。草莓果實(shí)中與“草莓味”正相關(guān)的揮發(fā)性成分是3-甲基丁酸甲酯(2=0.90)和丁酸乙酯(2=0.96)[14],如果將本研究供試樣品的3-甲基丁酸甲酯和丁酸乙酯的香氣值相加進(jìn)行比較,結(jié)果會(huì)發(fā)現(xiàn)該值在第I類中最高,第II類居中,第III類最低,與熱圖分類結(jié)果完全相同,也與樣品的田間表現(xiàn)基本一致。總的來(lái)說(shuō),4個(gè)供試品種中,‘阿爾比’表現(xiàn)為果實(shí)香氣較濃,而且香氣的季節(jié)間差異最小,而‘圣安德瑞斯’果實(shí)香氣受季節(jié)影響最大。
4 結(jié)論
季節(jié)變化能顯著影響日中性草莓果實(shí)的酯類和萜烯類成分構(gòu)成,與秋季相比,4個(gè)供試品種冬季果實(shí)的酯類含量、比例升高,其中丁酸甲酯、丁酸丁酯的平均含量提高1—2倍,而萜烯類含量、比例降低,芳樟醇、橙花叔醇、D-檸檬烯等成分平均含量明顯降低。香氣值>1的成分在不同季節(jié)間的分布趨勢(shì)因品種而異,丁酸乙酯、己酸乙酯、芳樟醇是平均香氣值最高的3種成分,丁酸乙酯、己酸乙酯、丁酸丁酯、DMMF、芳樟醇等成分對(duì)不同季節(jié)果實(shí)香氣差異貢獻(xiàn)較大。供試品種中,‘波特拉’果實(shí)揮發(fā)性成分含量受季節(jié)影響最小;‘阿爾比’在秋季和冬季均表現(xiàn)為總香氣值高,丁酸乙酯香氣值也高,果實(shí)香氣受季節(jié)影響最小;而‘圣安德瑞斯’果實(shí)揮發(fā)性成分含量和香氣均受季節(jié)影響最大。季節(jié)轉(zhuǎn)換引起的光照和溫度變化可能是造成日中性草莓果實(shí)香氣季節(jié)性差異的主要原因。
[1] ZHANG Y T, WANG G X, DONG J, ZHONG C F, CHANG L L, ZHANG H L. The current progress in strawberry breeding in China., 2017, 1156: 7-11.
[2] DALE A, HANCOCK J F, LUBY J J. Breeding dayneutral strawberries for northern North America., 2002, 567: 133-136.
[3] 沈嵐, 張國(guó)芳, 呂鵬飛, 趙天榮, 趙已棟, 黃堅(jiān). 美系日中性草莓寧波引種試驗(yàn). 黑龍江農(nóng)業(yè)科學(xué), 2016, 3: 74-76.
SHEN L, ZHANG G F, LYU P F, ZHAO T R, ZHAO Y D, HUANG J. Experiment of introduction American mid-day strawberry varieties in Ningbo., 2016, 3: 74-76. (in Chinese)
[4] 董靜, 張運(yùn)濤, 王桂霞, 鐘傳飛, 常琳琳. 日光溫室栽培不同四季草莓的產(chǎn)量品質(zhì)比較//張運(yùn)濤, 雷家軍, 王桂霞. 草莓研究進(jìn)展(V). 北京: 中國(guó)農(nóng)業(yè)出版社, 2017: 365-369.
DONG J, ZHANG Y T, WANG G X, ZHONG C F, CHANG L L. Comparison on yield and fruit quality of day-neutral and everbearing strawberries planted in greenhouse//ZHANG Y T, LEI J J, WANG G X.. Beijing: Chinese Agriculture Press, 2017: 365-369. (in Chinese)
[5] 汪國(guó)鮮, 孫建麗, 蔣海玉, 楊春梅, 吳麗芳, 單芹麗, 余蓉培, 阮繼偉. 日中性草莓品種主要經(jīng)濟(jì)性狀評(píng)價(jià). 西南農(nóng)業(yè)學(xué)報(bào), 2018, 31(10): 2179-2184.
WANG G X, SUN J L, JIANG H Y, YANG C M, WU L F, SHAN Q L, YU R P, RUAN J W. Determination and analysis of major economic traits of day-neutral strawberry cultivars., 2018, 31(10): 2179-2184. (in Chinese)
[6] PYYSALO T, HONKANEN E, HIRVI T. Volatiles of wild strawberries,L, compared to those of cultivated berries,cv Senga sengana., 1979, 27: 19-22.
[7] ULRICH D, HOBERG E, RAPP A, KECKE S. Analysis of strawberry flavor-discrimination of aroma types by quantification of volatile compounds., 1997, 205: 218-223.
[8] ZABETAKIS I, HOLDEN M A. Strawberry flavor: analysis and biosynthesis., 1997, 74: 421-434.
[9] DOUILLARD C, GUICHARD E. The aroma of strawberry (): Characterization of some cultivars and influence of freezing., 1990, 50: 517-531.
[10] LARSEN M, POLL L. Odour thresholds of some important aroma compounds in strawberries., 1992, 195: 120-123.
[11] SCHIEBERLE P, HOFMANN T. Evaluation of the character impact odorants in fresh strawberry juice by quantitative measurements after sensory studies on model mixtures., 1997, 45: 227-232.
[12] PéREZ A G, RIOS J J, SANZ C, OLíAS J M. Aroma components and free amino acids in strawberry variety Chandler during ripening., 1992, 40: 2232-2235.
[13] PéREZ A G, SANZ C, OLíAS J M. Aroma quality evaluation of strawberry cultivars in Southern Spain., 1997, 439: 337-340.
[14] DU X, PLOTTO A, BALDWIN E, ROUSEFF R. Evaluation of volatiles from two subtropical strawberry cultivars using GC- Olfactometry, GC-MS Odor activity values, and sensory analysis., 2011, 59(23): 12569-12577.
[15] 張運(yùn)濤, 董靜, 王桂霞. 草莓香味的形成和香味育種. 中國(guó)農(nóng)業(yè)科學(xué), 2004, 37(7): 1039-1044.
ZHANG Y T, DONG J, WANG G X. Formation of aroma volatiles in strawberry fruit and aroma breeding., 2004, 37(7): 1039-1044. (in Chinese)
[16] FORNEY C F, KALT W, JORDAN M A. The composition of strawberry aroma is influenced by cultivars, maturity, and storage., 2000, 35: 1022-1026.
[17] GóMEZ-MíGUEZ M J, GóMEZ-MíGUEZ M, VICARIO I M, HEREDIA F J. Assessment of colour and aroma in white wines vinifications: Effects of grape maturity and soil type., 2007, 79(3): 758-764.
[18] 孫磊, 錢(qián)旭, 張國(guó)軍, 閆愛(ài)玲, 王曉玥, 王慧玲, 任建成, 徐海英. ‘香妃’和‘早玫瑰香’葡萄溫室與露地栽培單萜積累差異分析. 園藝學(xué)報(bào), 2018,45(8): 1467-1478.
SUN L, QIAN X, ZHANG G J, YAN A L, WANG X Y, WANG H L, REN J C, XU H Y. Differential monoterpenes accumulation of ‘Xiangfei’ and ‘Zao Meiguixiang’ grapes between greenhouse and open-field cultivation., 2018, 45(8): 1467-1478. (in Chinese)
[19] LU H Y, WANG K D, WANG L, LI D, YAN J W, BAN Z J, LUO Z S, LI L, YANG D M. Effect of superatmospheric oxygen exposure on strawberry (Duch.) volatiles, sensory and chemical attributes., 2018, 142: 60-71.
[20] FU X M, CHENG S H, ZHANG Y Q, DU B, FENG C, ZHOU Y, MEI X, JIANG Y M, DUAN X W, YANG Z Y. Differential responses of four biosynthetic pathways of aroma compounds in postharvest strawberry (Duch.) under interaction of light and temperature., 2017, 221: 356-364.
[21] IHSAN K, INCILAY G, TUGCA B, KUBRA S, IHRAHIM SANI O, BANU B, BELGIN C, FERDA S. Effect of fruit maturity level on quality, sensory properties and volatile composition of two common apricot (L.) varieties., 2018, 55(7): 2671-2678.
[22] DONG J, ZHANG Y T, TANG X W, JIN W M, HAN Z H. Differences in volatile ester composition betweenandand implications for strawberry aroma patterns., 2013, 150: 47-53.
[23] 馬微, 牛瑩瑩, 駱強(qiáng)偉, 孫峰, 伍國(guó)紅, 廖康. 吐魯番地區(qū)溫室與露地栽培葡萄枝葉生長(zhǎng)及果實(shí)品質(zhì)差異分析. 新疆農(nóng)業(yè)科學(xué), 2016, 53(7): 1204-1209.
MA W, NIU Y Y, LUO Q W, SUN F, WU G H, LIAO K. Variance analysis on vegetative organ growth and fruit quality of grape in the greenhouses and open field in Turpan., 2016, 53(7): 1204-1209. (in Chinese)
[24] SAMYKANNO K, PANG E, MARRIOTT P J. Genotypic and environmental effects on flavor attributes of ‘Albion’ and ‘Juliette’ strawberry fruits., 2013, 164: 633-642.
[25] KAFKAS E, TüREMIS N, BILGILI B, ZARIFIKHOSROSHAHI M, BURGUT A, KAFKAS S. Aroma profiles of organically grown ‘Benicia’ and ‘Albion’ strawberries., 2017, 1156: 703-708.
[26] 謝劍平. 煙草香原料. 北京: 化學(xué)工業(yè)出版社, 2009: 261-355.
XIE J P.. Beijing: Chemical Industry Press, 2009: 261-355. (in Chinese)
[27] PINO J A, MESA J. Contribution of volatile compounds to mango (L.) aroma., 2006, 21(2): 207-213.
[28] JETTI R R, YANG E, KURNIANTA A, FINN C, QIAN M C. Quantification of selected aroma-active compounds in strawberries by headspace solid-phase microextraction gas chromatography and correlation with sensory descriptive analysis., 2007, 72 (7): S487-S496.
[29] QIAN M C, WANG Y Y. Seasonal variation of volatile composition and odor activity value of ‘Marion’ (spp.) and ‘Thornless Evergreen’ (L.) blackberries., 2005, 70(1): C13-C20.
[30] NOGUEROL-PATO R, GONZáLEZ-BARREIRO C, CANCHO- GRANDE B, SIMAL-GáNDARA J. Quantitative determination and characterization of the main odourants ofl red wines., 2009, 117: 473-484.
[31] FERREIRA V, AZNAR M, LóPEZ R, CACHO J. Quantitative gas chromatography-olfactometry carried out at different dilutions of an extract. Key differences in the odor profiles of four high-quality Spanish aged red wines., 2001, 49(10): 4818-4824.
[32] FRANCO M, PEINADO R A, MEDINA M, MORENO J. Off-vine grape drying effect on volatile compounds and aromatic series in must from Pedro Ximénez grape variety., 2004, 52(12): 3905-3910.
[33] WU Y S, DUAN S Y, ZHAO L P, GAO Z, LUO M, SONG S R, XU W P, ZHANG C X, MA C, WANG S P. Aroma characterization based on aromatic series analysis in table grapes., 2016, DOI: 10.1038/srep31116.
[34] ACREE T, ARN H. Flavornet and human odor space: Gas chromatography-olfactometry (GCO) of natural products. 2010, http://www.flavornet.org/flavornet.html.
[35] MOYANO L, ZEA L, MORENO J, MEDINA M. Analytical study of aromatic series in sherry wines subjected to biological aging., 2002, 50(25): 7356-7361.
[36] GENOVESE A, LAMORTE S A, GAMBUTI A, MOIO L. Aroma of Aglianico and Uva di Troia grapes by aromatic series., 2013, 53(1): 15-23.
[37] KIM Y, LEE K G, KIM M K. Volatile and non-volatile compounds in green tea affected in harvesting time and their correlation to consumer preference., 2016, 53(10): 3735-3743.
[38] RICCI A, CIRLINI M, LEVANTE A, DALL’ASTA C, GALAVERNA G, LAZZI C. Volatile profile of elderberry juice: Effect of lactic acid fermentation using,andstrains., 2018, 105: 412-422.
[39] WANG L B, BAI J H, YU Z F. Responses of volatile compounds in inner tissues on refrigeration in full ripe tomatoes., 2017: 41: e13272, DOI: 10. 1111/jfpp. 13272
[40] PEINADO R A, MORENO J, BUENO J E, MORENO J A, MAURICIO J C. Comparative study of aromatic compounds in two young white wines subjected to pre-fermentative cryomaceration., 2004, 84: 585-590.
[41] PEINADO R A, MAURICIO J C, MORENO J. Aromatic series in sherry wines with gluconic acid subjected to different biological aging conditions byvar.., 2006, 94: 232-239.
[42] CHEN S X, ZHANG R R, HAO L N, CHEN W F, CHENG S Q. Profiling of volatile compounds and associated gene expression and enzyme activity during fruit development in two cucumber cultivars., 2015, DOI: 10.1371/journal.pone.0119444.
[43] CNA’ANI A, MüHLEMANN J K, RAVID J, MASCI T, KLEMPIEN A, NGUYEN T T H, DUDAREVA N, PICHERSKY E, VAINSTEIN A.×floral scent production is negatively affected by high-temperature growth conditions., 2015(38): 1333-1346.
[44] CHENG S H, FU X M, MEI X, ZHOU Y, DU B, WATANABE N, YANG Z Y. Regulation of biosynthesis and emission of volatile phenylpropanoids/benzenoids in×flowers by multi- factors of circadian clock, light, and temperature., 2016, 107: 1-8.
[45] ANDRADE H B, BRAGA A F, BERTOLUCCI S K V, HSIE B S, SILVA S T, PINTO J E B P. Effect of plant growth regulators, light intensity and LED on growth and volatile compound of(L.) poitplantlets., 2017, 1155: 277-284.
[46] COLQUHOUN T A, SCHWIETERMAN M L, GILBERT J L, JAWORSKI E A, LANGER K M, JONES C R, RUSHING G V, HUNTER T M, OLMSTEAD J, CLARK D G, FOLTA K M. Light modulation of volatile organic compounds from petunia flowers and select fruits., 2013, 86: 37-44.
[47] SAGAE M, OYAMA-OKUBO N, ANDO T, MARCHESI E, NAKAYAMA M. Effect of temperature on the floral scent emission and endogenous volatile profile of., 2008, 72: 110-115.
[48] FARRé-ARMENGOL G, FILELLA I, LLUSIà J, NIINEMETS ü, PE?UELAS J. Changes in floral bouquets from compound-specific responses to increasing temperatures., 2014, 20: 3660-3669.
[49] FOLKERS A, HüVE K, AMMANN C, DINDORF T, KESSELMEIER J, KLEIST E, KUHN U, UERLINGS R, WILDT J. Methanol emissions from deciduous tree species: dependence on temperature and light intensity., 2008, 10: 65-75.
[50] WENDA-PIESIK A. Volatile organic compound emissions by winter wheat plants (L.) underspp. Infestation and various abiotic conditions., 2011, 20(5): 1335-1342.
[51] GOH H H, KHAIRUDIN K, SUKIRAN N A, NORMAH M N, BAHARUM S N. Metabolite profiling reveals temperature effects on the VOCs and flavonoids of different plant populations., 2016, 18(S1): 130-139.
[52] UBI B E, HONDA C, BESSHO H, KONDO S, WADA M, KOBAYASHI S, MORIGUCHI T. Expression analysis of anthocyanin biosynthetic genes in apple skin: Effect of UV-B and temperature., 2006, 170: 571-578.
[53] BOATRIGHT J, NEGRE F, CHEN X L, KISH C M, WOOD B, PEEL G, ORLOVA I, GANG D, RHODES D, DUDAREVA N. Understandingbenzenoid metabolism in petunia petal tissue., 2004, 135: 1993-2011.
[54] PROCISSI A, DOLFINI S, RONCHI A, TONELLI C. Light- dependent spatial and temporal expression of pigment regulatory genes in developing maize seeds., 1997, 9: 1547-1557.
[55] TAKOS A M, JAFFé F W, JACOB S R, BOGS J, ROBINSON S P, WALKER A R. Light-induced expression of agene regulates anthocyanin biosynthesis in red apples., 2006, 142: 1216-1232.
[56] BOREVITZ J O, XIA Y, BLOUNT J, DIXON R A, LAMB C. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis., 2000, 12: 2383-2394.
[57] ZVI M M B, NEGRE-ZAKHAROV F, MASCI T, OVADIS M, SHKLARMAN E, BEN-MEIR H, TZFIRA T, DUDAREVA N, VAINSTEIN A. Interlinking showy traits: co-engineering of scent and colour biosynthesis in flowers., 2008, 6: 403-415.
[58] ZVI M M B, SHKLARMAN E, MASCI T, KALEV H, DEBENER T, SHAFIR S, OVADIS M, VAINSTEIN A.transcription factor enhances production of phenylpropanoid and terpenoid scent compounds in rose flowers., 2012, 195: 335-345.
[59] SHEN X J, ZHAO K, LIU L L, ZHANG K C, YUAN H Z, LIAO X, WANG Q, GUO X W, LI F, LI T H. A role for PacMYBA in ABA-regulated anthocyanin biosynthesis in red-colored sweet cherry cv. Hong Deng (L.)., 2014, 55(5): 862-880.
[60] RAYMOND O, GOUZY J, JUST J, BADOUIN H, VERDENAUD M, LEMAINQUE A, VERGNE P, MOJA S, CHOISNE N, PONT C, CARRèRE S, CAISSARD J, COULOUX A, COTTRET L, AURY J, SZéCSI J, LATRASSE D, MADOUI M, FRAN?OIS L, FU X P, YANG S H, DUBOIS A, PIOLA F, LARRIEU A, PEREZ M, LABADIE K, PERRIER L, GOVETTO B, LABROUSSE Y, VILLAND P, BARDOUX C, BOLTZ V, LOPEZ-ROQUES C, HEITZLER P, VERNOUX T, VANDENBUSSCHE M, QUESNEVILLE H, BOUALEM A, BENDAHMANE A, LIU C, BRIS M L, SALSE J, BAUDINO S, BENHAMED M, WINCKER P, BENDAHMANE M. Thegenome provides new insights into the domestication of modern roses., 2018. https://doi. org/10.1038/s41588- 018-0110-3.
[61] XU X Q, LIU B, ZHU B Q, LAN Y B, GAO Y, WANG D, REEVES M J, DUAN C Q. Differences in volatile profiles of Cabernet Sauvignon grapes grown in two distinct regions of China and their responses to weather conditions., 2015, 89: 123-133.
[62] DUAN L L, PAN Q H, TANG X J, YANG Q, JIANG R, SHI Y, DUAN C Q. Characteristic aroma compounds in two newcultivars (table grapes) and impact of vintage and greenhouse cultivation., 2014, 35(2): 264-277.
[63] LEE J H J, JAYAPRAKASHA G K, RUSH C M, CROSBY K M, PATIL B S. Production system influences volatile biomarkers in tomato., 2018, 14: 99.
[64] FRIEDEL M, FROTSCHER J, NITSCH M, HOFMANN M, BOGS J, STOLL M, DIETRICH H. Light promotes expression of monoterpene and flavonol metabolic genes and enhances flavor of winegrape berries (L. cv. Riesling)., 2016, 22(3): 409-421.
[65] PASCUAL G A, SERRA I, CALDERóN-ORELLANA A, LAURIE V F, LOPéZ M D. Changes in concentration of volatile compounds in response to defoliation of Muscat of Alexandria grapevines grown under a traditional farming system., 2017, 77(4): 373-381.
[66] ZHANG E, CHAI F, ZHANG H, LI S, LIANG Z, FAN P. Effects of sunlight exclusion on the profiles of monoterpene biosynthesis and accumulation in grape exocarp and mesocarp., 2017, 237: 379-389.
[67] MILLER T W, FELLMAN J K, MATTHEIS J P, MATTINSON D S. Factors that influence volatile ester biosynthesis in ‘Delicious’ apples., 1998, 464: 195-200.
[68] SANZ C, PéREZ A G, OLíAS R. Effects of temperature on flavor components in “” grown strawberry., 2002, 567: 365-368.
[69] EDGER P P, POORTEN T J, BUREN R V, HARDIGAN M A, COLLE M, MCKAIN M R, SMITH R D, TERESI S J, NELSON A D L, WAI C M, ALGER E I, BIRD K A, YOCCA A E, PUMPLIN N, OU S J, BEN-ZVI G, BRODT A, BARUCH K, SWALE T, SHIUE L, ACHARYA C B, COLE G S, MOWER J P, CHILDS K L, JIANG N, LYONS E, FREELING M, PUZEY J R, KNAPP S J. Origin and evolution of the octoploid strawberry genome., 2019, https://doi.org/10.1038/s41588-019-0356-4.
[70] ZHANG B, XI W P, WEI W W, SHEN J Y, FERGUSON I, CHEN K S. Changes in aroma-related volatiles and gene expression during low temperature storage and subsequent shelf-life of peach fruit., 2011, 60: 7-16.
[71] SCHREIER P. Quantitative composition of volatile constituents in cultivated strawberries,cv. Senga sengana, Senga litessa, and Senga gourmella., 1980, 31: 487-494.
[72] HAKALA M A, LAPVETELAINEN A T, KALLIA H P. Volatile compounds of selected strawberry varieties ananlyzed by purge-and- trap headspace GC-MS., 2002, 50: 1133-1142.
[73] OLBRICHT K, GRAFE C, WEISS K, ULRICH D. Inheritance of aroma compounds in a model population ofDuch., 2008, 127: 87-93.
[74] OLBRICHT K, ULRICH D. Domestication effects in European breeding history of strawberry demonstrated by aroma compound pattern., 2017, 1156: 61-68.
Comparative Study on Fruit Volatiles of Different Day-neutral Strawberry Cultivars in Autumn and Winter
DONG Jing, ZHONG ChuanFei, WANG GuiXia, CHANG LinLin, SUN Jian, SUN Rui, ZHANG HongLi, LI Rui, WEI YongQing, ZHENG ShuQi, ZHANG YunTao
(Beijing Academy of Forestry and Pomology Sciences/Beijing Engineering Research Center for Strawberry/Key Laboratory of Biology and Genetic Improvement of Horticultural Crops (North China), Ministry of Agriculture, Beijing 100093)
【】Fruit volatiles of day-neutral strawberries harvested in autumn and winter were compared to clarify the effects of seasonal changes on volatile composition and odor activity value (OAV) of volatiles, aiming to provide theoretical basis for research on strawberry aroma and utilization of day-neutral cultivars. 【】Matured fruits of day-neutral cultivars (including Albion, Monterey, Portola, and San Andreas) were harvested from high-tunnel in autumn and greenhouse in winter, respectively. Volatiles were extracted by using solid-phase microextraction (SPME), detected by gas chromatograph-mass spectrometer (GC-MS) and analyzed qualitatively and quantitatively. The numbers, contents, percentages and OAVs of volatiles were calculated. Principal component analysis (PCA) was employed to analyze the differences in volatiles content between autumn and winter strawberries. Clustered heatmap was used to classify the samples depending on the volatiles with OVA higher than 1. 【】A total of 88 individual volatiles were identified and the OAVs of 57 volatiles were calculated. It was found that there were 30 components with OAV higher than 1. Effects of seasonal changes on the number of each chemical category and the total content of volatiles varied among cultivars. Compared to the autumn fruits, the content and percentage of esters in winter fruits were significantly higher, while terpenes were remarkably lower. The result of PCA indicated that, compared with other cultivars, volatiles pattern of Portola showed less variation, whereas San Andreas showed the highest variation between two seasons. Ethyl butanoate was very important for these tested cultivars since its average OAV was the highest in all volatiles. The OAVs of butyl acetate and methyl butanoate in winter fruits were noticeably higher than that in autumn fruits. However, compared with autumn fruits, the winter fruits of each cultivar had a lower sum of terpene OAVs that were higher than 1. Ethyl butanoate, butyl butanoate, 4-methoxy-2, 5-dimethyl- 3()-furanone, ethyl hexanoate and linalool played an important role in the aroma differences between fruits of different seasons. The eight tested samples were classified into 3 groups using clustered heatmap. The group I included Albionautumn, Albionwinter and Montereyautumn, of which total OAVs and the OAVs of ethyl butanoate were significantly higher than other groups.【】Seasonal change had a remarkable impact on esters and terpenes in fruit volatiles of day-neutral strawberry cultivars. It was showed that the content and percentage of esters in winter fruits were significantly higher than autumn fruits, while the percentage of terpenes was remarkably lower in winter fruits. Among the 4 tested cultivars, Portola showed the least variation in volatile content between autumn and winter. Albion was the cultivar with intense aroma and its aroma performance presented the least changes when season changed, however, San Andreas was the one with the largest volatile variation both in volatile content and fruit aroma.
×; day-neutral cultivars; fruit volatiles composition; odor activity value; esters; terpenes
10.3864/j.issn.0578-1752.2019.13.010
2019-01-18;
2019-05-13
國(guó)家重點(diǎn)研發(fā)計(jì)劃項(xiàng)目(2016YFE0112400)、國(guó)家自然科學(xué)基金(31601714)、北京市糧經(jīng)作物產(chǎn)業(yè)創(chuàng)新團(tuán)隊(duì)項(xiàng)目(BAIC09-2019)
董靜,E-mail:dj310730@sohu.com。
張運(yùn)濤,E-mail:zhytao1963@126.com
(責(zé)任編輯 趙伶俐)
