中華鱉NR5a2基因的克隆與表達分析
2019-07-26 06:58:18吳利敏劉慧芬李學軍王璐明
淡水漁業 2019年4期
張 超,馬 曉,劉 倩,吳利敏,劉慧芬,李學軍,王璐明
(河南師范大學水產學院,河南新鄉 453007)
NR5a2(nuclear receptor subfamily 5,group A,member 2)又稱LRH-1(肝受體同系物),是一種孤核受體,也是需要配體激活的轉錄因子。在小鼠[1](Musmusculus)、牙鲆[2](Paralichthysolivaceus)、赤點石斑魚[3](Epinephelusakaara)中發現NR5a2可以結合Cyp19a1a啟動子,進而激活其轉錄活性。此外,在牙鲆中還發現NR5a2可以與Foxl2協同作用,調節Cyp19a1a的轉錄表達[2]。當NR5a2僅在赤點石斑魚成熟卵巢細胞細胞質中表達時,Cyp19a1a基因的表達水平下降[3-4]。Cyp19a1a基因編碼的芳香化酶是雄激素轉換為雌激素的關鍵限速酶,而雌激素在脊椎動物卵巢分化,雌性性別特征的維持中有重要的作用。NR5a2作為Cyp19a1a轉錄因子之一,對Cyp19a1a基因的表達也有一定的調節作用,深入研究NR5a2和Cyp19a1a在性別分化中的功能具有重要意義。
中華鱉(Pelodiscussinensis),又稱甲魚、水魚、團魚等,隸屬于龜鱉目(Testudinate)鱉科(Tironychidae)中華鱉屬(Pelodiscus),是我國廣泛養殖的名貴水產品之一,具有較高的營養價值和藥用價值。目前對中華鱉的研究多集中在性別決定與分化、免疫和營養等方面[5-7]。爬行動物的性別決定有基因型性別決定(GSD)和環境性別決定類型(ESD)。前期研究多認為中華鱉屬于ESD中的TSD類型,隨著研究發展深入,越來越多證據表明中華鱉屬于GSD型[8]。養殖過程中,雄鱉比雌鱉生長速度快。因此,深入研究中華鱉性別分化與發育機制,有助于建立提高雄鱉孵化率或全雄鱉培育方法,對中華鱉養殖業發展有重要意義。本實驗克隆了中華鱉NR5a2基因,通過實時熒光定量PCR技術研究中華鱉NR5a2基因在不同組織以及不同時期胚胎中的表達,為進一步研究其功能提供依據,也為中華鱉性別發育研究提供資料。
1 材料與方法
1.1 實驗用中華鱉組織及胚胎的收集
本實驗所選用的成熟雌性/雄性中華鱉和受精卵采自固始縣晨源水產養殖合作社,隨機選取成熟雌性、雄性中華鱉各6只,分別取心臟、肝臟、脾臟、腦、胃、肌肉、精巢、卵巢組織置于無酶管中,后將組織存放在-80 ℃冰箱備用。受精卵通過恒溫恒濕培養箱進行孵化,孵化溫度為31.5 ℃,濕度控制在75%~90%。孵化過程中根據中華鱉胚胎發育圖譜[9]收集性腺分化前后不同時期(16-23期)[10-11]性腺-中腎復合體(AKG),每個時期取6個樣品。
1.2 中華鱉NR5a2基因全長的克隆
1.2.1 引物設計
根據Ensemble數據庫中的中華鱉NR5a2基因cDNA序列(ENSPSIG00000003632.1)設計引物F1/R1進行cDNA序列的驗證,同時設計GSP 5′/NP5′和GSP3′/NP3′引物用于基因全長的克隆(表1)。此外,根據克隆得到的中華鱉NR5a2基因cDNA全長設計特異性引物用于熒光定量PCR表達實驗。在熒光定量PCR實驗中選用的內參基因為GAPDH[9,12]。引物通過Primer 5.0 設計,由上海生工生物技術有限公司合成。

表1 實驗中所用引物序列
1.2.2 RNA的提取和NR5a2的克隆
取卵巢組織以及不同發育時期個體AKG,RNAiso plus(TaKaRa,大連)提取總RNA。ND-2000核酸蛋白儀測定RNA濃度與OD值,瓊脂糖凝膠電泳檢測RNA完整性,總RNA置于-80 ℃保存。按照Prime ScriptTMII 1st Strand cDNA Synthesis Kit試劑盒(TaKaRa,大連)的操作說明進行中華鱉卵巢總RNA反轉錄,通過特異性引物NR5a2-F1/R1擴增NR5a2 cDNA部分序列。在已獲得部分序列的基礎上設計3′ RACE 和5′RACE的基因特異性引物(表1),按照SMART 5′ RACE & 3′ RACE 試劑盒(TaKaRa,大連)說明書合成3′-RACE-Ready cDNA和5′-RACE-Ready cDNA,分別進行3′非編碼區和5′非編碼區的擴增。將擴增結果進行克隆、測序,獲得所需片段并進行拼接,得到該基因cDNA序列全長。
1.3 中華鱉NR5a2基因的序列分析
利用DNAman軟件對中華鱉NR5a2基因的測序結果進行拼接;利用Open Reading Frame(ORF)Finder(https://www.ncbi.nlm.nih.gov/orffinder/)推算其開放閱讀框及編碼的氨基酸序列;通過SignalP 4.1 serves(http://www.cbs.dtu.dk/services/SignalP/)預測其信號肽結合位點; SMART(http://smart.embl-heidelberg.de/)進行蛋白質結構域分析;使用ClustalX2進行氨基酸序列同源性比對;運用MAGA7.0軟件通過鄰接法(NJ法)(bootstrap values為1 000)構建系統進化樹。
1.4 qRT-PCR分析中華鱉NR5a2基因的組織特異性表達
按照Prime ScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)試劑盒(TaKaRa,大連)操作說明進行不同發育時期性腺中腎復合體RNA的反轉錄,選用特異性引物qNR5a2-F/R(表1)進行qRT-PCR,以GAPDH為內參基因(表1)。反應體系(10 μL):TB Green Master Mix 5 μL,上下游引物各0.4 μL,cDNA模板1 μL,ddH2O 3.2 μL;PCR反應程序:95 ℃ 預變性30 s,95 ℃ 變性5 s,58 ℃ 退火30 s,40個循環,每個組織分別取自3只中華鱉,即3個平行。采用2-△△Ct法計算中華鱉NR5a2基因的相對表達量,結果采用SPSS16.0軟件進行ANOVA單因素方差分析,以平均值±標準誤(Mean±SD)表示。
2 結果分析
2.1 中華鱉NR5a2基因序列分析
本實驗克隆得到中華鱉NR5a2基因cDNA序列全長為2 674 bp,開放閱讀框長1 608 bp,編碼535個氨基酸,5′-UTR長40 bp,3′-UTR長1 026 bp(圖1)。使用ProtParam預測該基因編碼的蛋白質分子質量為60.5 kDa,等電點為8.57。

圖1 中華鱉NR5a2基因序列和氨基酸序列Fig.1 Full-length nucleotide cDNA sequences and deduced amino acid sequences of NR5a2 in P.sinensis小寫字母表示5′和3′非編碼區;大寫字母表示編碼區。預測的結構域ZnF-C4用下劃線表示,預測的結構域HOLI由陰影區表示,*表示終止密碼子。
對中華鱉NR5a2編碼的氨基酸序列進行分析,發現該序列無明顯信號肽位點,也不存在蛋白跨膜區域。在疏水性/親水性分析中發現其疏水性殘基比例大于親水性殘基比例,其中疏水性最大值為-2.778,親水性最大值為2.411。SMART分析表明中華鱉NR5a2含有一個保守的ZnF-C4結構域和一個保守的HOLI結構域(圖2-A)。通過 DNASTAR.Lasergene.v7.1 軟件中的Protean程序對中華鱉NR5a2基因蛋白質二級結構組成進行預測,其蛋白質二級結構中的Alpha螺旋、Beta折疊、Turn轉角以及Coil卷曲螺旋分布如圖所示(圖2-B),結果表明該蛋白質二級結構主要由Alpha螺旋和Coil卷曲螺旋構成,分別占45.61%和45.05%。SWISS-MODEL在線預測軟件對中華鱉NR5a2基因編碼蛋白質的三級結構進行預測結果如圖所示(圖2-C)。
2.2 中華鱉NR5a2基因氨基酸同源性分析及系統發育樹的構建
使用Megalign軟件進行中華鱉NR5a2氨基酸序列與其他物種NR5a2氨基酸進行比對(圖3)。結果表明中華鱉與西部錦龜的相似性最高,為99.2%,其次是原雞,相似性為98.4%(表2)。通過MEGA5.1軟件,采用鄰接(Neighbor-Joining)法構建系統發育樹(圖4)。結果顯示中華鱉與西部錦龜的進化關系最近,其次是原雞、紫翅瓊鳥和揚子鱷。
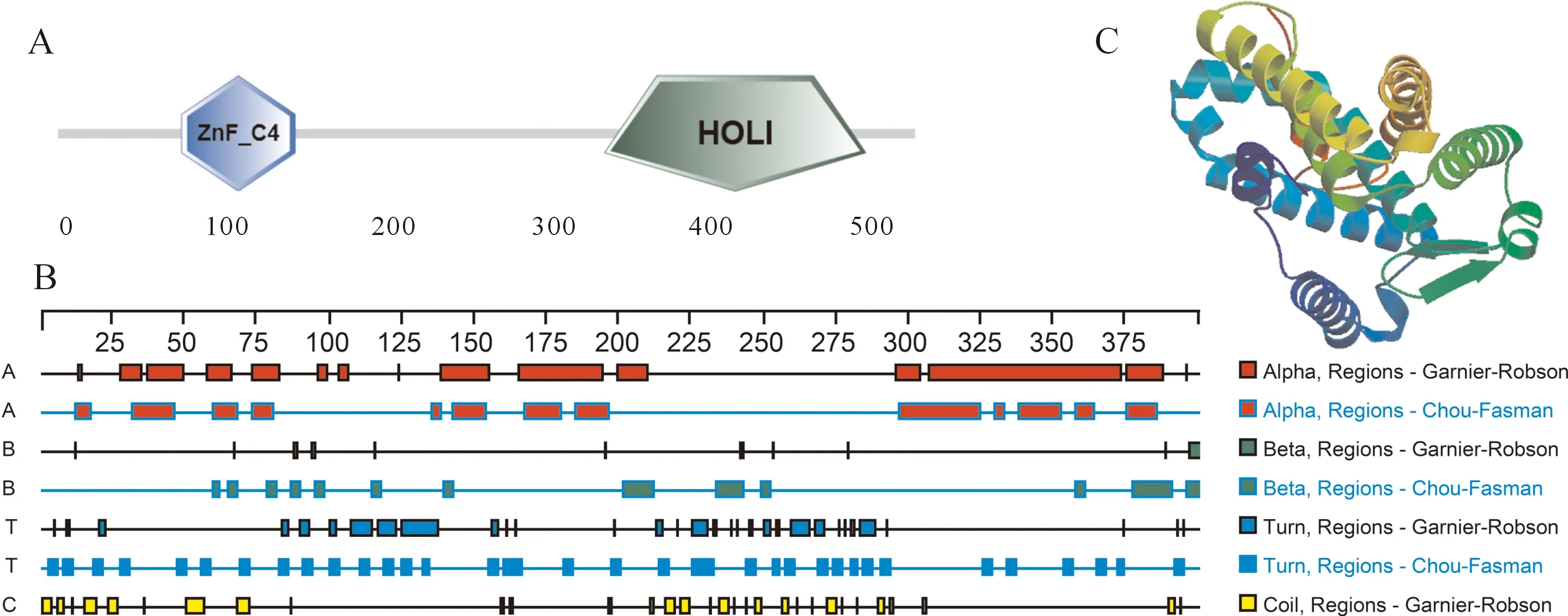
圖2 NR5a2編碼氨基酸序列分析Fig.2 The sequence analysis of protein of NR5a2A:NR5a2編碼蛋白結構域預測;B:NR5a2編碼蛋白質二級結構預測;C:NR5a2編碼蛋白質三級結構預測。

圖3 不同物種NR5a2基因氨基酸同源性比較Fig.3 Alignment of amino acid sequences of NR5a2 gene in different species使用Megalign軟件進行多序列對比,其中黑色底紋代表氨基酸相似度為100%,灰色底紋代表氨基酸相似度為75%。
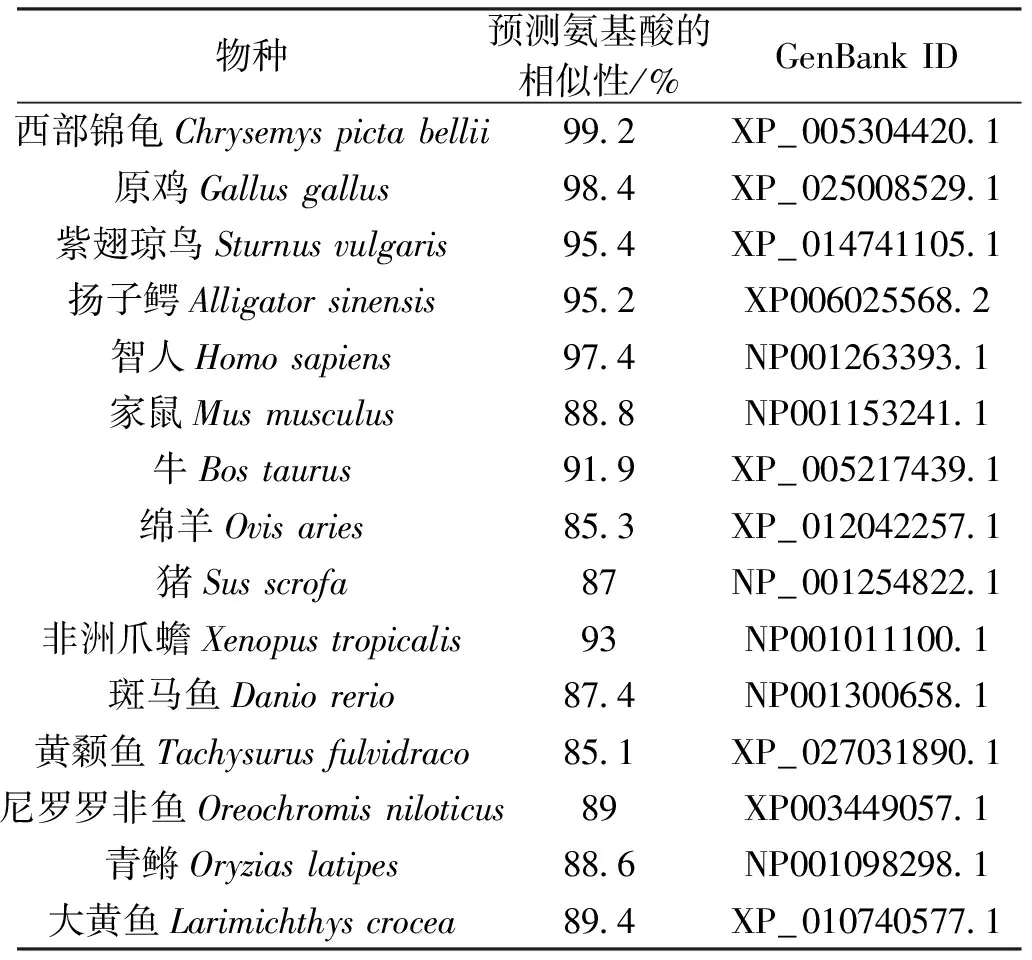
表2 預測的中華鱉NR5a2氨基酸序列與其他物種NR5a2氨基酸序列的比對
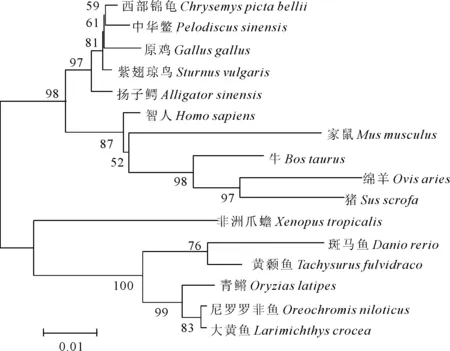
圖4 NR5a2基因編碼氨基酸序列系統發育樹Fig.4 Phylogenetic tree based amino acid sequences of NR5a2
2.3 中華鱉NR5a2基因的組織特異性表達及不同發育階段變化
熒光定量PCR結果表明,NR5a2基因在中華鱉的心臟、肝臟、脾臟、腦、胃、肌肉、精巢、卵巢中均表達。其中,在肝臟中表達量最高,其次是卵巢,在肝臟和卵巢組織中NR5a2的相對表達量顯著高于其他組織(P<0.05)(圖5)。31.5 ℃條件下孵育,在中華鱉胚胎發育不同時期均在AKG中檢測到NR5a2表達,在胚胎發育17期(性腺分化開始階段)時表達量最高,隨著發育的進程表達量顯著下降(圖6)。
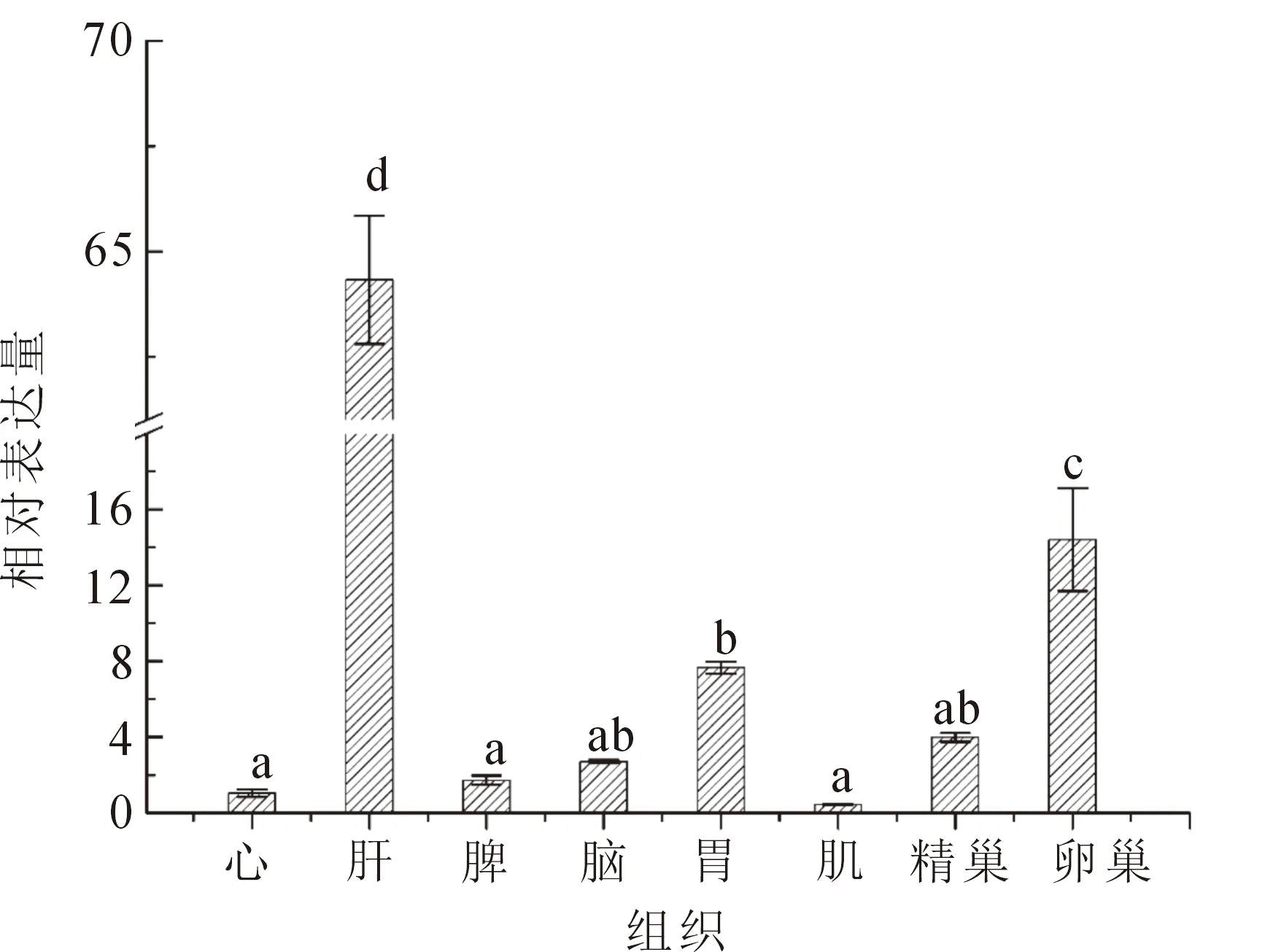
圖5 NR5a2基因在中華鱉不同組織中的相對表達量Fig.5 Real-time PCR quantification of NR5a2 expression in different tissues of P.sinensis不同小寫字母表示NR5a2基因在不同組織中的差異顯著性(P<0.05)。
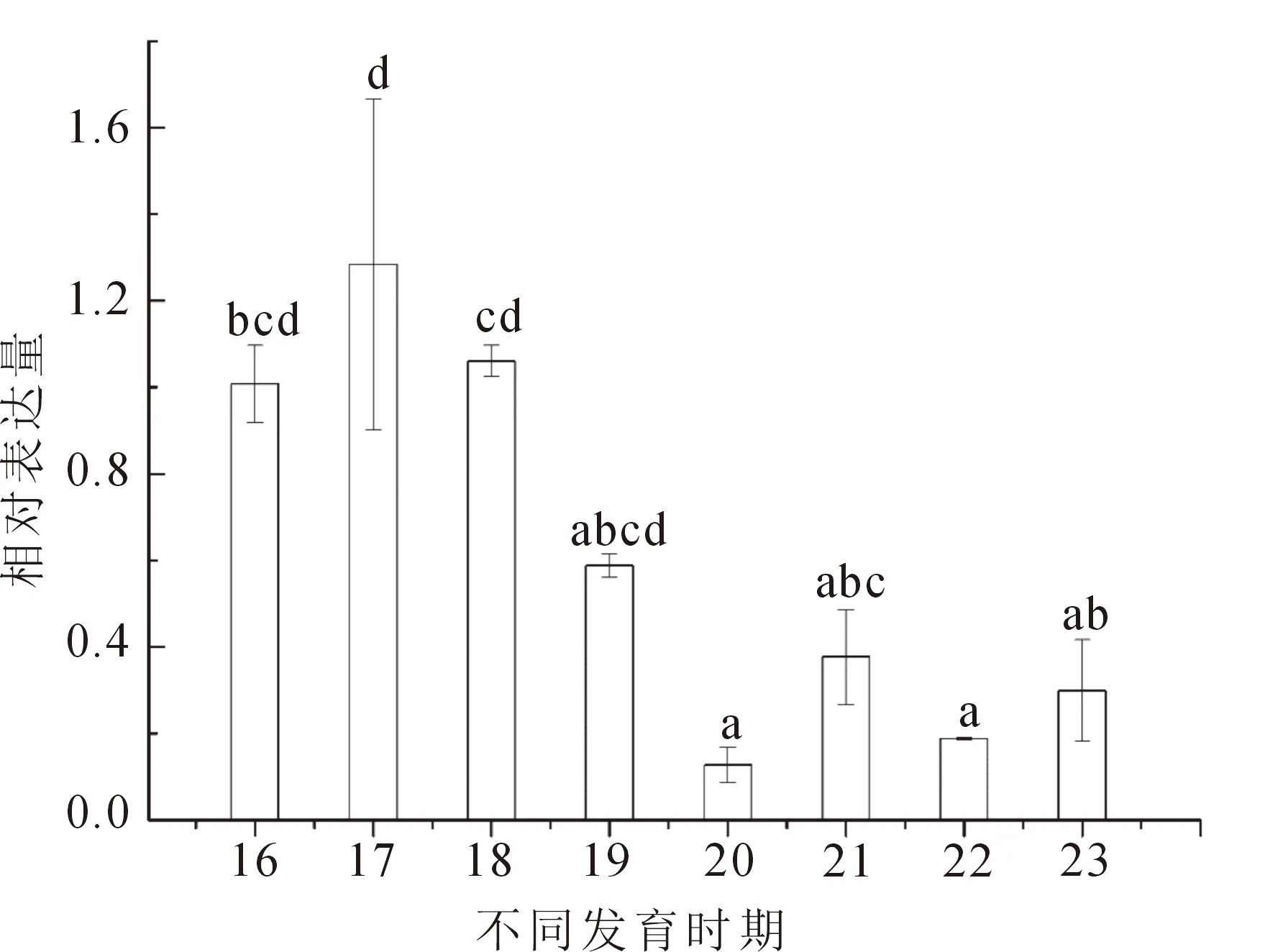
圖6 NR5a2基因在中華鱉胚胎不同發育時期的相對表達量(孵化溫度:31.5 ℃)Fig.6 The relative expression of NR5a2 gene at different developmental stages of P.sinensis embryo( incubation temperature :31.5 ℃)不同小寫字母表示NR5a2基因在不同發育時期的差異顯著性(P<0.05)。
3 討論
本實驗通過RACE方法克隆得到中華鱉NR5a2基因cDNA序列全長,共2 674 bp,其ORF框長度為1 608 bp,編碼535個氨基酸,具有一個ZnF-C4結構域和一個HOLI結構域,這與二花臉豬[13](Erhualianpig)、黃顙魚[14](Pelteobagrusfulvidraco)等物種中的研究結果類似。系統進化樹分析表明中華鱉NR5a2編碼氨基酸序列與西部錦龜、雞、揚子鱷等物種具有較高的相似性,親緣關系較近,和斑馬魚(Daniorerio)、青鳉(Oryziaslatipes)、尼羅羅非魚(Oreochromisniloticus)等物種的相似性較低,親緣關系較遠,這一結果與中華鱉在不同物種中的分類地位表現一致。
NR5a2是核受體超家族中的重要一員,在生長發育、細胞分化等方面發揮重要的作用,研究發現該基因主要在肝胰臟以及卵巢組織中表達。NR5a2可與7-α-羥化酶基因啟動子區特異性結合并激活其轉錄,平衡動物體內膽汁酸水平[15]。Falender等[16]在嚙齒動物卵巢研究表明NR5a2可以調節雌激素的合成。Duggavathi等[17]敲除小鼠顆粒細胞NR5a2后發現卵巢中雌激素含量降低,排卵量減少。姚勇等[13]研究指出NR5a2在二花臉豬卵巢組織中高表達,可能促進卵泡的形成和排卵,參與生殖活動。本實驗通過熒光定量PCR技術檢測NR5a2在中華鱉不同組織中的表達情況,結果顯示,NR5a2在各組織中均有表達,在肝臟組織和卵巢組織中表達量較高,這表明NR5a2可能在中華鱉生殖功能和肝功能調控中有重要作用。
性別分化是水產動物研究熱點之一,Cyp19a1a是雌激素合成的關鍵基因,對動物的性別分化與發育,尤其是卵巢的分化和功能維持有重要作用。在虹鱒(Oncorhynchusmykiss)[4]、赤點石斑魚[18]等研究中,NR5a2可作為轉錄因子與Cyp19a1a啟動子結合,調控Cyp19a1a轉錄表達從而調節雌激素水平。在牙鲆[19]中,NR0b1和NR5a2可通過調控Cyp19a1a的表達間接調節E2水平,參與牙鲆性腺分化。此外,Yan等[20]在河豚(Tetraodontidae)性別分化關鍵時期檢測到NR5a2高表達。王莉等[11]、葛楚天等[11]研究表明,中華鱉性腺分化時間為胚胎發育17期。本實驗通過qRT-PCR檢測NR5a2在中華鱉胚胎不同發育時期的表達情況。結果表明,NR5a2在胚胎發育16期(性腺分化之前)已表達,在 17期(性腺分化初期)表達水平最高,隨后表達水平下降。Cyp19a1在胚胎發育17期開始顯著增加[12],推測NR5a2可能作為轉錄因子參與調控Cyp19a1表達,從而參與中華鱉性腺分化調控。
