新疆野蘋果次生代謝產物對蟲害脅迫的響應
2019-07-18 00:56:44艾沙江買買提朱燕飛韓立群王繼勛
新疆農業科學 2019年4期
梅 闖,閆 鵬,艾沙江·買買提,朱燕飛,馬 凱,韓立群,王繼勛
(新疆農業科學院園藝作物研究所/農業部新疆地區果樹科學觀測試驗站,烏魯木齊 830091)
0 引 言
【研究意義】新疆野蘋果(Malussieversii),分布于天山山脈,同源性高且最原始[1],其花、果實、營養成分、生長習性具有豐富的多樣性[2],遺傳基因豐富可促進蘋果品種的選育[3]。蘋果小吉丁蟲(AgrilusmaliMats.),屬鞘翅目,是蘋果毀滅性鉆蛀害蟲,其幼蟲隱藏于樹干或枝干皮層內盤旋蛀食,致使木質部和韌皮部分離,造成枝干枯死,為害嚴重時造成果樹整株死亡[4]。中國多省份零星分布為害,1995年在新疆發現,并入侵至天山野果林,快速繁殖并最終暴發成災[5],被我國定為高度危險有害生物,該蟲害枯死野蘋果達到600多公頃[6]。栽培蘋果是世界上最重要的水果之一,具有自交不親和,無性繁殖的特點[7],這些特征導致新疆野蘋果重要基因消亡后,將不可再生。研究新疆野蘋果次生代謝物對蟲害脅迫的響應,對保護利用新疆野蘋果資源有重要意義。【前人研究進展】昆蟲取食寄生植物后,會誘導植物產生系統性的抗蟲響應,表現為受損植物細胞信號輸入,隨后進行信號轉導(鈣離子通量,磷酸化級聯和激素串擾)和信號處理(轉錄組,蛋白質組和代謝組的重構),隨后發生信號響應(防御化合物,蛋白質和次級代謝產物的產生)[8, 9]。次生代謝物質是復雜的分支代謝途徑產物,不直接參與維持植物的生長發育和生殖有關的基礎生化活動,但可作為信號分子,或直接參與化學防御,包括酚類化合物、萜類化合物、含氮有機堿類化合物、硫代葡萄糖苷等[10]。同時可以利用植物自身的防御代謝物質合成植物源殺蟲劑消滅害蟲[11]。擬南芥中研究表明,硫代葡萄糖苷構成了植物-昆蟲互作主要的防御性狀,作為對昆蟲取食的響應,MYR催化硫代葡萄糖苷的氰基化以釋放毒性化合物,例如,腈,異硫氰酸酯和硫氰酸甲酯[12]。柳枝稷在受到蚜蟲侵染后,其體內的ROS和多種次級代謝產物發生顯著變化,并伴隨著NAC,WRKY和MYB轉錄因子的上調表達[13]。研究發現,不同品系楊樹咖啡酸、肉桂酸、丁香酸對青楊脊虎天牛(Xylotrechusrusticus)的驅避效果明顯[14]。另外黃酮類物質也對昆蟲有毒性,可以影響昆蟲的行為和代謝,或破壞昆蟲的正常代謝過程,對其生長、發育和繁殖均有明顯的抑制作用[15]。【本研究切入點】研究新疆野蘋果受到蟲害誘導前后某些次生代謝物質的變化情況及其產生規律。【擬解決的關鍵問題】以經過前期鑒定穩定遺傳的抗蟲株系為材料,利用液相色譜-質譜(LC-MS)聯用技術,定性定量測定次生代謝物質含量變化趨勢,研究新疆野蘋果(Malussieversii)受到蟲害脅迫誘導后相關次生代謝物質的變化及其產生規律,為保護利用新疆野蘋果資源提供理論依據。
1 材料與方法
1.1 材 料
2011~2016年在實地調查新疆新源縣蘋果小吉丁蟲疫區新疆野蘋果資源,總計調查面積2 500余hm2,新疆野蘋果受蘋果小吉丁蟲危害嚴重(干枯死枝率達到90%以上),同海拔同一地點有5株新疆野蘋果受害較輕(干枯死枝率低于10%,且枝干生長旺盛)[16]從表型性狀分析該類單株可能具備一定的抗蟲能力,選定一株為害等級最低植株定為抗蟲株系[6]。在具有抗蟲株系新疆野蘋果周邊選取1株具有代表性的受侵害單株作為對照株系,樹齡在45~60 a。基于廣泛靶向代謝組技術平臺的代謝分析,檢測分析16個抗蟲代謝物,對代謝物的定量和差異分析,揭示新疆野蘋果在不同處理條件下抗蟲代謝物的代謝機制。表1
表1 檢測代謝物種類及數量
Table 1 Detecting the type and quantity of metabolites

ID英文名稱English name中文名稱Chinese NameMas01Theophylline茶堿Mas02Syringic acid丁香酸Mas03Chlorogenic acid綠原酸Mas04Vanillic acid香草酸Mas05Ferulic acid阿魏酸Mas06Caffeic acid咖啡酸Mas07p-Coumaric acid4-香豆酸Mas08Benzoic acid苯甲酸Mas09Quercetin3-O-a-L-rhamnoside (Quercitrin)槲皮素 3-O-鼠李糖苷 (槲皮苷)Mas10Quercetin槲皮素Mas11aloperine苦豆堿Mas12Theobromine咖啡堿Mas13Gramine蘆竹堿Mas14Salicylic acid (SA)水楊酸Mas15trans-Cinnamic acid反式肉桂酸Mas16Rutin蕓香甙
1.2 方 法
1.2.1 侵染處理
根據昆蟲危害習性,蘋果小吉丁蟲寄生于蘋果枝條韌皮部,蟲卵于上一年產于韌皮部,次年5月幼蟲開始為害,至7月為高峰,持續到9月。選擇經過多年鑒定,抗蟲表現穩定的新疆野蘋果單株。根據蘋果小吉丁蟲生育期,分別對兩種處理進行幼蟲侵染(模擬幼蟲孔道,接入成齡幼蟲),采集樣品,分別是侵染后10 d(機械傷害愈合且接入幼蟲取食距傷口>3 cm處),采集抗蟲株系與對照株系的健康部位和侵染部位韌皮部50 g,立即在液氮中冷凍并儲存在-80℃冰箱中,每個株系3個生物學重復,共計12個樣品。
1.2.2 樣品提取
取出超低溫冷凍保存的生物材料樣本,利用研磨儀(MM 400, Retsch)在30Hz條件下研磨1 min,稱取100 mg的粉末,利用80%甲醇1.0 mL于4℃提取過夜,期間渦旋3次,使提取更為充分。提取后,12 000 g離心15 min,吸取上清,35℃加熱模式下氮氣吹干,用100 μL 30%甲醇水復溶樣品,渦旋使目標物質充分溶解,12 000 g離心15 min,吸取上清,并保存在進樣瓶中用于LC-MS分析。
1.2.3 色譜質譜采集條件
參照Pan、Francescato和Wojakowska[17-19]的方法,采用液相色譜串聯質譜(LC-MS)分析方法測定次生代謝物含量。所有的化學試劑都是分析純或色譜純的制劑。化學標準品從BioBioPha公司(http://www.biobiopha.com/)與美國Sigma-Aldrich公司購買(http://www.sigmaaldrich.com/united-states.html)。分析條件主要包括:(1)色譜柱:WatersACQUITY UPLC HSS T3 C18 1.8 μm,2.1 mm ×100 mm。(2)流動相:水相為超純水(加入0.1%的甲酸),有機相為乙腈(加入0.1%的甲酸);(3)洗脫梯度,水:乙腈,0 min為95∶5 V/V,11.0 min為5∶95 V/V,12.0 min為5∶95 V/V,12.1 min為95∶5 V/V,15.0 min為95∶5 V/V;(4)流速為0.4 mL/min;(5)柱溫為40℃;(6)進樣量為5 μL。樣品經過色譜分離后進入到質譜進行分析。
配置不同濃度的標準品溶液,進行質譜分析,其中,標準品濃度包括0.1、0.2、0.5、1.0、2.0、5.0和10.0 μg/mL等。
1.3 數據處理
采用 Microsoft Excel 2010軟件對數據進行整理,使用 SPSS 17.0 軟件進行方差分析(P<0.05)及標準誤的計算。繪圖用 origin 8.5 軟件。
2 結果與分析
2.1 新疆野蘋果自然條件下次生代謝組分差異
研究表明,此次靶向代謝檢測分為四個大類,分別是羥基肉桂酰衍生物(圖1-A)、生物堿(圖1-B)、黃酮醇(圖1-C)和其它物質(圖1-D)。
羥基肉桂酰衍生物中對照株系香豆酸含量顯著高于抗蟲株系,含量分別為2.2×104和0.5×104μg/mL,差異倍數達到4.4。丁香酸含量高于抗蟲株系42.3%,未達到顯著差異。其中反式肉桂酸、咖啡酸、香草酸、阿魏酸含量變化不大;生物堿含量中,抗蟲株系普遍低于對照株系,其中苦豆堿含量最低,分別是3.9×103和3.1×103μg/mL,咖啡堿含量最高,達到1.1×106和9.5×105μg/mL,對照株系高于抗蟲株系16.9%。蘆竹堿和茶堿含量差異均不顯著。
黃酮醇含量測定中,各組分含量呈現不同差異變化趨勢,其中抗蟲株系的蕓香苷含量顯著高于對照株系,含量分別為9.8×106和4.4×106μg/mL,差異倍數為2.3。抗蟲株系的榭皮苷含量高于對照株系,分別達到2.5×107和2.1×107μg/mL;在檢測的其它組分中,差異均不顯著,其中綠原酸含量最高,苯甲酸含量最低。圖1
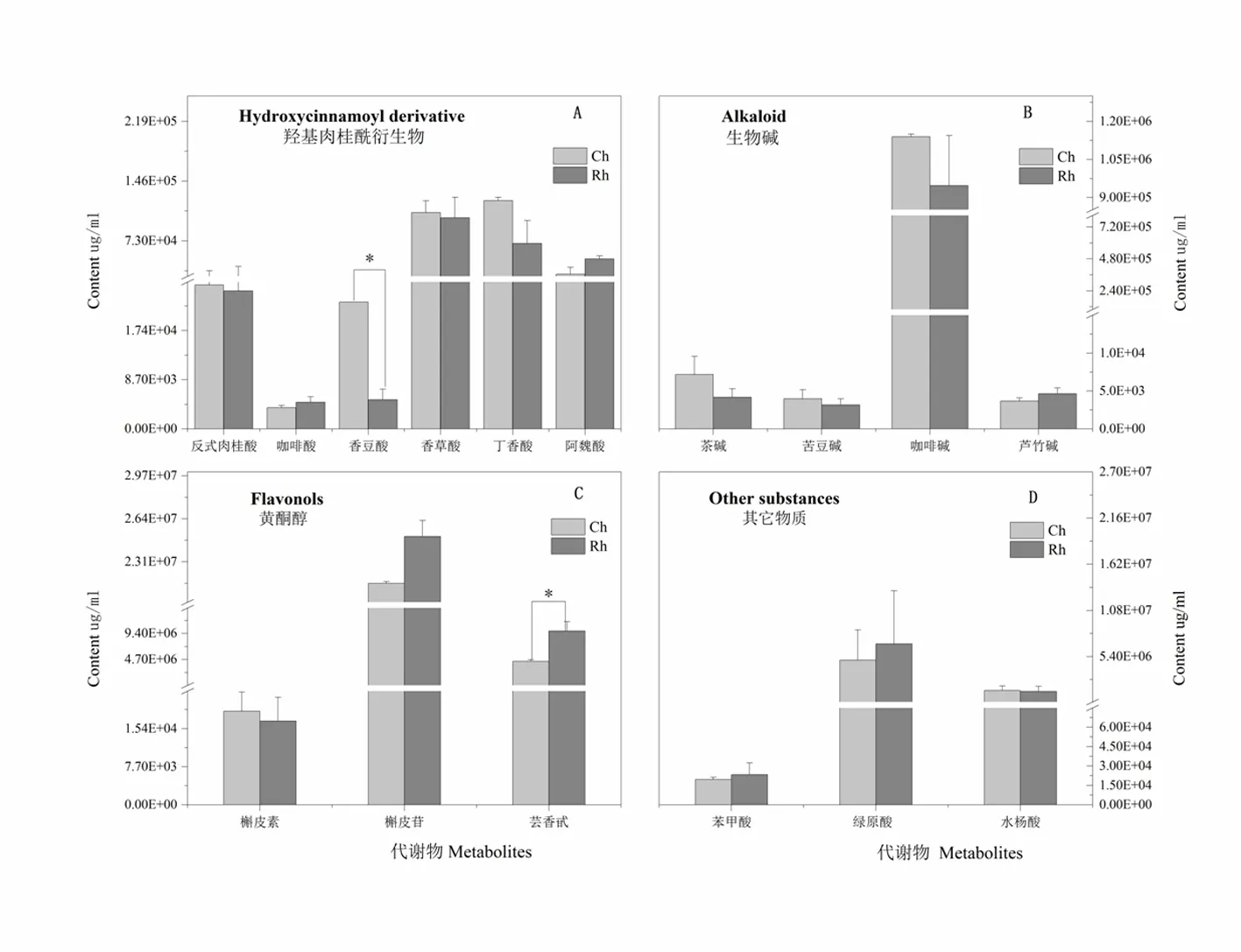
注:Ch:對照株系健康部位。Rh:抗蟲株系健康部位
Note:Ch: Healthy parts of control strains. Rh: Healthy parts of insect-resistant strains
圖1 新疆野蘋果自然條件下次生代謝組分差異
Fig. 1 Differences of the secondary metabolites in Xinjiang wild apple under normal conditions
2.2 新疆野蘋果蟲害侵染條件下次生代謝組分對蟲害脅迫的響應
研究表明,抗蟲株系與對照株系在被小吉丁蟲侵染處理后,羥基肉桂酰衍生物中的反式肉桂酸含量上升明顯,達到極顯著水平,含量分別為3.3×104和2.2×106μg/mL,差異倍數為6.8。同時抗蟲株系的咖啡酸含量極顯著高于對照株系,提升倍數為3.8。此外,阿魏酸含量也顯著上升,含量分別為4.5×104和5.3×104μg/mL(圖2-A)。由此可見,變化明顯的大多屬于羥基肉桂酰衍生物一大類,說明抗蟲植株在受到蟲害脅迫后,激發植物體自身調節變化,提高羥基肉桂酰衍生物次生代謝含量以抵御侵害;生物堿組分含量測定中,蟲害侵染處理后,各組分均沒有達到顯著差異水平,其中咖啡堿含量最高,對照株系高于抗蟲株系,含量為9.7×105和7.1×105μg/mL,差異倍數為1.4,苦豆堿含量最低(圖2-B)。
黃酮醇類組分中,對照株系榭皮素含量顯著高于抗蟲株系,達到44.3%,榭皮苷與蕓香苷含量在蟲害侵染前后變化差異不明顯,但榭皮苷含量在黃酮醇類組分中最高,分別為2.4×107和2.3×107μg/mL;在其它組分含量測定中,苯甲酸、綠原酸、水楊酸對蟲害脅迫響應明顯,均有不同程度的提高,其中水楊酸上升達到顯著水平。圖2
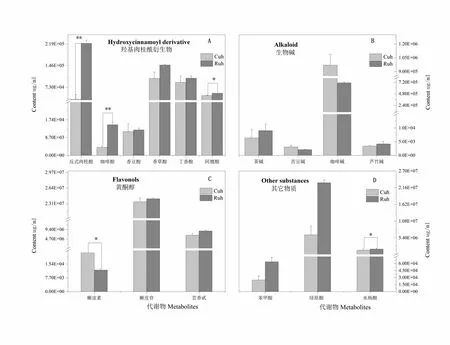
注:Cuh:對照株系侵染部位。Ruh:抗蟲株系侵染部位
Note:Cuh: Infection site of control strain. Ruh: Infection site of insect-resistant strains
圖2 新疆野蘋果蟲害侵染后次生代謝組分的響應
Fig. 2 Response of secondary metabolites after pest infestation in Xinjiang wild apple
3 討 論
植物的次生代謝產物非常豐富,根據成分,主要分為萜類、生物堿、苯丙烷類和相關苯基化合物四大類[20],植物受到蟲害侵染后,會產生誘導抗性,從而產生相關萜類化合物,組織防御體系[21],研究認為,在自然條件下,測定健康部位羥基肉桂酰衍生物、生物堿、黃酮醇及其它代謝物質變化差異不明顯。在蟲害脅迫條件下反式肉桂酸、咖啡酸、阿魏酸、綠原酸、水楊酸含量均顯著上升,說明這些代謝物對蟲害脅迫產生積極響應。此外,酚類物質、單寧、生物堿與蛋白質及酶作用,形成不易消化的復合物,影響昆蟲消化率及攝食量[22]。苯丙烷類途徑可生成反式肉桂酸、香豆酸、阿魏酸、芥子酸等中間產物,這些中間產物可進一步轉化為香豆素、綠原酸,也可形成反式香豆酰輔酶A酯,再通過多條途徑進一步轉化為木質素、黃酮、異黃酮、生物堿、苯甲酸酯糖苷等次生代謝產物,這些產物對昆蟲具有一定的毒害與防御作用[23]。興安落葉松受損后其葉內單寧含量呈顯著增加的趨勢,北美云杉(Piceasitchensis) 和多葉羽扇豆(Lupinuspolyphyllus) 在受到傷害后,創傷樹脂和生物堿的含量分別增加[24, 25]。與此同時許多酚酸對害蟲威懾非常有效,是許多具有傳遞抗性的復合分子前體[26]。通過代謝組與轉錄組聯合分析二化螟對水稻的響應機制認為,莽草酸、糖代謝、氨基酸、有機酸和萜類代謝等途徑共同參與了水稻抗蟲性調控過程[27]。這與研究結果相似,即新疆野蘋果受蟲害脅迫逆境條件下,通過調節自身免疫系統,造成植物“自身抗蟲”的表型。
植物在受到昆蟲危害后的防御反應是多維的動態過程,包含的層次與功能相對復雜[25],植物與昆蟲互作機理研究通常會使用高通量組學技術(轉錄組學、基因組學、代謝組學等)揭示其抗性產生機理[28]。通過轉錄組分析并鑒定候選基因,了解植物在生物脅迫和非生物脅迫過程中的應激反應[29-31]。蘋果小吉丁蟲是新疆野蘋果這一“起源種”的毀滅性蟲害,進一步通過轉錄組、代謝組和生理水平研究新疆野蘋果抗蘋果小吉丁蟲的生理生化響應機理、轉錄調控機制以及如何增強抗蘋果小吉丁蟲功能是接下來重點擬解決的關鍵科學問題。為有效保護、利用新疆野蘋果這一珍貴資源提供理論基礎。
4 結 論
新疆野蘋果在被蘋果小吉丁蟲侵染后反式肉桂酸含量極顯著提高,含量分別為3.3×104和2.2×106μg/mL,差異倍數為6.8。咖啡酸、綠原酸、槲皮素、阿魏酸組分含量顯著提高,且上升明顯的次生代謝物大多屬羥基肉桂酰衍生物一大類,抗蟲植株在受到蟲害脅迫后,此類代謝物質產生積極響應,激發植物體自身調節變化,提高羥基肉桂酰衍生物等次生代謝含量。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
國畫家(2022年2期)2022-04-13 09:07:46
四川文學(2021年4期)2021-07-22 07:11:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
絲綢之路(2014年9期)2015-01-22 04:24:46
山東女子學院學報(2014年6期)2014-03-01 02:24:55
兒童與健康(2011年4期)2011-04-12 00:00:00
