類烏齊牦牛ACTA1基因克隆、分子特性及差異表達(dá)分析
2019-07-18 05:32:50柴志欣王吉坤信金偉姬秋梅張成福鐘金城羅曉林
西南農(nóng)業(yè)學(xué)報(bào) 2019年6期
關(guān)鍵詞:分析
楊 勤,柴志欣,王 會,王吉坤,信金偉,姬秋梅,張成福,鐘金城*,羅曉林
(1.西南民族大學(xué) 青藏高原動物遺傳資源保護(hù)與利用教育部重點(diǎn)實(shí)驗(yàn)室,四川 成都 610041;2.西藏自治區(qū)農(nóng)牧科學(xué)院省部共建青稞和牦牛種質(zhì)資源與遺傳改良國家重點(diǎn)實(shí)驗(yàn)室,西藏 拉薩 850009;3.四川省草原科學(xué)研究院,四川 成都 611731)
【研究意義】隨著社會發(fā)展和人民生活水平的提高,人們的膳食結(jié)構(gòu)也在不斷發(fā)生變化,綠色食品日漸成為消費(fèi)熱點(diǎn)。其中牛肉營養(yǎng)豐富,含有人體所需的必需氨基酸,且具有高蛋白、低脂肪、低膽固醇等特點(diǎn),成為消費(fèi)者日常生活主要的肉質(zhì)來源之一;同時(shí),人們對“優(yōu)質(zhì)牛肉”、“高檔牛肉”的需求越來越高。牦牛是分布于青藏高原及其毗鄰的高山、亞高山地區(qū)的主要畜種,因主產(chǎn)區(qū)特有的生態(tài)環(huán)境和地理位置形成了許多地方品種或類群。類烏齊牦牛主要分布于西藏自治區(qū)東部昌都市類烏齊縣境內(nèi)海拔4500 m以上的草甸草原地區(qū),因氣候宜人,水草豐茂,俗稱西藏“小瑞士”,并以其肥膘體壯、肉質(zhì)鮮嫩、口感好、遺傳多樣性豐富等特點(diǎn),于2017年通過國家農(nóng)業(yè)資源委員會審定,成為國家優(yōu)良牦牛遺傳資源[1]。且因其形成歷史悠久,經(jīng)長期自群繁育,具有基本一致的外貌特征、繁殖性能和生產(chǎn)性能,在遺傳上是一個(gè)寶貴的基因庫,可作為培育牦牛新品種或品系的重要遺傳資源[2]。肌動蛋白(Actin)是真核細(xì)胞中含量最豐富的蛋白質(zhì)之一,主要分布于細(xì)胞質(zhì),參與肌小節(jié)和細(xì)胞骨架主要成分—微絲的組成,細(xì)胞骨架之所以能夠形成穩(wěn)定結(jié)構(gòu)及動態(tài)結(jié)構(gòu),是由于微絲特別是肌動蛋白微絲和微管的共同作用。微絲可與肌球蛋白相互作用產(chǎn)生滑動效應(yīng),是細(xì)胞運(yùn)動及肌肉收縮的生理基礎(chǔ)。肌動蛋白除參與肌球蛋白介導(dǎo)的肌肉收縮運(yùn)動外還參與細(xì)胞黏附、信號轉(zhuǎn)導(dǎo)、離子通道調(diào)控等相關(guān)調(diào)控機(jī)制[3]。α肌動蛋白1(actin alpha 1, ACTA1)是肌動蛋白的一類異構(gòu)形式,以收縮蛋白在骨骼肌的活動中發(fā)揮重要作用,是骨骼肌中一種收縮裝置的主要組成部分,并廣泛參與肌細(xì)絲組裝、骨骼肌纖維發(fā)育以及細(xì)胞和細(xì)胞器的運(yùn)動[4]。研究表明,ACTA1基因突變會導(dǎo)致多種肌肉疾病發(fā)生,如人類骨骼肌先天性肌病、橫紋肌疾病、桿狀體肌病與蓋肌病等[5-9]。【前人研究進(jìn)展】近年來,國內(nèi)外許多研究已對畜禽ACTA1基因進(jìn)行了研究報(bào)道。Sewry等[10]研究發(fā)現(xiàn),ACTA1基因突變與斑馬的肌無力相關(guān)。Venturini等[11]研究顯示,ACTA1基因多態(tài)性與肉雞的生產(chǎn)性能及胴體性狀顯著相關(guān)。Lee等[12]研究表明,12月齡韓牛背最長肌中ACTA1基因表達(dá)水平顯著高于27月齡韓牛,ACTA1基因可能是調(diào)控韓牛肉質(zhì)性狀的重要的基因。Shin等[13]研究發(fā)現(xiàn)ACTA1基因在韓牛低等級大理石花紋牛肉中表達(dá)量顯著高于高等級大理石花紋個(gè)體,ACTA1基因可能是影響肉質(zhì)性狀的重要基因。田萬年等[14-15]研究報(bào)道,延邊黃牛ACTA1基因與體長、體重、平均日增重和胸圍上顯著相關(guān)。張斌等[16]研究發(fā)現(xiàn),檳榔江水牛ACTA1基因存在2個(gè)SNPs多態(tài)位點(diǎn),多態(tài)類型與其體重、管圍等性狀存在關(guān)聯(lián)。以上研究表明,ACTA1基因是影響畜禽肉質(zhì)及生長性狀的主要候選基因,但有關(guān)牦牛ACTA1基因生物學(xué)功能及其表達(dá)規(guī)律的研究尚未見報(bào)道。【本研究切入點(diǎn)】本研究以類烏齊牦牛臀肌為試驗(yàn)材料,通過RT-PCR方法獲得牦牛ACTA1基因全長cDNA序列并進(jìn)行生物信息學(xué)分析;同時(shí)運(yùn)用RT-qPCR法檢測牦牛ACTA1基因在不同組織中mRNA水平的表達(dá)規(guī)律。【擬解決的關(guān)鍵問題】為進(jìn)一步研究該基因?qū)﹃笈<∪獍l(fā)育的調(diào)控作用及其與肉質(zhì)性狀的相關(guān)性等奠定理論依據(jù)。
1 材料與方法
1.1 樣品采集
于西藏類烏齊縣選取3頭4.5歲健康西藏類烏齊牦牛,采集心臟、臀肌、臀脂、脾臟、大腦、肺臟、肝臟和乳腺8種組織,DEPC水清洗干凈,錫箔紙包裝好迅速置于液氮中保存?zhèn)溆谩?/p>
1.2 主要試劑
Trizol RNA提取試劑盒(Thermo);瓊脂糖(Amresco);RNase-Free ddH2O;DNA純化試劑盒(TIANGEN);反轉(zhuǎn)錄試劑盒(Takara);DL2000 DNA Ladder(Takara);pMD19-T載體(TaKaRa);DH5α感受態(tài)細(xì)胞(TIANGEN)等。
1.3 總RNA的提取及cDNA的合成
稱取80~110 mg組織樣品于研缽中,邊研磨邊倒液氮,將研磨好的組織迅速轉(zhuǎn)移到含Trizol裂解液的離心管,提取總RNA。采用分光光度計(jì)測定RNA樣品的濃度及OD260nm/280nm值,并通過瓊脂糖凝膠電泳檢驗(yàn)RNA質(zhì)量,置于-80 ℃保存?zhèn)溆谩S梅崔D(zhuǎn)錄試劑盒PrimerScript?1stStrand cDNA Synthesis kit將提取的總RNA反轉(zhuǎn)錄為cDNA。
1.4 引物設(shè)計(jì)與合成
根據(jù)GenBank公布的普通牛ACTA1基因mRNA參考序列(XM_005226192)設(shè)計(jì)1對引物(表1)。

表1 ACTA1-PCR引物Table 1 ACTA1-PCR primer
同時(shí)根據(jù)克隆得到類烏齊牦牛ACTA1基因cDNA序列設(shè)計(jì)實(shí)時(shí)熒光定量qPCR引物,選用β-actin作為內(nèi)參基因。引物均由Thermo公司合成。
1.5 類烏齊牦牛ACTA1基因TA克隆與鑒定
以類烏齊牦牛臀肌組織cDNA為模板,以ACTA1-PCR 引物擴(kuò)增牦牛ACTA1基因CDS區(qū)。體系(25 μl):TaqGreen PCR Master Mix(2×, 1.25 mL)12.5 μl;上下游引物F/R各1 μl;cDNA 1μl,ddH2O 9.5 μl。PCR反應(yīng)程序:94 ℃預(yù)變性5 min;94 ℃變性30 s,57.2 ℃退火30 s,72 ℃延伸1 min,32個(gè)循環(huán);72 ℃終延伸5 min;4 ℃保存。PCR產(chǎn)物經(jīng)1 %瓊脂糖凝膠電泳檢測,利用DNA純化試劑盒回收PCR產(chǎn)物,4 ℃保存?zhèn)溆谩?/p>
將回收純化后的PCR產(chǎn)物與載體pMD19-T 16 ℃過夜連接,將連接載體轉(zhuǎn)化至DH5α感受態(tài)細(xì)胞,經(jīng)復(fù)蘇后,均勻涂布在制備好的Amp+-LB固體培養(yǎng)基平板上,37 ℃恒溫過夜培養(yǎng)10~12 h,挑取白色大小合適的單菌落,篩選陽性質(zhì)粒DNA送生工生物工程(上海)股份有限公司雙向測序。
1.6 qPCR分析
將3頭類烏齊牦牛的心臟、臀肌、臀脂、脾臟、大腦、肺臟、肝臟和乳腺共24種組織的RNA稀釋成100 ng·μl-1,分別取5 μl RNA 進(jìn)行反轉(zhuǎn)錄反應(yīng),得到的cDNA 產(chǎn)物采用實(shí)時(shí)熒光定量 PCR(RT-qPCR) 檢測ACTA1基因和β-actin基因的表達(dá)。反應(yīng)體系(10 μl) : SYBR premix Dimer Eraser(2×)5 μl,ddH2O 3.2 μl,的上下游引物(10 μmol·L-1)各 0.4 μl,cDNA 1 μl。qPCR 反應(yīng)條件:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,39個(gè)循環(huán)。
1.7 生物信息學(xué)分析
對類烏齊牦牛ACTA1基因CDS區(qū)進(jìn)行序列分析和蛋白結(jié)構(gòu)、功能預(yù)測:利用NCBI-BLAST對所測序列進(jìn)行比對校正;ORF finder程序分析ACTA1基因開放閱讀框并獲得氨基酸序列;利用DNAStar進(jìn)行物種間序列的同源性分析;利用MEGA7.0軟件構(gòu)建系統(tǒng)進(jìn)化樹;利用在線工具Protparam(https://web.expasy.org/protparam/)和Protscale(https://web.expasy.org/protscale/)分別分析ACTA1氨基酸的理化性質(zhì)和親水性;利用在線軟件預(yù)測Netphos(http://www.cbs.dtu.dk/services/NetPhos/)ACTA1蛋白潛在磷酸化位點(diǎn);利用在線軟件NetOGlyc(http://www.cbs.dtu.dk/services/NetOGlyc)和NetNGlyc(http://www.cbs.dtu.dk/services/NetNGlyc/)進(jìn)行潛在O、N糖基化位點(diǎn)預(yù)測;利用在線軟件TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)預(yù)測ACTA1蛋白跨膜結(jié)構(gòu)域和跨膜方向;利用在線軟件Sopma(https://npsa-prabi.ibcp.fr/cgibin/secpred sopma.pl)預(yù)測ACTA1蛋白二級結(jié)構(gòu);利用NCBI工具Conserved Domain Search Service (CD Search)預(yù)測ACTA1蛋白保守結(jié)構(gòu)域;利用在線軟件SWISSMODEL(https://www.swissmodel.expasy.org/)預(yù)測ACTA1蛋白質(zhì)三級結(jié)構(gòu)。
1.8 qPCR數(shù)據(jù)搜集與分析
RT-qPCR結(jié)果采用 2-△△CT法分別計(jì)算每個(gè)樣品ACTA1和 β-actin基因表達(dá)量相對值,用軟件GraphPad Prism 5繪制ACTA1在不同組織中 mRNA 表達(dá)量的柱狀圖,采用SPSS22.0軟件分析各組織間差異顯著性。
2 結(jié)果與分析
2.1 類烏齊牦牛ACTA1基因RT-PCR
由圖1所示,以類烏齊牦牛臀肌總RNA為模板,反轉(zhuǎn)錄獲得cDNA,PCR擴(kuò)增序列長1239 bp與目的片段大小一致。
2.2 ACTA1基因的序列分析
如圖2可知,經(jīng)ORF finder分析顯示類烏齊牦牛ACTA1基因CDS區(qū)全長1134 bp,起始密碼子為ATG,終止密碼子為TAG,共編碼377個(gè)氨基酸。

圖1 ACTA1基因PCR擴(kuò)增Fig.1 PCR amplification products of ACTA1 gene
2.3 同源性及進(jìn)化樹分析
從NCBI數(shù)據(jù)庫搜集12個(gè)物種ACTA1基因編碼區(qū)mRNA序列和對應(yīng)氨基酸序列,利用MegAlign軟件對類烏齊牦牛、黃牛、水牛等13個(gè)物種ACTA1基因編碼區(qū)核苷酸序列及氨基酸序列進(jìn)行比對,結(jié)果表明ACTA1基因編碼區(qū)序列在哺乳動物間高度保守(表2)。利用NJ法構(gòu)建系統(tǒng)進(jìn)化樹,結(jié)果顯示,類烏齊牦牛與水牛、黃牛、綿羊及山羊遺傳距離較近,而與熱帶爪蟾較遠(yuǎn)(圖3)。
2.4 氨基酸序列理化特性預(yù)測與分析
運(yùn)用ExPASy中 ProtParam程序?qū)CTA1蛋白氨基酸序列的理化性質(zhì)進(jìn)行預(yù)測,ACTA1蛋白分子式為C1860H2917N495O568S23,理論分子質(zhì)量為42.03 ku,理論等電點(diǎn)(PI)為5.23,為酸性蛋白,共有50個(gè)負(fù)電荷氨基酸殘基(Asp + Glu)和37個(gè)正電荷氨基酸殘基(Arg + Lys)。ACTA1蛋白含有20種標(biāo)準(zhǔn)氨基酸,由氨基酸組成可知(表3),Ala(A)和Ile(I)分別占比7.7 %,含量最多,Trp(W)含量最低,僅占1.1%。ACTA1蛋白的不穩(wěn)定系數(shù)為36.14,低于閾值40,屬穩(wěn)定蛋白。估測其在體外哺乳動物網(wǎng)織紅細(xì)胞內(nèi)的半衰期為30 h,脂肪系數(shù)為80.74,總平均親水系數(shù)為(GRAVY)為-0.246,為親水性蛋白。
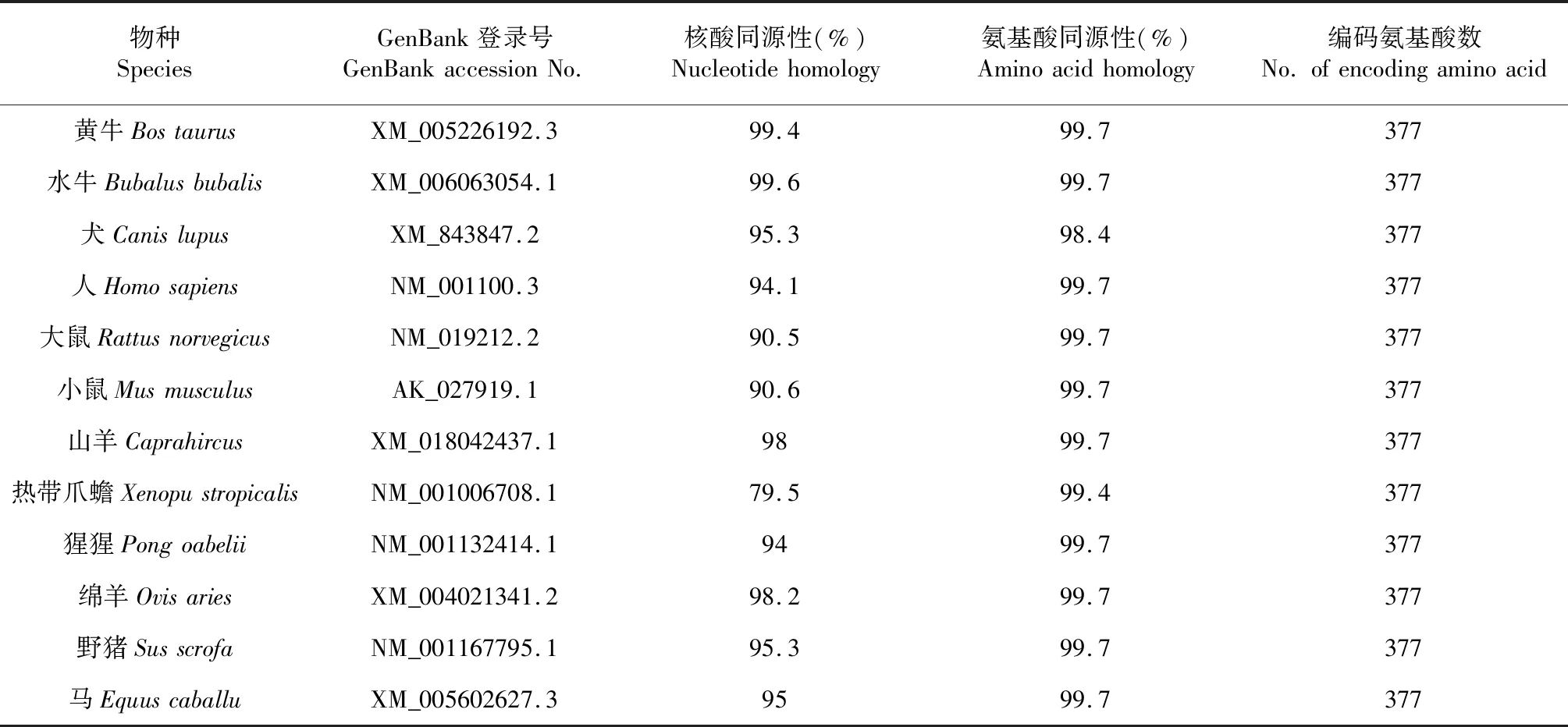
表2 類烏齊牦牛與其他物種ACTA1基因編碼區(qū)同源性比較Table 2 Homology comparison of ACTA1 gene CDS between Leiwuqi yak and other species
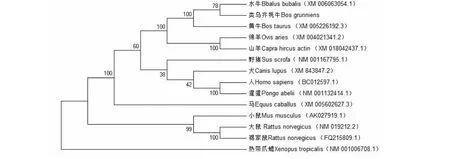
圖3 類烏齊牦牛與其他物種的ACTA1基因系統(tǒng)進(jìn)化樹分析Fig.3 Phylogenetic relationship of ACTA1 genes based on Leiwuqi yak and other species
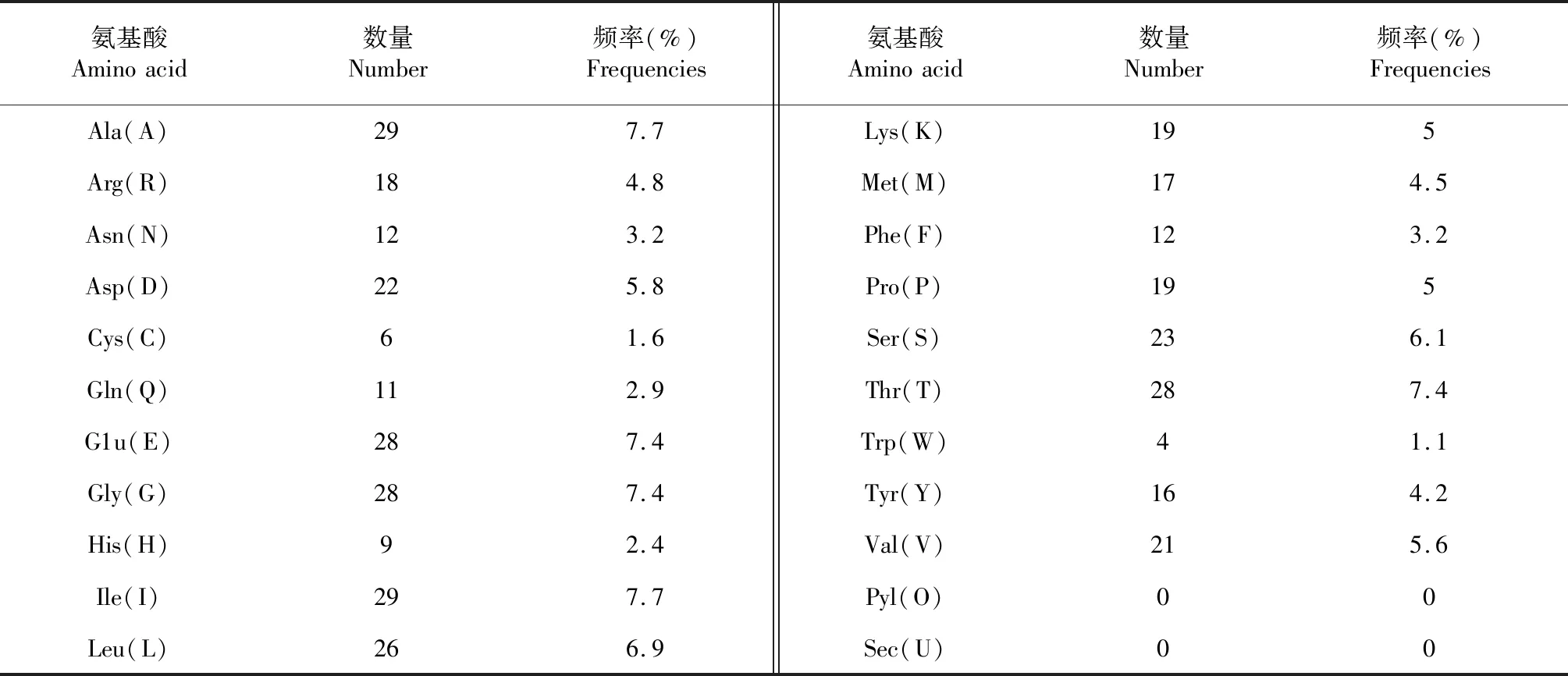
表3 類烏齊牦牛ACTA1基因編碼蛋白氨基酸組成Table 3 Composition of amino acid coded by Leiwuqi yak ACTA1 gene
2.5 ACTA1蛋白親疏水性預(yù)測與分析
根據(jù)蛋白質(zhì)親水性/疏水性分析原理,利用ExPASy在線工具ProtScale程序?qū)CTA1基因編碼產(chǎn)物進(jìn)行親/疏水性分析。由圖4可知,縱坐標(biāo)0值以上為疏水區(qū),其分值越高,疏水性越強(qiáng);0值以下為親水區(qū)。整個(gè)多肽鏈包含2個(gè)較明顯疏水區(qū)和2個(gè)較明顯親水區(qū),第1疏水區(qū)為第137~142位氨基酸,第2疏水區(qū)為第344~347位氨基酸;第1親水區(qū)為第115~119位氨基酸,第2親水區(qū)為362~366位氨基酸。該氨基酸多肽鏈的第140位Ala(A)分值最高(2.200),疏水性最強(qiáng);第117位Asn(N)分值最低(-2.300),親水性最強(qiáng)。分析數(shù)據(jù)顯示,該氨基酸序列內(nèi)超過半數(shù)為親水性殘基,整條多肽鏈表現(xiàn)為親水性。
2.6 ACTA1蛋白結(jié)構(gòu)功能預(yù)測與分析
2.6.1 ACTA1蛋白結(jié)構(gòu)預(yù)測與分析 使用在線軟件SOPMA預(yù)測類烏齊牦牛ACTA1的蛋白質(zhì)二級結(jié)構(gòu),發(fā)現(xiàn)該蛋白由無規(guī)則卷曲(Random coil)、α-螺旋(Alpha helix)和β-轉(zhuǎn)角(Beta turn)以及延伸鏈(Extended strand)構(gòu)成(表4),為其三級結(jié)構(gòu)預(yù)測提供參考。利用SWISS-MODEL在線軟件基于PDB數(shù)據(jù)庫對類烏齊牦牛ACTA1蛋白進(jìn)行同源建模,獲得ACTA1蛋白三級結(jié)構(gòu)模型(圖5)。由三級結(jié)構(gòu)可知,α-螺旋占主導(dǎo)地位,與二級結(jié)構(gòu)預(yù)測結(jié)果一致。
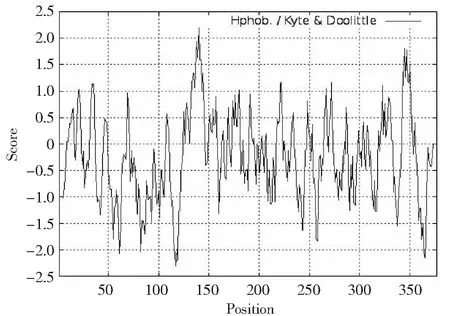
正值表示疏水,負(fù)值表示親水窗口大小為n=9Positive value indicates hydrophobicity, negative value indicates hydrophilicity; The window size is n=9圖4 類烏齊牦牛ACTA1基因編碼蛋白疏水性預(yù)測Fig.4 Hydrophobicity prediction of Leiwuqi yak ACTA1 gene encoding protein

表4 類烏齊牦牛ACTA1基因編碼蛋白二級結(jié)構(gòu)預(yù)測Table 4 Secondary structure prediction of Leiwuqi yak ACTA1 gene encoding protein
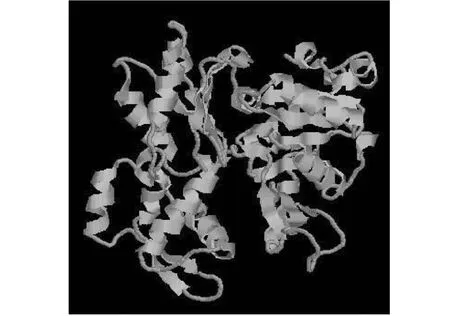
圖5 類烏齊牦牛ACTA1基因編碼蛋白三級結(jié)構(gòu)預(yù)測Fig.5 Tertiary structure prediction of Leiwuqi yak ACTA1 gene encoding protein

圖6 類烏齊牦牛ACTA1基因編碼蛋白跨膜結(jié)構(gòu)域分析Fig.6 Analysis of transmembrane domain of Leiwuqi yak ACTA1 gene encoding protein
2.6.2 跨膜區(qū)結(jié)構(gòu)預(yù)測與分析 利用軟件TMHMM2.0軟件預(yù)測ACTA1蛋白跨膜結(jié)構(gòu)域。由圖6可知,該蛋白含有2個(gè)跨膜螺旋結(jié)構(gòu),即為二次跨膜蛋白,在第131~148、339~357位氨基酸處分別存在1個(gè)跨膜螺旋,該蛋白所有氨基酸均為胞內(nèi)區(qū),屬胞內(nèi)蛋白,這是因?yàn)榧拥鞍字饕嬖谟诩∪饧?xì)胞細(xì)胞質(zhì)內(nèi),與肌球蛋白結(jié)合參與肌小節(jié)收縮。
2.6.3 信號肽預(yù)測與分析 如圖7所示,在線工具SignalP4.1Server預(yù)測ACTA1蛋白信號肽,表明ACTA1蛋白無信號肽序列,說明該蛋白為非分泌型蛋白,進(jìn)一步說明該蛋白為胞內(nèi)蛋白,主要在骨骼肌細(xì)胞內(nèi)發(fā)揮作用。
2.6.4 保守結(jié)構(gòu)域預(yù)測與分析 利用NCBI工具Conserved Domain Search Service (CD Search)預(yù)測ACTA1蛋白保守結(jié)構(gòu)域。如圖8所示,ACTIN蛋白家族為糖激酶/HSP70/肌動蛋白超家族成員之一,其中ACTA1蛋白多肽鏈第7~377位氨基酸為肌動蛋白保守結(jié)構(gòu)域,第10~183位為肌動蛋白NBD(Nucleotide binding site)區(qū)域,肌動蛋白在生物進(jìn)化過程中高度保守。
2.6.5 磷酸化位點(diǎn)預(yù)測與分析 利用NetPhos3.1 Server預(yù)測潛在磷酸化位點(diǎn),由圖9可知,閾值為0.5時(shí),ACTA1蛋白存在38個(gè)潛在的磷酸化位點(diǎn),其中包括16處絲氨酸(Serine)磷酸化位點(diǎn)、13處蘇氨酸(Thronine)磷酸化位點(diǎn)和9處酪氨酸(Tyrosine)磷酸化位點(diǎn)。
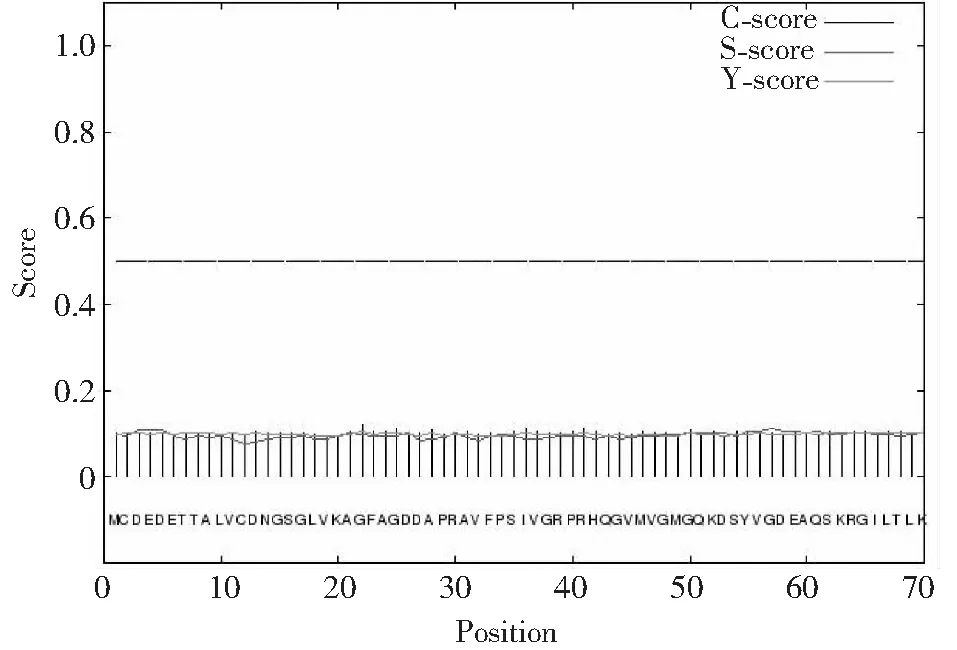
圖7 類烏齊牦牛ACTA1基因編碼蛋白信號肽預(yù)測Fig.7 Signal peptide prediction of Leiwuqi yak ACTA1 gene encoding protein

圖8 類烏齊牦牛ACTA1基因編碼蛋白保守位點(diǎn)預(yù)測Fig.8 Conserved sites prediction of Leiwuqi yak ACTA1 gene encoding protein
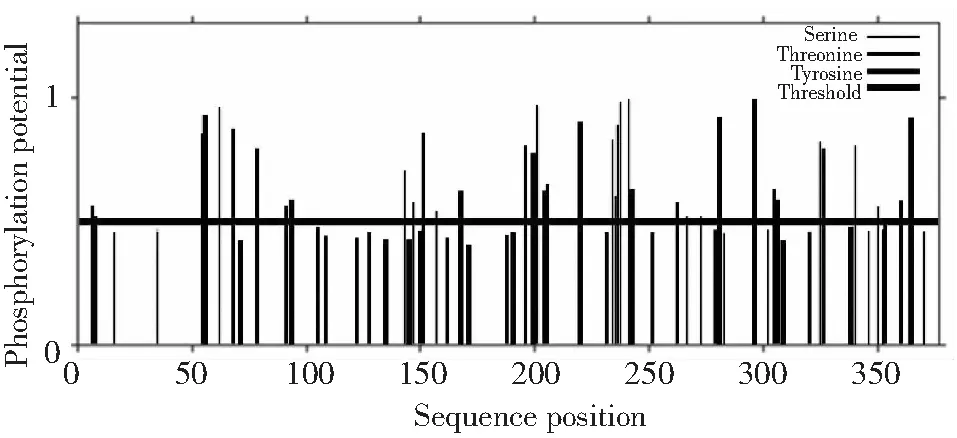
圖9 類烏齊牦牛ACTA1基因編碼蛋白磷酸化位點(diǎn)預(yù)測Fig.9 Phosphorylation sites prediction of Leiwuqi yak ACTA1 gene encoding protein
2.6.6 O/N-糖基化位點(diǎn)預(yù)測與分析 利用軟件NetOGlyc4.0 Server和NetNGlyc1.0 Server進(jìn)行潛在O、N糖基化位點(diǎn)預(yù)測。分析結(jié)果顯示,ACTA1蛋白僅存在1處潛在O-糖基化修飾位點(diǎn),其概率為0.50365;而在第14位對應(yīng)天冬酰胺殘基的酰胺氮原子存在1處潛在N-糖基化修飾位點(diǎn),其概率為0.6867(圖10)。
2.7 類烏齊牦牛ACTA1 mRNA組織表達(dá)
實(shí)時(shí)熒光定量PCR結(jié)果顯示,ACTA1基因在檢測的3頭類烏齊牦牛臀肌、大腦、心臟等8種組織中均有表達(dá);利用Spass 22.0進(jìn)行單因素方差分析,P<0.05為差異顯著,P<0.01為差異極顯著,結(jié)果發(fā)現(xiàn)ACTA1mRNA在臀肌中表達(dá)水平極顯著高于大腦和心臟(P<0.01),大腦和心臟ACTA1mRNA表達(dá)差異不顯著(P>0.05),而在肝臟及臀脂中表達(dá)量極低(圖11)。
3 討 論
目前,對人類ACTA1基因的研究主要集中于疾病方面,如基因突變與人骨骼先天性疾病、橫紋肌疾病相關(guān)性等,而畜禽方面則主要分析ACTA1基因突變與肌無力、肉質(zhì)性狀及生長性狀相關(guān)性等方面。本試驗(yàn)成功克隆類烏齊牦牛ACTA1基因CDS區(qū),并通過生物信息學(xué)分析詳細(xì)闡釋ACTA1蛋白理化性質(zhì)、結(jié)構(gòu)功能。類烏齊牦牛ACTA1基因CDS區(qū)全長1134 bp,共編碼377個(gè)氨基酸。

圖10 類烏齊牦牛ACTA1基因編碼蛋白N-糖基化位點(diǎn)分析Fig.10 N-glycosylation site analysis of Leiwuqi yak ACTA1 gene encoding protein

**表示差異極顯著(P<0.01)**mean significant difference at 0.01 level圖11 類烏齊牦牛ACTA1基因在不同組織中的表達(dá)量(內(nèi)參基因: β-actin; n= 3)Fig.11 Expression of Leiwuqi yak ACTA1 gene in various tissues
類烏齊牦牛肉質(zhì)鮮美,口味獨(dú)特,可能與肉中氨基酸含量顯著相關(guān),羅毅皓等[16]研究報(bào)道,鮮味氨基酸天門冬氨酸(Asp)和谷氨酸(Glu)決定了牦牛肉鮮味優(yōu)于黃牛肉,而本研究顯示ACTA1蛋白天門冬氨酸與谷氨酸含量分別高達(dá)5.8 %、7.4 %,谷氨酸含量越高,肉品質(zhì)越好,且易消化吸收,屬于優(yōu)質(zhì)牛肉。膠原是人體結(jié)締組織的主要組成,肉質(zhì)中精氨酸(Arg)和丙氨酸(Ala)含量高有利于人體膠原的合成,特別是對兒童生長發(fā)育有重要促進(jìn)作用,本研究發(fā)現(xiàn)類烏齊牦牛肉中精氨酸和丙氨酸含量分別高達(dá)5.8 %、7.7 %,有利于膠原的合成,老人、兒童皆宜食用。另親水性/疏水性分析顯示,類烏齊牦牛ACTA1蛋白多肽鏈所含疏水性氨基酸多于親水性氨基酸,但多肽鏈經(jīng)過二、三級結(jié)構(gòu)在空間上的盤繞、折疊后,表現(xiàn)為較強(qiáng)親水性,易與水分子結(jié)合,同時(shí)其總平均親水系數(shù)(GRAVY)預(yù)測也表明該蛋白屬親水性蛋白,因此,ACTA1蛋白在其執(zhí)行功能上表現(xiàn)為親水性蛋白。跨膜區(qū)預(yù)測顯示,ACTA1蛋白所有氨基酸均為胞內(nèi)區(qū),存在兩個(gè)較明顯的跨膜螺旋區(qū)域;類烏齊牦牛ACTA1蛋白無信號肽,屬非分泌蛋白,表明該蛋白是由細(xì)胞質(zhì)內(nèi)游離的核糖體合成,不經(jīng)過內(nèi)質(zhì)網(wǎng)、高爾基體的加工和細(xì)胞膜的胞吐,只在細(xì)胞內(nèi)產(chǎn)生影響,為胞內(nèi)蛋白[17],而胞內(nèi)蛋白一般為親水性蛋白[18],這與預(yù)測結(jié)果一致。且ACTA1蛋白在肌肉收縮及興奮偶聯(lián)過程中,促進(jìn)肌動蛋白微絲與橫橋間的相互作用,產(chǎn)生收縮,推測該蛋白跨膜螺旋區(qū)域可能與肌動蛋白微絲與橫橋間存在信號轉(zhuǎn)導(dǎo)有關(guān)[20]。ACTA1蛋白二級和三級結(jié)構(gòu)顯示,多肽鏈在空間上的螺旋、折疊均以α-螺旋占主導(dǎo),有助于肌纖維肥大,從而有助于動物軀體肌肉塊增大增粗[21]。
序列同源性和系統(tǒng)進(jìn)化樹分析在一定程度上反應(yīng)出物種間的親緣關(guān)系[22]。本研究發(fā)現(xiàn),類烏齊牦牛ACTA1基因與水牛的親緣關(guān)系最近,CDS區(qū)序列同源性可達(dá)99.6 %,其次是黃牛(99.4 %)和綿羊(98.6 %),與馬、大鼠,小鼠等同源性較低,該結(jié)果與NJ法構(gòu)建的ACTA1基因系統(tǒng)進(jìn)化樹分析結(jié)果基本吻合。雖然熱帶爪蟾與類烏齊牦牛的基因序列同源性僅79.5 %,但其編碼的氨基酸序列同源性卻高達(dá)99.4 %,這說明在各物種的ACTA1基因氨基酸在長期生物進(jìn)化中具有較強(qiáng)的保守性[23];同時(shí)也反映了ACTA1基因編碼產(chǎn)物結(jié)構(gòu)和功能的穩(wěn)定性遺傳。
在不同組織器官、不同發(fā)育階段肌動蛋白mRNA相對表達(dá)量均不同。α肌動蛋白2(ACTA2)蛋白是ACTA1蛋白的異構(gòu)形式,黃京書等[24]研究了骨骼肌ACTA2蛋白在豬的不同發(fā)育階段呈差異表達(dá),且ACTA2mRNA表達(dá)量隨日齡增加而降低。田萬年等[14]對延邊黃牛8個(gè)組織器官表達(dá)水平的研究表明,ACTA1mRNA在不同組織器官呈差異表達(dá),且主要集中在心臟和背最長肌,在其他組織器官不表達(dá)。本研究分析了類烏齊牦牛8個(gè)不同組織器官之間的差異表達(dá),結(jié)果發(fā)現(xiàn)ACTA1基因在8個(gè)組織中均有表達(dá),且呈現(xiàn)一定差異性,具體表現(xiàn)為臀肌>大腦>心臟>肺臟>脾臟>乳腺>肝臟>臀脂。結(jié)果與田萬年等[14]報(bào)道的延邊黃牛ACTA1基因僅在心臟和背最長肌中表達(dá)的結(jié)論存在差異,可能類烏齊牦牛常年生活在平均海拔4000 m的草甸草原地區(qū),是世界上生活在海拔最高處的(除人類外)哺乳動物,且善走陡坡險(xiǎn)路、雪山沼澤,能游渡江河激流,其骨骼肌為適應(yīng)代謝及高強(qiáng)度運(yùn)動需求,臀肌等骨骼肌肌纖維含量非常豐富且發(fā)達(dá)。已有研究證實(shí)[25],在小鼠胚胎發(fā)育過程中,低氧環(huán)境可誘導(dǎo)其胚胎干細(xì)胞分化成心肌細(xì)胞及神經(jīng)纖維細(xì)胞,以適應(yīng)低氧環(huán)境;因此,本研究中類烏齊牦牛ACTA1基因在心臟和大腦組織中均有表達(dá),且表達(dá)量僅次于臀肌,是其對高原低氧環(huán)境具有很好適應(yīng)性的另一有力證明。
4 結(jié) 論
成功克隆了類烏齊牦牛ACTA1基因編碼區(qū),系統(tǒng)闡述了ACTA1蛋白理化性質(zhì)、結(jié)構(gòu)功能,以及其在長期生物進(jìn)化過程中高度保守;通過組織表達(dá)闡釋了ACTA1基因編碼蛋白是骨骼肌肌纖維的重要組成部分。為進(jìn)一步研究牦牛ACTA1基因在調(diào)控牦牛肌肉發(fā)育及其分子機(jī)制等方面奠定基礎(chǔ),也為西藏類烏齊牦牛相關(guān)研究提供數(shù)據(jù)資料。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財(cái)經(jīng)界(學(xué)術(shù)版)(2015年20期)2015-12-23 09:20:13
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
