重組人PLCζ蛋白在昆蟲細胞/桿狀病毒表達系統內的表達、純化及活性測定
2019-07-10 08:38:56陳鑫胡玥玥徐鴻毅王曉燕鄧鍇
生物工程學報 2019年6期
陳鑫,胡玥玥,徐鴻毅,王曉燕,鄧鍇
?
重組人PLCζ蛋白在昆蟲細胞/桿狀病毒表達系統內的表達、純化及活性測定
陳鑫1,胡玥玥1,徐鴻毅1,王曉燕2,鄧鍇1
1 湖北醫藥學院附屬人民醫院 生殖醫學中心,湖北 十堰 442000 2 湖北醫藥學院 基礎醫學院,湖北 十堰 442000
PLCζ是PLC家族的一種新型同工酶,在哺乳動物卵母細胞激活中起著極其重要的作用。近年來,體外大量表達和純化有活性的PLCζ蛋白用于結構生物學研究一直未能獲得成功。本研究首次在桿狀病毒表達系統中表達和純化重組人PLCζ蛋白,首先將人PLCζ基因克隆至pFastBac-HTA質粒構建重組載體,轉化DH10Bac發生位點特異性轉座,經抗性和藍白斑篩選,獲得重組穿梭質粒Bacmid-PLCζ;在脂質體介導下將穿梭質粒轉染Sf9昆蟲細胞產生重組病毒,擴增病毒感染Sf9昆蟲細胞進行蛋白表達;利用Ni2+親和柱及分子篩來純化蛋白,并通過考馬斯亮藍染色、Western blotting及飛行時間質譜對蛋白進行鑒定,并進行酶活性測定。結果顯示重組蛋白在Sf9昆蟲細胞感染桿狀病毒后72 h達到峰值并以分泌形式表達在細胞培養基中,Ni2+親和柱及分子篩純化后的重組蛋白經Western blotting及電離飛行時間質譜鑒定為PLCζ蛋白,酶活性可達326.8 U/mL。該實驗結果為重組人PLCζ蛋白大規模生產和生物醫學應用研究提供了可參考利用的技術。
PLCζ蛋白,昆蟲細胞,桿狀病毒表達系統,表達,純化,鑒定
磷脂酶C-zeta (Phospholipase C-zeta,PLCζ)是磷脂酶C家族的一種新型同工酶,是哺乳動物精子激活卵母細胞重要的生理性刺激因子[1]。在哺乳動物受精過程中,PLCζ蛋白作為“精子因子”被釋放入卵母細胞中,水解磷脂酰肌醇-4,5-二磷酸脂 (Phosphatidylinositol 4,5-bisphosphate,PIP2)產生第二信使1,4,5-三磷酸肌醇(Inositol-1,4,5- triphosphate,IP3),誘導卵母細胞內Ca2+周期性增加,從而使處于停滯期的卵母細胞恢復第二次減數分裂并啟動胚胎發育[2-3]。精子中PLCζ蛋白水平降低或缺失與卵母細胞激活缺陷型男性不育密切相關,人工輔助卵母細胞激活是解決此類男性不育的重要方法[4-5]。目前常用的人工輔助卵母細胞激活方法是機械刺激及化學激活,考慮到這些外界操作影響卵母細胞質量以及化學物質對胚胎的潛在危害,人工輔助卵母細胞激活技術未能廣泛開展[6-7]。PLCζ蛋白在輔助卵母細胞激活中有著較大的應用潛能,使用純化的具有活性的重組人PLCζ蛋白可能是一種更安全有效的治療卵母細胞激活缺陷型男性不育癥的潛在方法[8-10]。
體外表達和純化有活性的重組人PLCζ蛋白用以生物學應用的研究一直備受關注,國內外學者對重組人PLCζ蛋白的體外表達進行了廣泛的探索,Nomikos等嘗試用含組氨酸標簽的質粒載體通過大腸桿菌原核表達系統純化重組蛋白,發現原核系統表達量極低,將這些蛋白顯微注射到小鼠卵母細胞中時不能引起細胞內Ca2+振蕩啟動胚胎發育[11]。與大腸桿菌等原核表達系統相比,桿狀病毒表達系統在增加重組蛋白的可溶性、蛋白質翻譯后加工修飾方面具有天然優勢,能更好地保持蛋白的生物活性[12]。為了大量表達和純化出有活性的重組人PLCζ蛋白,進一步探索精卵融合時其蛋白與受體之間相互作用以及其作為輔助卵母細胞激活生物制劑的可行性,本研究從質粒構建開始,利用昆蟲桿狀病毒表達系統成功地表達并純化出有活性的重組人PLCζ蛋白,為接下來對PLCζ蛋白質的結構-功能關系及PLCζ生物醫學應用研究奠定基礎。
1 材料與方法
1.1 主要實驗材料
草地貪夜蛾Sf9昆蟲細胞、桿狀病毒Bac-to-Bac表達系統(重組桿狀病毒轉移載體pFastBac HTA、DH10Bac感受態大腸桿菌、Cellfectin Ⅱ試劑)、SF-900II SFM培養基、Grace’s Insect Medium、ECL顯色試劑盒等購自Invitrogen公司;DNA 高保真聚合酶、限制性內切酶Ⅰ、dⅢ等購自寶生物工程(大連) 有限公司;對硝基磷酸膽堿(p-nitrophehylphosphoryl choline,p-NPPC)、十二烷基硫酸鈉(SDS)、考馬斯亮藍R-250、甲基纖維素等購自Sigma公司;His GraviTrap親和層析柱購自GE Healthcare公司;兔源抗人PLCζ蛋白單克隆抗體和HRP標記的山羊抗兔IgG抗體均購自Abcam公司;PureLink HiPure Plasmid Miniprep kit購自QIAGEN公司;其他試劑均為國產分析純級。
1.2 引物的設計與轉移載體構建
檢索GenBank數據庫獲取人PLCζ基因(登錄號:NM_033123) 的mRNA序列,由南京金斯瑞生物科技有限公司合成;合成的全長人PLCζ編碼基因為1 841bp,其中5′端插入Ⅰ酶切位點,3′端插入dⅢ酶切位點。將合成的PLCζ目的片段克隆至經Ⅰ、dⅢ雙酶切后的pFastBac HTA載體中,構建重組轉移質粒pFastBac-PLCζ。PLCζ特異性引物參照Primer Premier 5擴增全長設計,引物由南京金斯瑞生物科技有限公司合成,引物序列見表1。
1.3 重組桿狀病毒穿梭載體的制備
按照Invitrogen公司的Bac-to-Bac Baculovirus Expression System使用說明將pFastBac-PLCζ構建體轉化DH10Bac感受態大腸桿菌細胞,用未插入外源基因的pFastBac HTA質粒轉化DH10Bac感受態細胞作為陰性對照[13]。取50 μL菌液涂布含有50 μg/mL卡那霉素、10 μg/mL四環素、 7 μg/mL慶大霉素、100 μg/mL X-gal和40 μg/mL IPTG的LB瓊脂平板,置于37 ℃培養箱內培養,48 h后進行藍白斑篩選。隨機挑取白色單克隆菌落接種于LB液體培養基37 ℃過夜培養。使用PureLink Hipure Plasmid Miniprep kit提取重組桿粒并用PLCζ特異性引物進行PCR擴增,PCR產物用1%瓊脂糖凝膠電泳檢測,鑒定正確的重組桿粒命名為Bacmid-PLCζ。

表1 載體構建及鑒定所用的引物
1.4 重組桿狀病毒的制備與鑒定
按照Invitrogen公司的CellfectinⅡ陽離子脂質體轉染試劑操作說明轉染對數生長期Sf9昆蟲細胞,5 h后更換新鮮的完全培養基,置于27 ℃培養箱培養72?96 h。當細胞出現明顯病變時離心收集上清液即為P1代重組病毒,將P1代重組桿狀病毒按照感染復數(MOI) 為0.1接種對數期生長的Sf9細胞進行重組病毒的增殖與傳代,得到P2代重組桿狀病毒。按照相同的方法將病毒傳至P3代,采用改良的病毒噬斑法即來測定桿狀病毒儲液的滴度[14],即用甲基纖維素代替傳統的瓊脂糖覆蓋病毒感染后的細胞,之后進行病毒噬斑的檢測。
1.5 重組PLCζ蛋白的表達、純化及鑒定
將P3代重組桿狀病毒以MOI=3的比例感染處于對數生長中期、密度為2×106/mL的Sf9昆蟲細胞,100 r/min、27 ℃培養96 h。收獲細胞混合物,3 800 r/min離心40 min,上清液經0.22 μm濾膜過濾后經切向流超濾膜包VIVAFLOW 200濃縮至50 mL,采用Ni-NTA親和層析柱純化上清液,以含250 mmol/L咪唑的變性緩沖液進行洗脫,收集洗脫液后經分子篩進一步純化。純化的PLCζ蛋白樣品經煮沸變性處理后用兩塊8%分離膠進行SDS-PAGE。其中一塊凝膠用考馬斯亮藍快速染色液染色,隨后用脫色液進行脫色,以鑒定目的蛋白的分子質量,同時切膠后經基質輔助激光解析電離-飛行時間質譜儀進行質譜鑒定。另一塊凝膠經半干法轉印PVDF膜,以兔源的抗重組人PLCζ蛋白抗體作為一抗,以HRP標記的山羊抗兔作為二抗進行常規Western blotting鑒定。
1.6 重組人PLCζ蛋白的活性檢測
PLC可水解p-NPPC生成黃色物質對硝基苯酚,對硝基苯酚在410 nm處有最大吸光度值,故可以通過酶標儀測定反應前后在410 nm處吸光度值的變化,利用標準曲線法定量計算重組人PLCζ蛋白的活性[15]。反應混合物中含0.25 mol/L Tris-HCl (pH 7.2) 緩沖液,1 mmol/L的ZnCl2,60%的山梨醇(/),10 mmol/L的p-NPPC。取20 μL 0.5 μg/mL的PLCζ蛋白樣品溶液加入到200 μL反應混合物中混勻,37 ℃水浴反應30 min,加入0.5 mol/L的NaOH 200 μL終止酶促反應后測定在410 nm波長處的吸收值。酶活單位的定義:pH 7.2、溫度為37 ℃條件下,每分鐘水解p-NPPC產生1 nmol對硝基苯酚所需要的酶量為1個酶活力單位(U)。
2 結果與分析
2.1 pFast-Bac-HTA-PLCζ重組轉移質粒的鑒定
將重組轉移質粒pFast-Bac-HTA-PLCζ用Ⅰ和dⅢ雙酶切后進行瓊脂糖電泳,結果(圖1) 顯示重組質粒經雙酶切后有2條特異性條帶,其中一條為PLCζ基因約為1 800 bp,另一條為載體pFast-Bac-HTA片段約為4 700 bp,均與目的基因大小和載體質粒片段相符,表明載體構建成功,結果見圖1。
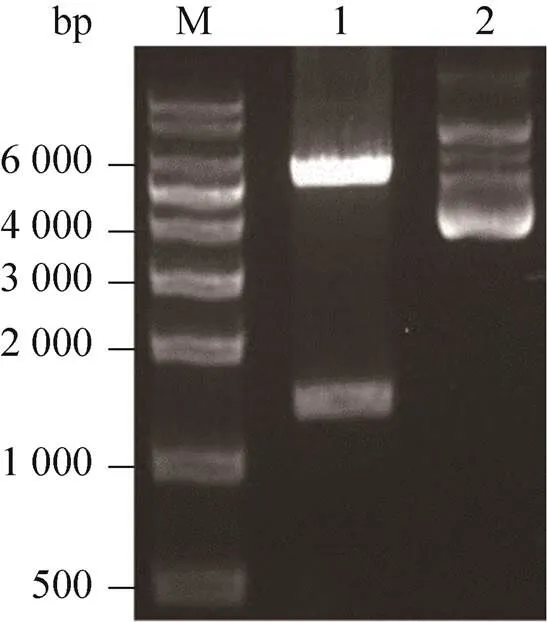
圖1 重組質粒pFast-Bac-HTA-PLCζ雙酶切產物
2.2 Bacmid-PLCζ構建及鑒定
提取重組桿粒,以PLCζ-for/PLCζ-rev為引物對重組桿粒進行PCR鑒定,取出5 μL產物經0.8%瓊脂糖凝膠電泳后,在約1 840 bp處有一特異性條帶,與預期片段大小相符,結果見圖2。插入Bacmid的PLCζ基因經測序表明序列正確,可用于制備重組桿狀病毒顆粒。
2.3 重組Bacmid-PLCζ轉染Sf9細胞及病毒擴增
將重組桿粒Bacmid-PLCζ用脂質體法轉染Sf9昆蟲細胞,27 ℃培養72 h后細胞有明顯的病毒感染跡象,主要表現為細胞生長停滯,胞體脹大變圓,胞核明顯,部分細胞裂解,未轉染的細胞未出現此現象。離心收集上清,標記為PLCζ P1代病毒,收獲病毒液連續傳代后收取P3代桿狀病毒,利用甲基纖維素改良的病毒噬斑法測定P3代病毒滴度為5.2×108PFU/mL (圖3)。

圖2 重組桿粒Bacmid-PLCζ的PCR鑒定

圖3 甲基纖維素噬斑法測定病毒滴度結果分析
2.4 PLCζ重組蛋白的表達及鑒定
重組Bacmid-PLCζ病毒感染Sf9昆蟲細胞,72 h后離心收集細胞培養基,濃縮親和層析純化后的目的蛋白經SDS-PAGE分離,考馬斯亮藍染色顯示相對分子質量約為70 kDa的條帶,其大小與重組人PLCζ蛋白的理論相對分子質量相符,該條帶經飛行時間質譜鑒定為重組人PLCζ蛋白(圖4),其中Mascot得分為151,蛋白序列覆蓋率為50%。用抗人PLCζ蛋白單克隆抗體進行Western blotting分析,成像結果顯示一條相對分子質量約為70 kDa的特異性條帶,與人PLCζ融合蛋白的理論分子質量相符 (圖5)。
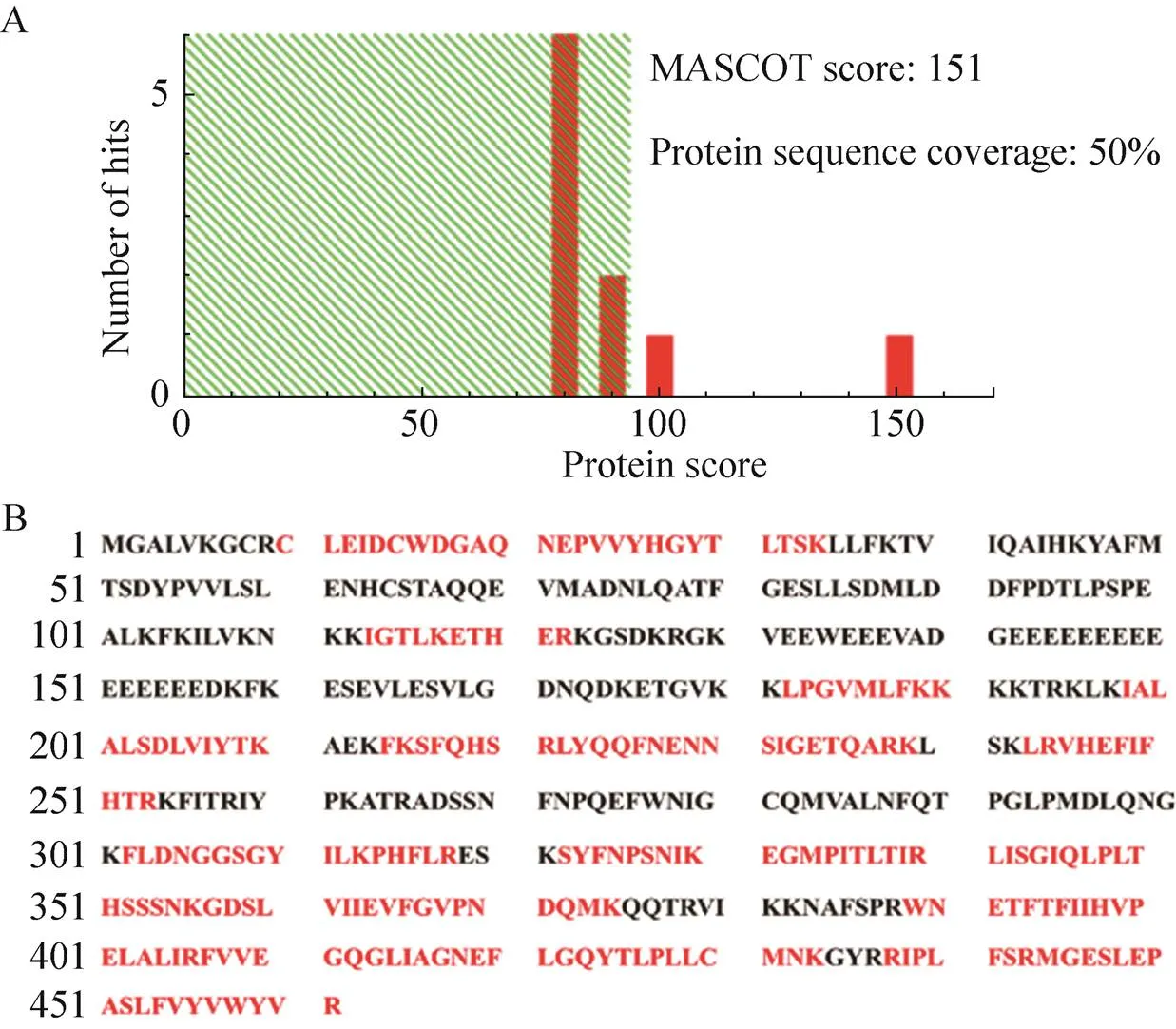
圖4 重組人PLCζ蛋白的肽指紋圖譜鑒定結果
2.5 重組人PLCζ蛋白的活性測定
重組人PLCζ蛋白可水解p-NPPC生成對硝基苯酚,其在410 nm處有最大吸收峰,此吸光值可反映PLCζ水解p-NPPC產生對硝基苯酚的量。將濃度為0.025 g/L的對硝基苯酚溶液梯度稀釋后測定在410 nm處的吸光度值,繪制標準曲線得到的回歸方程為:=94.51?0.043,2=0.999 1,y為對硝基苯酚的濃度(nmol/mL),為不同濃度對硝基苯酚在410 nm處吸光值。酶活力計算公式為:
PLCζ酶活力U (U/mL)=(94.51?0.053)總/30·酶
注:為反應后410,總為反應體系總體積(mL),酶為加入重組人PLCζ蛋白體積(mL)。
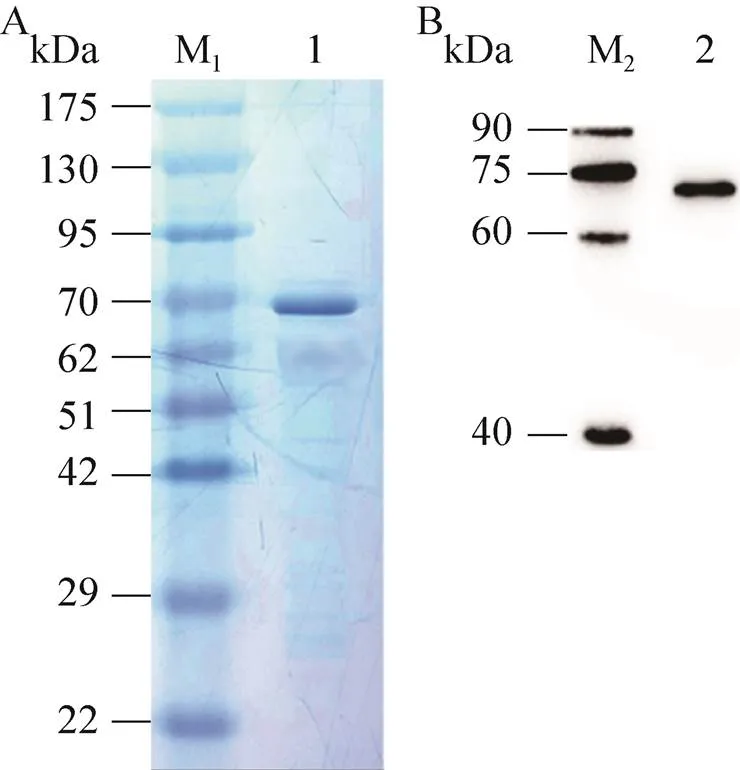
圖5 重組人PLCζ蛋白的SDS-PAGE和Western blotting鑒定
經過優化表達、純化條件,最終獲得重組人PLCζ蛋白酶活力可達到326.8 U/mL。
3 討論
PLCζ蛋白在卵母細胞活化中發揮著重要作用,其異常形式或含量減低可能是某些精子激活卵母細胞失敗型男性不育的根本原因[16-18]。Kouchi等純化出有活性的重組小鼠PLCζ蛋白并將其注射入小鼠卵母細胞,可觀察到類似于精子顯微注射時引發的細胞內Ca2+振蕩,啟動小鼠胚胎發育[19]。雖然這些研究意味著PLCζ在卵母細胞激活缺陷型男性不育治療中的應用潛力,但是關于有活性的人PLCζ重組蛋白的體外表達及純化一直沒有取得大的進展。為更好地分析PLCζ的結構及功能,探究其生物醫學應用,體外表達和純化有活性的人PLCζ重組蛋白顯得尤為重要。
昆蟲桿狀病毒表達系統是繼大腸桿菌、哺乳動物細胞表達系統后的另一個高效的蛋白表達系統。其系統適當的分子伴侶和翻譯后修飾加工體系使其表達的蛋白更加接近體內天然形式同時具有較高的生物學活性[20-21]。為了得到具有生物學活性的PLCζ蛋白,分析其酶學分子特性以及在激活卵母細胞時與其他分子之間的相互作用,進一步探究其作為人工輔助卵母細胞激活生物制劑的可行性,本研究利用桿狀病毒/昆蟲細胞系統表達并分離純化重組人PLCζ蛋白。
為了便于蛋白純化,本研究中我們將人PLCζ基因克隆至含有6×His標簽及AcTEVTM白酶剪切位點的pFastBac HTA質粒中,所表達蛋白經親和層析純化后,重組PLCζ蛋白純度高,無雜蛋白污染。擴增桿狀病毒顆粒后認MOI=5感染Sf9昆蟲細胞,培養72 h后PLCζ蛋白以胞外分泌的形式表達,可以從無血清培養基中分離純化出靶蛋白。本研究中重組桿狀病毒經過3次擴增病毒后滴度經測定為5.2×108PFU/mL,滴度不夠高在一定程度上可能會影響重組蛋白的大批量純化。考慮到影響病毒感染效率的因素有目的基因片段的大小,重組Bacmid DNA的轉染效率、病毒存放時間、保存條件等,本實驗中采用新鮮的病毒進行Sf9昆蟲細胞的感染,并加入了終濃度2%的胎牛血清避光存放于4 ℃,病毒降解可能不是影響桿狀病毒毒力的主要因素,而采用有效的方法提高轉染效率是提高病毒滴度的關鍵。有研究表明通過多次傳代可以提高病毒毒力,但應當注意桿狀病毒在細胞中多次傳代后可能引起基因組的改變,出現多角體表型變化,導致蛋白表達水平降低[22]。為了減少重組桿狀病毒在傳代后存在突變導致蛋白表達水平降低的可能,通過擴大轉染體積、離心濃縮病毒顆粒等方法來獲取低代數、高滴度的重組病毒是后續研究中需要實現的目標。影響重組蛋白表達的因素除了表達載體選擇和目的基因本身大小、重組病毒感染效率之外還取決于宿主細胞狀態及培養條件和蛋白收獲時間[23-24]。本實驗中發現宿主細胞狀態對目的蛋白的產量至關重要,感染前應選擇傳代次數少、細胞存活率高于95%、無異常形態的健康對數生長期細胞進行重組病毒接種;宿主細胞狀態不佳時無論是通過增加病毒接種量、延長細胞培養時間還是增加宿主細胞數量均不能提高蛋白表達量。考慮到表達的重組蛋白會被細胞培養時自身蛋白酶所降解,我們選擇在感染72 h作為收獲PLCζ蛋白的最佳時間點。為了把目的蛋白從細胞培養基中分離出來,同時還要保持其生物學活性,蛋白純化過程中盡量在低溫環境中進行[25]。收集感染病毒后的昆蟲細胞培養基后,可以加入蛋白酶抑制劑以防止目的蛋白降解,利用重組蛋白上所帶的His標簽與Ni2+親和層析達到分離純化目的蛋白與雜蛋白,最后經高濃度咪唑洗脫得到目的蛋白。考馬斯亮藍染色及Western blotting檢測蛋白大小為68 kDa左右,與人PLCζ蛋白的理論分子大小一致,該蛋白經電離飛行時間質譜鑒定為PLCζ蛋白,經酶學測定顯示有生物學活性,說明本方法可成功獲得有活性的重組人PLCζ蛋白。
PLCζ蛋白是PLC家族的最新成員,目前關于人PLCζ蛋白結構及功能的研究鮮見報道。本研究利用桿狀病毒/昆蟲細胞真核表達系統獲得了在結構及功能上都更接近天然蛋白特性的重組人PLCζ蛋白,這為下一步進行人PLCζ蛋白晶體結構解析的生物學研究奠定了堅實的基礎,也為加深對PLCζ在卵母細胞激活缺陷型男性不育診斷和治療上的探索提供了可能。
[1] Saunders CM, Larman MG, Parrington J, et al. PLC zeta: a sperm-specific trigger of Ca2+oscillations in eggs and embryo development. Development, 2002, 129(15): 3533–3544.
[2] Swann K, Saunders CM, Rogers NT, et al. PLCζ(zeta): a sperm protein that triggers Ca2+oscillations and egg activation in mammals. Semin Cell Dev Biol, 2006, 17(2): 264–273.
[3] Xu HY, Deng K, Luo QB, et al. High Serum FSH is associated with brown oocyte formation and a lower pregnacy rate in human IVF parctice. Cell Physiol Biochem, 2016, 39(2): 677–684.
[4] Nomikos M, Elgmati K, Theodoridou M, et al. Male infertility-linked point mutation disrupts the Ca2+oscillation-inducing and PIP2, hydrolysis activity of sperm PLCζ. Biochem J, 2011, 434(2): 211–217.
[5] Heytens E, Parrington J, Coward K, et al. Reduced amounts and abnormal forms of phospholipase C zeta (PLCζ) in spermatozoa from infertile men. Hum Reprod, 2009, 24(10): 2417–2428.
[6] Ramadan WM, Kashir J, Jones C, et al. Oocyte activation and phospholipase C zeta (PLCζ): diagnostic and therapeutic implications for assisted reproductive technology. Cell Commun Signal, 2012, 10(1): 12.
[7] Ozil JP, Banrezes B, Tóth S, et al. Ca2+oscillatory pattern in fertilized mouse eggs affects gene expression and development to term. Dev Biol, 2006, 300(2): 534–544.
[8] Tavalaee M, Nasr-Esfahani MH. Expression profile of,, and-in association with fertilization potential, embryo development, and pregnancy outcomes in globozoospermic candidates for intra-cytoplasmic sperm injection and artificial oocyte activation. Andrology, 2016, 4(5): 850–856.
[9] Amdani SN, Yeste M, Jones C, et al. Phospholipase C zeta (PLCζ) and male infertility: clinical update and topical developments. Adv Biol Regul, 2016, 61: 58–67.
[10] Yoon SY, Eum JH, Lee JE, et al. Recombinant human phospholipase C zeta 1 induces intracellular calcium oscillations and oocyte activation in mouse and human oocytes. Hum Reprod, 2012, 27(6): 1768–1780.
[11] Nomikos M, Yu YS, Elgmati K, et al. Phospholipase Cζ rescues failed oocyte activation in a prototype of male factor infertility. Fertil Steril, 2013, 99(1): 76–85.
[12] Li ZH, Wang QJ. Expression, purification and characterization of catalytic domain of protein kinase D1 in baculovirus-insect cell expression system. Chin J Biotech, 2014, 30(8): 1291?1298 (in Chinese). 李志紅, Wang QJ. 蛋白激酶D1催化結構域在昆蟲細胞/桿狀病毒表達系統內的表達、純化和活性測定. 生物工程學報, 2014, 30(8): 1291?1298.
[13] Luckow VA, Lee SC, Barry GF, et al. Efficient generation of infectious recombinant baculoviruses by site-specific transposon-mediated insertion of foreign genes into a baculovirus genome propagated in. J Virol, 1993, 67(8): 4566–4579.
[14] Baer A, Kehn-Hall K. Viral concentration determination through plaque assays: using traditional and novel overlay systems. J Vis Exp, 2014, (93): e52065.
[15] Flieger A, Gong SM, Faigle M, et al. Critical evaluation of p-nitrophenylphosphorylcholine (p-NPPC) as artificial substrate for the detection of phospholipase C. Enzyme Microb Technol, 2000, 26(5/6): 451–458.
[16] Kashir J, Heindryckx B, Jones C, et al. Oocyte activation, phospholipase C zeta and human infertility. Hum Reprod Update, 2010, 16(6): 690–703.
[17] Parrington J. Does a soluble sperm factor trigger calcium release in the egg at fertilization? J Androl, 2001, 22(1): 1–11.
[18] Rogers NT, Halet G, Piao Y, et al. The absence of a Ca2+signal during mouse egg activation can affect parthenogenetic preimplantation development, gene expression patterns, and blastocyst quality. Reproduction, 2006, 132(1): 45–57.
[19] Kouchi Z, Fukami K, Shikano T, et al. Recombinant phospholipase Czeta has high Ca2+sensitivity and induces Ca2+oscillations in mouse eggs. J Biol Chem, 2004, 279(11): 10408–10412.
[20] Lin SY, Chen GY, Hu YC. Recent patents on the baculovirus systems. Recent Pat Biotechnol, 2011, 5(1): 1–11.
[21] Zhang ZG, Jing Y, Barford D. Recombinant expression and reconstitution of multiprotein complexes by the USER cloning method in the insect cell-baculovirus expression system. Methods, 2016, 95: 13–25.
[22] Kool M, Ahrens CH, Vlak JM, et al. Replication of baculovirus DNA. J Gen Virol, 1995, 76(9): 2103–2118.
[23] Ren YY, Chen D, Guo YZ, et al. Expression of human retinol-binding protein 4 in insect baculovirus system and preparation of its polyclonal antibody. Chin J Biotech, 2013, 29(7): 974?985 (in Chinese). 任玉瑩, 陳丹, 郭玉爭, 等. 人視黃醇結合蛋白4在桿狀病毒系統中的表達及其多克隆抗體制備. 生物工程學報, 2013, 29(7): 974?985.
[24] Jarvis DL. Developing baculovirus-insect cell expression systems for humanized recombinant glycoprotein production. Virology, 2003, 310(1): 1–7.
[25] Wingfield PT. Overview of the purification of recombinant proteins. Curr Protoc Protein Sci, 2015, 80(6): 1–35.
Expression, purification and characterization of recombinant PLCζ protein in baculovirus-insect cell expression system
Xin Chen1, Yueyue Hu1, Hongyi Xu1, Xiaoyan Wang2, and Kai Deng1
1Reproductive Medicine Center, Renmin Hospital, Hubei University of Medicine, Shiyan 442000, Hubei, China 2 School of Basic Medical Sciences, Hubei University of Medicine, Shiyan 442000, Hubei, China
PLCζ is a new isoenzyme of the PLC family which plays an important role in activating mammalian oocytes. In recent years, large-scale expression and purification of active PLCζ proteinfor structural biology research has not been successful. In this study, the recombinant human PLCζ protein was expressed and purified in the baculovirus expression system. First, the full length of human PLCζ gene was cloned into the pFastBac-HTA plasmid to form the recombinant donor plasmid that was further transformed into DH10Baccells to construct the recombined bacmid by the site-specific transposition that was screened by resistance and blue-white spots. Then the bacmid was transfected to Sf9 insect cells via cellfectin to package the recombinant baculovirus. After the amplification of the recombinant baculovirous, the recombinant protein was expressed from the cells transduced by the recombinant baculovirus and was purified by Ni-NTA resin. Purified protein was identified by Western blotting and time-of-flight mass spectrometry and the enzyme activity was determined. The results showed that the recombinant PLCζ protein in the Sf9 cells was achieved at 72 hours after baculovirus infection and expressed in secreted form in cell culture medium. The recombinant protein purified by Ni2+affinity column was identified as PLCζ by Western blotting and ionization time-of-flight mass spectrometry and the enzyme activity was up to 326.8 U/mL. The experimental results provide a reference for the large-scale production and biological application of recombinant human PLCζ protein.
PLCζ protein, insect cell, baculovirus expression system, expression, purification, identification
December 6, 2018;
February 18, 2019
National Natural Science Foundation of China (No. 81401200), Natural Science Foundation of Hubei Province of China (Nos. 2018CFB219, 2013CFB479), Research Project of Hubei Provincial Department of Education (Nos. B2015478, B2015493).
Xiaoyan Wang. Tel: +86-719-8875302; E-mail: xywangdk@163.com
Kai Deng. Tel: +86-719-8637123; E-mail: dkeanig@163.com
國家自然科學基金 (No. 81401200),湖北省自然科學基金 (Nos. 2018CFB219,2013CFB479),湖北省教育廳科學技術研究項目 (Nos. B2015478,B2015493) 資助。
10.13345/j.cjb.180507
陳鑫, 胡玥玥, 徐鴻毅, 等. 重組人PLCζ蛋白在昆蟲細胞/桿狀病毒表達系統內的表達、純化及活性測定. 生物工程學報, 2019, 35(6): 1135–1142.
Chen X, Hu YY, Xu HY, et al. Expression, purification and characterization of recombinant PLCζ protein in baculovirus-insect cell expression system. Chin J Biotech, 2019, 35(6): 1135–1142.
(本文責編 陳宏宇)
