Lager啤酒酵母RLM1基因調控對其抗自溶性能的影響
2019-07-10 08:38:36王金晶李夢琦侯丹許維娜鄭飛云劉春鳳鈕成拓李崎
生物工程學報 2019年6期
王金晶,李夢琦,侯丹,許維娜,鄭飛云,劉春鳳,鈕成拓,李崎
?
Lager啤酒酵母基因調控對其抗自溶性能的影響
王金晶1,2,李夢琦1,2,侯丹1,2,許維娜1,2,鄭飛云1,2,劉春鳳1,2,鈕成拓1,2,李崎1,2
1 江南大學 生物工程學院 工業生物技術教育部重點實驗室,江蘇 無錫 214122 2 江南大學 釀酒科學與工程研究室,江蘇 無錫 214122
啤酒酵母的自溶會嚴重影響啤酒的品質,而酵母的質量也被認為是啤酒釀造的關鍵因素之一。前期在啤酒酵母自溶的研究中發現細胞完整性途徑中重要的轉錄因子基因與酵母自溶有密切關系。本研究在啤酒酵母單倍體菌株中對進行敲除與過表達,發現敲除后,酵母菌抗自溶性能差,而過表達則有助于酵母的抗自溶。另外,發現基因的敲除影響了酵母的抗滲透壓性能、細胞壁損傷的耐受性、抗氮饑餓性能和溫度耐受性。研究發現細胞壁組裝及DNA損傷應答相關基因的表達隨的過表達與敲除而調整,而CWI途徑中其他相關基因的調控方式并沒有明顯的規律,推測可能主要影響了CWI途徑中基因的表達,進而提高啤酒酵母在惡劣環境中的抗逆性。此研究結果對于進一步選育抗自溶啤酒酵母以及了解啤酒酵母的自溶機制提供了基礎。
lager啤酒酵母,抗自溶性能,,細胞壁完整性
環境脅迫一直是發酵工業中最普遍且不容忽視的問題之一。酵母是飲料工業中常用的菌種之一,也被認為是啤酒釀造的關鍵因素之一。啤酒發酵過程中,酵母細胞在多次脅迫下會發生自降解,這種自降解過程通常發生在啤酒的后酵期,對啤酒的品質有顯著影響[1]。啤酒釀造過程中如有超過5%的酵母細胞自溶就會對啤酒風味、泡沫穩定性、膠體穩定性造成影響并可能帶來微生物污染的危害[2]。
啤酒發酵后期發生的酵母自溶主要是由營養缺乏、應激反應以及酒精壓力引起的。細胞壁是細胞的第一道防線,對環境中的不利因素作出反應。細胞壁完整性 (Cell wall integrity,CWI)、細胞壁的組織和生物合成主要由該通路負責[2-3]。以CWI通路激活為主導的細胞轉錄重編程將細胞從各種應激條件下解救出來。酵母細胞壁上存在壓力傳感因子,包括Mtl1、Mid2、Wsc1、Wsc2以及Wsc3,當這些因子感受到壓力時,觸發了絲裂原活化蛋白激酶(Mitogen-activated protein kinase,MAPK) 級聯反應,在細胞內進行全局轉錄調控進而形成不同的應激反應來維持酵母細胞的生存[3-4]。在CWI通路下游,Slt2磷酸化后激活包括Swi4/Swi6和Rlm1[5]在內的轉錄因子。據報道,Slt2和Rlm1是密切相關的,在細胞壁受到損傷時,Rlm1傳導了大多數依賴于Slt2的轉錄激活作用,同時,CWI途徑與其他信號通路的轉錄因子之間存在的連接也必須通過Rlm1依賴的通路進行調控[6]。此外,最近的研究顯示,基因參與了乙酰轉移酶 (Spt-Ada-Gcn5-acetyltransferase,SAGA) 復合物對細胞壁應激反應的轉錄調控,敲除將導致CWI通路失活[4]。
自溶過程中,啤酒酵母細胞逐漸喪失生存能力,許多與應激反應相關的基因受到調控,以適應環境的變化。酵母細胞壁結構和形態將逐漸改變以應對環境中的脅迫壓力[1,7]。前期研究發現在酵母自溶過程中,CWI途徑中的壓力傳感因子編碼基因包括和表達水平下調,而CWI途徑中和表達水平發生上調[8]。同時,具有較強抗自溶能力的酵母細胞表現出剛性的細胞壁結構和較強的抗應激能力[9]。雖然轉錄因子Rlm1在CWI通路中的作用在釀酒酵母中已有相關的報道,但是其在啤酒酵母抗自溶方面的研究尚少,同時,啤酒酵母由于其遺傳背景的復雜性表現出與釀酒酵母不同的發酵及生理特性[10-11]。因此,本研究對lager型啤酒酵母菌中的進行調控,研究其對啤酒酵母抗自溶能力的影響及作用機制。
1 材料與方法
1.1 材料
1.1.1 菌種和培養基
菌株H5為本研究室保藏的啤酒酵母單倍體菌株;克隆擴增質粒pMD19-T simple及大腸桿菌宿主菌JM109均購于TaKaRa公司;質粒pUG6為本研究室保藏[12]。
LB培養基 (1 L):10 g蛋白胨,5 g酵母抽提物,10 g NaCl,pH 7.4,固體培養基添加20 g瓊脂粉,121 ℃滅菌20 min[12]。含有氨芐青霉素的抗性培養基終濃度為100 μg/mL。
YPD培養基 (1 L):10 g酵母抽提物,20 g蛋白胨,20 g葡萄糖,pH 6.0,115 ℃滅菌15 min。含有G418的抗性培養基終濃度分別為200 μg/mL、300 μg/mL和400 μg/mL。含有NaCl的抗性培養基的濃度為1 mol/L。米卡芬凈抗性培養基終濃度為30 ng/mL。Calcofluor white抗性培養基終濃度為10 μg/mL。
YNB-N培養基 (1 L):1.7 g無氨基酸無硫酸銨酵母氮源,20 g葡萄糖,固體培養基添加20 g瓊脂粉,115 ℃滅菌15 min[12]。
1.1.2 試劑
質粒抽提試劑盒、大腸桿菌感受態制備試劑盒、限制性內切酶及連接酶購自TaKaRa公司;YNB合成培養基、蛋白抽提試劑盒、溶菌酶購自生工生物工程 (上海) 股份有限公司;Fluorescentbrightener 28 (Calcofluor white) 購自Sigma-Aldrich公司;米卡芬凈購自上海灝云藥業。
1.2 主要方法
1.2.1基因的敲除與過表達
根據釀酒酵母的序列設計引物RLM1-F/RLM1-R擴增啤酒酵母基因,測序后以所測的序列設計中斷引物RLM1-LF/RLM1-LR。各引物如表1所示。以pUG6質粒為模板,RLM1-LF/RLM1-LR為引物用短側翼序列PCR介導的一步法 (SFH-PCR)擴增帶有上下游同源臂的基因,DNA片段經純化后進行測序,構建成功后被用于轉化啤酒酵母單倍體菌株H5,轉化子涂布于新鮮的YPD平板,24 h后影印至含有200 μg/mL的G418抗性平板上,28 ℃培養48 h,挑取陽性轉化子進行菌落PCR驗證,獲得敲除菌株。

表1 構建菌株所用引物
以基因組DNA為模板,以通用引物EF3、EF4擴增啤酒酵母18S rDNA序列,測序后設計引物擴增其上下游各200 bp左右的片段作為同源臂。通過PCR獲得啟動子與目的基因,同時以pUG6為模板,PCR獲得抗性基因,表1為基因擴增所用引物及相應的酶切位點。將上述片段以TA克隆的方式連接至pMD19-T simple質粒上,經Ⅰ和Ⅰ雙酶切后線性轉化至H5。在G418抗性平板上挑選陽性菌落并進行菌落PCR驗證,獲得過表達菌株。根據G418抗性的強弱篩選高拷貝數的過表達菌株[12]。挑取陽性轉化子接種至10 mL YPD培養基中,28 ℃、200 r/min振蕩培養12 h,3 000 ×離心5 min,收集菌體并在無菌生理鹽水中重懸,調節菌體濃度至600=1.0,梯度稀釋后點滴至G418濃度為200 μg/mL、300 μg/mL及400 μg/mL的抗性平板上[12]。28 ℃培養48 h,觀察菌株的生長狀況,最終篩選獲得含有高拷貝數的過表達菌株。
1.2.2 Rlm1蛋白的SDS-PAGE分離及質譜鑒定
將出發菌株H5和過表達菌株在5 mL YPD培養基中28 ℃、200 r/min振蕩培養12 h,后將菌液全部轉接至50 mL YPD培養基,28 ℃振蕩培養3–5 h,至菌液濃度為600=1.0–1.5,3 000×離心5 min,棄上清收集菌體,用PBS緩沖液清洗 3次,再用Tris-HCl緩沖液重懸菌體,加入蛋白酶抑制劑復合物,超聲破碎提取蛋白質[12-13]。Rlm1是核蛋白,因此本研究采用細胞核蛋白/漿蛋白抽提試劑盒同時抽提細胞質蛋白和核蛋白。將蛋白樣品與上樣緩沖液混合,沸水浴5 min后上樣,進行SDS-PAGE檢測。將目的條帶切下并進行MALDI-TOF-TOF/MS分析,鑒定其是否為目的蛋白Rlm1[12]。
1.2.3 抗自溶指數測定
參考前期研究的酵母抗自溶指數檢測方法,抗自溶指數為 (260/280)/死亡率,跟蹤測定出發菌H5、基因敲除菌株及過表達菌株的260、280和死亡率,計算獲得抗自溶指數來表征各菌株的抗自溶性能[14]。
1.2.4 脅迫耐受性分析
將H5、基因敲除菌株和過表達菌株培養12 h后離心收集菌體,重懸于無菌水中,分別取樣點滴至NaCl抗性平板、熒光增白劑抗性平板和米卡芬凈抗性平板,同時1 000倍稀釋后涂布于YNB-N平板,分析其對滲透壓、細胞壁損傷和氮饑餓的抗性;再將菌液點樣于YPD平板,在22 ℃、28 ℃、37 ℃和42 ℃培養,檢測其對溫度的耐受性。
1.2.5 啤酒發酵實驗
取一環保藏的斜面菌轉接至 10 mL 12°P麥汁試管中,于 28 ℃下活化24 h,后以1%的接種量轉接至含有70 mL 12°P麥汁的三角瓶中于25 ℃培養48 h,按2×107CFU/mL酵母細胞濃度轉接至含2 L 12°P麥汁的3 L帶發酵栓的錐形瓶中,11 ℃發酵5 d。主酵結束后收集發酵液及酵母細胞進行理化指標分析。
1.2.6 細胞形態學分析
發酵結束后,收集酵母細胞,使用5%戊二醛 (0.1 mol/L磷酸緩沖液,pH 7.2) 進行前固定,之后使用0.1 mol/L磷酸緩沖液漂洗酵母細胞,用1%鋨酸 (0.1 mol/L磷酸緩沖液,pH 7.2) 進行后固定,再用0.1 mol/L磷酸緩沖液漂洗,乙醇梯度脫水,樣品進行臨界點干燥后轉載至樣品臺上,離子濺射儀鍍膜后置于掃描電子顯微鏡下觀察其細胞形態[13],醋酸雙氧鈾和檸檬酸鉛染色后進行超薄切片,置于透射電子顯微鏡下觀察[13-14]。
1.2.7 qRT-PCR分析
收集酵母細胞重懸于無菌水中,用試劑盒提取總RNA并按TaKaRa公司反轉錄試劑盒PrimeScript RT reagent Kit進行反轉錄,獲得的cDNA保存待用。使用TaKaRa公司的SYBR Premix Ex試劑盒和Roche Light Cycler 480 Ⅱ PCR擴增儀 (Roche Diagnostics) 對CWI途徑中關鍵基因進行qRT-PCR檢測[13],條件如下: 95 ℃預變性30 s;變性95 ℃ 5 s,退火/延伸55 ℃ 20 s,循環40次;95 ℃ 10 s,65 ℃ 1 min,95 ℃ 10 s。每個樣品3個平行,取平均值作為計算的基礎。
2 結果與分析
2.1 RLM1基因的敲除及過表達
用引物RLM1-F/RLM1-R以啤酒酵母單倍體菌株H5的基因組DNA為模板,PCR擴增目的基因。產物純化后測序,測序結果表明,啤酒酵母H5的序列長度為2 043 bp[14],與NCBI公布的釀酒酵母S288c的相似性達到98%。根據H5的基因序列設計中斷引物RLM1-LF/RLM1-LR,同時以pUG6質粒為模板擴增,構建基因中斷盒[12],總長度為1 704 bp。片段線性化后轉化啤酒酵母單倍體菌株H5,在含有200 μg/mL G418的YPD平板上篩選獲得陽性轉化子,并進行PCR驗證,結果如圖1A所示。出發菌H5應獲得大小為2 043 bp的片段,陽性轉化子應獲得1 704 bp的片段,結果與預期相符,最終得到基因敲除菌命名為HΔ。
將18S rDNA上游同源臂、啟動子目的基因抗性基因18S rDNA下游同源臂依次連接至pMD19-T simple上,構建過表達載體。獲得的質粒經Ⅰ和Ⅰ雙酶切后轉化至啤酒酵母單倍體菌株H5,與H5基因組中18S rDNA發生同源重組,替換原有片段,采用影印法篩選陽性轉化子,目的基因拷貝數的高低以重組菌G418抗性的強弱來表征。即G418抗性強的重組菌,其拷貝數高,對應的拷貝數也高[12,15]。過表達所獲得的陽性重組菌影印至濃度為200、300、400 μg/mL的G418抗性平板上。結果發現,重組菌最高能耐受400 μg/mL G418,28 ℃培養48 h后,在含有400 μg/mL G418的抗性平板上挑選生長情況較好的菌株進行PCR驗證,分別以Up-1-F和Up-1-R、Up-2-F和Up-2-R、Down-1-F和Down-1-R作為引物,以H5為對照。目標片段大小應為 964 bp、944 bp及856 bp,結果 (圖1A) 與預期相符。同時,提取篩選獲得的過表達菌株的細胞核蛋白和細胞質蛋白進行SDS-PAGE檢驗,結果如圖1B所示。目的條帶被切下后進行MALDI- TOF-TOF/MS分析,其中得分最高的蛋白質為Rlm1 (登錄號gi|6325168),說明基因在重組菌中得到了過表達[12],將獲得的過表達菌株命名為H/。
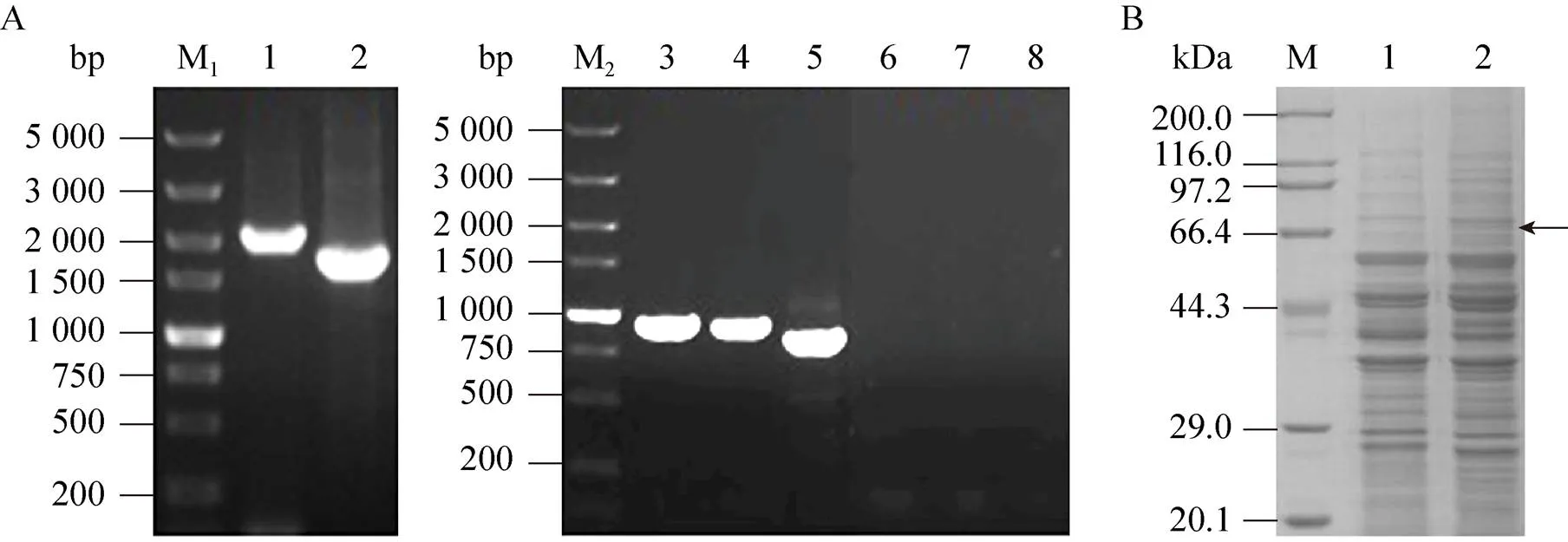
圖1 重組菌H-rlm1Δ及H/rlm1的構建及驗證
2.2 RLM1基因調控對菌株抗自溶性能的影響
啤酒酵母對于啤酒的釀造至關重要,啤酒優良的風味主要是由其代謝產物產生,而啤酒酵母在發酵后期發生的自溶會導致細胞內容物的滲漏和溶出,這是啤酒中雜異味的主要來源。在有些情況下,例如在貯酒期間環境壓力大、代謝產物積累和營養物質枯竭等,啤酒酵母在短時間內發生大量的自溶,對啤酒的質量造成劇烈的影響。具有較好抗自溶能力的啤酒酵母將對啤酒的風味有積極的貢獻。
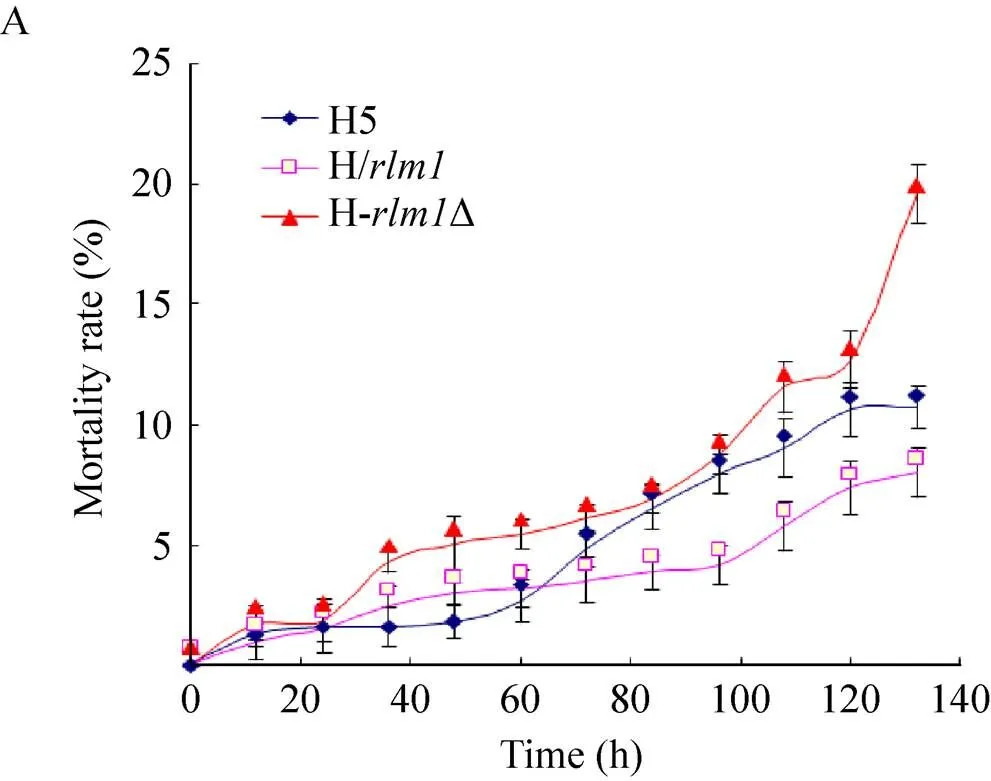
將H-Δ、H/與H5培養至穩定期后期,收集酵母泥,用檸檬酸緩沖液洗滌2–3遍,每100 mL緩沖液中加入1 g酵母泥,30℃下進行模擬自溶,跟蹤260、280和死亡率的數值[12],計算抗自溶指數,各酵母的抗自溶能力以抗自溶指數來表征[14]。圖2為3株菌的抗自溶指數和死亡率的比較結果。敲除菌H-Δ死亡速度最快,抗自溶指數數值相對最低,表明其自溶程度高,抗自溶性能差。開始時,H/死亡速度快于H5,但隨后逐漸減慢,可發現其抗自溶指數高于H5,表明其抗自溶性能較好。
前期研究中發現在不同酵母及不同自溶態下的轉錄水平各異,抗自溶性能好的酵母中的轉錄水平較高,而抗自溶性能差的酵母中的轉錄水平較低[8,12]。本研究結果也進一步印證了前期研究的結果,同時可以推斷的敲除影響了啤酒酵母的抗自溶能力,而高表達量的可以減緩自溶的進程。
2.3 RLM1基因調控對菌株脅迫耐受性的影響
2.3.1 滲透壓及細胞壁損傷耐受性分析
在含有1 mol/L NaCl的YPD平板上對H5、HΔ和H/進行點樣分析,于28 ℃培養48 h后觀察其生長情況,檢測各菌株對滲透壓的耐受性,結果如圖3所示。H/對滲透壓的抗性稍強于出發菌H5,而HΔ對滲透壓的抗性明顯弱于出發菌H5[12]。
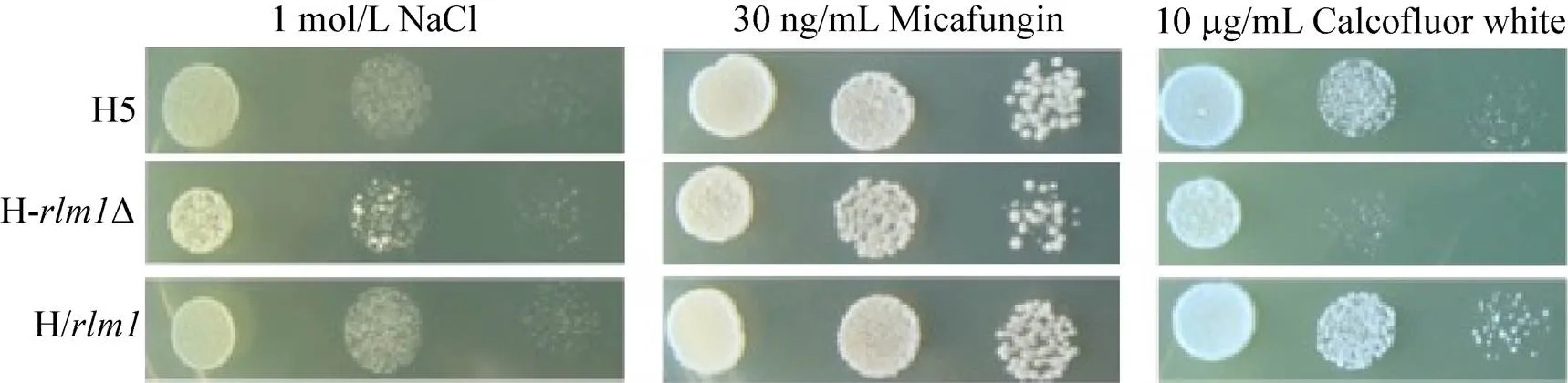
圖3 重組菌與原始菌對滲透壓及細胞壁損傷耐受性分析
米卡芬凈 (Micafungin) 能夠非競爭性抑制酵母細胞壁必需成分β-1,3-葡聚糖的合成,是葡聚糖合成酶的抑制劑[9],可導致細胞壁合成的缺陷。熒光增白劑 (Calcofluor white) 能夠與細胞壁組分幾丁質結合,細胞壁有破損的細胞會對熒光增白劑敏感[17]。米卡芬凈和熒光增白劑均能造成細胞壁壓力,進而引起細胞壁的損傷。將H5、HΔ和H/分別點滴至含有30 ng/mL米卡芬凈和含有10 μg/mL熒光增白劑的抗性平板上[12],在28 ℃條件下培養48 h,觀察其生長情況,評價各菌株對細胞壁損傷的耐受性情況。結果如圖3所示,HΔ對米卡芬凈和熒光增白劑均比較敏感,特別在熒光增白劑平板上,其生長被強烈抑制。H/對米卡芬凈的抗性與H5類似,但H/對熒光增白劑的抗性略強于H5。結果表明的缺失嚴重影響了細胞對幾丁質相關的細胞損傷的抗性。
2.3.2 溫度耐受性分析
在YPD平板上進行H5、HΔ和H/進行點樣,分別放置于22 ℃、28 ℃、37 ℃和42 ℃環境中培養,檢驗其對溫度的耐受性情況。三株菌在42 ℃均不能生長 (結果未展示),如圖4所示HΔ在低溫 (22 ℃) 和高溫 (37 ℃) 下的生長能力與H5相比略有下降,H/的生長狀態與H5相當,說明敲除后菌株對溫度的耐受性受到一定的影響。另外,lager型啤酒發酵溫度一般在11 ℃左右,因此考察了3株菌在11 ℃條件下的生長能力,在低溫脅迫的情況下,H-Δ生長能力稍差一些,但沒有明顯的區別。
2.3.3 氮饑餓耐受性分析
將H5、HΔ和H/菌液稀釋后涂布于YNB-N平板,28 ℃培養48 h,檢測各菌株對氮饑餓的耐受性情況。菌落計數結果為H5菌落數為70±5,HΔ菌落數為21±4,H菌落數為92±7。從饑餓抗性來看,H/強于H5,而H-Δ明顯弱于H5,結果表明表達對菌株的氮饑餓耐受性起到了增強作用。
2.4 RLM1基因調控對菌株發酵性能及抗自溶性能的影響
將重組菌與原始菌用于啤酒發酵試驗,在主發酵結束時,測定了發酵液的幾個主要參數,以評價不同菌株的發酵能力。如表2所示,從發酵度和真實濃度來看,不同菌株的發酵能力相當,主要風味化合物包括雙乙酰、乙醛、乙酸乙酯、乙酸異戊酯、異戊醇、異丁醇和雙硫醚 (DMS) 含量也沒有太大的差別,在正常發酵過程中,健康的酵母不會發生自溶,細胞內蛋白酶也不會水解自身物質,主酵結束后對發酵液相關指標的測定結果表明基因調控對啤酒酵母的發酵能力及啤酒的主體風味沒有直接的影響。
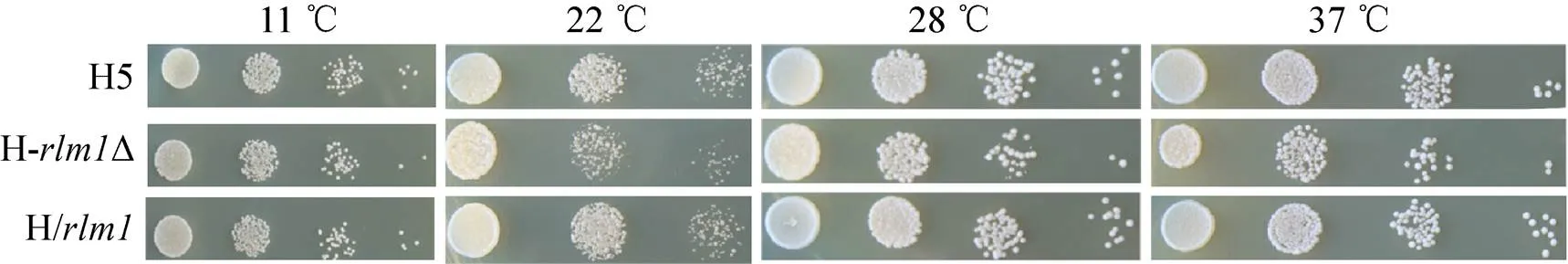
圖4 重組菌與原始菌對溫度耐受性的比較
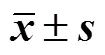
表2 啤酒發酵液指標分析
為了解釋不同菌株抗自溶能力的差異,收集發酵后細胞進行了掃描電鏡 (SEM) 及透射電鏡 (TEM) 分析。如圖5所示,與H5相比,H/細胞體積略變小,呈橢圓形,H-Δ細胞體積略大,且在發酵后更容易表現出皺縮及內凹的形態。與原始菌株比較,的敲除可能影響酵母細胞的大小及在發酵過程中的耐受性。但是,比較不同菌株的生長曲線,3個菌株之間沒有顯著的差異。因此,推測基因修飾導致了啤酒酵母的形態改變。同時,啤酒發酵結束后收集3株菌進行透射電鏡分析,結果與SEM結果吻合。H5細胞核區完整,胞質液泡大 (圖6A)。在H/細胞發現液泡變小且分散 (圖6B–C),H-Δ細胞中則發現了逐漸的染色質凝聚和核區的分散 (圖6D–E)。基因敲除的菌株中明顯出現了細胞損傷和衰老的現象。
Rlm1是CWI途徑中重要的轉錄因子,對其編碼基因的調控可能直接影響細胞完整性途徑中其他基因的表達。分析了發酵開始與結束時與CWI通路和細胞壁構建相關的基因 (包括、、、、、、、、、、、、、、、、、、、、、)[17-18]的相對mRNA水平,其中許多基因在調節啤酒酵母的自溶過程中起著重要作用。結果發現,H/中與CWI途徑相關的基因在啤酒發酵結束時大部分發生上調 (圖7),上調最多的基因為,該基因涉及細胞壁組裝[19]和DNA損傷應答[20],其表達水平較啤酒發酵開始時上調了12.75倍。而據報道參與細胞壁合成的上調了6.29倍。、、、、、、、、、在H/中表達相對上調,其他基因的調控不明顯。在H5菌株中也發現了類似的調控模式,盡管的上調并不充分。然而,與H/相比,H5中基因沒有發現明顯的調控。另一方面,大部分基因的調控在H-Δ中發生下調,基因表達也發生明顯下調,而和基因表達上調。是與細胞核的穩定性及細胞形態相關的基因[21],該基因在H-Δ發生了明顯上調 (11.76倍)。細胞在發酵過程中受到損傷時,基因可能發生上調以維持細胞的形態穩定。與發酵開始時相比,轉錄輔助因子Swi6編碼基因的表達水平上調了7.45倍。、等細胞壁應激傳感基因[22]在3株菌中均呈現相似的調控水平。

圖5 H5 (A)、H/rlm1 (B)及H-rlm1Δ (C)的掃描電鏡圖(×8 000)
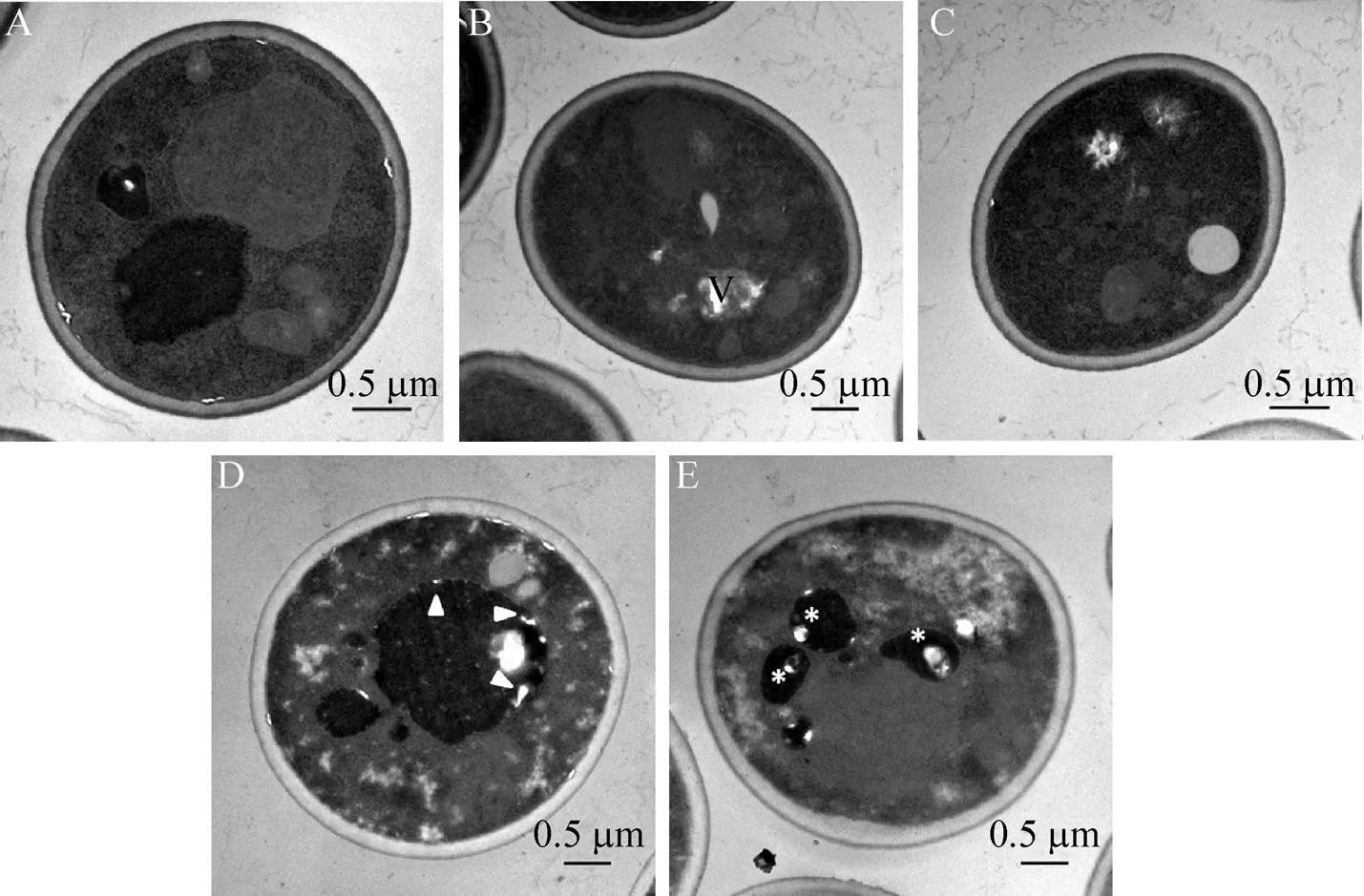
圖6 H5(A)、H/rlm1 (B、C)及H-rlm1Δ (D、E)的透射電鏡圖(×25 000)

圖7 H5、H-rlm1Δ和H/rlm1中細胞壁完整性途徑相關基因調控水平分析
3 討論
在工業生產中,由于受到成本的限制,啤酒酵母需要被重復使用,酵母的自溶一般會發生啤酒發酵的后酵期以及在多次反復回用之后,目前國內大多數啤酒廠的啤酒酵母大多使用代數為 5代,第5代之后的酵母的生理性能發生大幅度下降,自溶率顯著增加,造成釀造的啤酒的品質與風味受到極大的影響。而國際上優質的啤酒酵母一般可使用至15代。國內啤酒廠常用的啤酒酵母在傳代使用過程中,大部分啤酒酵母在使用至第3代時不會立即出現自溶的現象,而第4代及第5代發酵結束后,抗自溶性能較弱的酵母細胞就會出現明顯的自溶現象[16]。啤酒酵母抗自溶性能的提高能夠有效地增加酵母的回用次數,進一步提高生產效率,降低成本。酵母細胞的應激反應被認為是一種進化適應,使酵母能夠對不利的環境條件作出反應,并保持細胞的繁殖力。研究表明轉錄因子Rlm1參與了MAPK通路和細胞壁完整性 (CWI) 途徑的激活[23],其編碼基因是Mpk1在CWI通路下游的功能基因,也參與了細胞壁完整性的維護。如前所述,基因在酵母中表達不同,其在抗自溶能力較強的啤酒酵母菌中表達水平較高,而在抗自溶能力較弱的啤酒酵母菌中表達水平較低[8]。
自溶過程中,細胞逐漸喪失生存能力,電子運輸、戊糖-磷酸鹽途徑、脂肪酸氧化、糖原分解代謝、海藻糖合成代謝、呼吸代謝等大部分能量的產生和利用也都受到干擾[24]。在之前的研究中,我們發現25個基因在酵母自溶過程中對細胞的拯救、防御和毒性起作用,其中12個基因下調,13個基因上調。下調和上調的基因均參與了抗氧化應激、滲透壓、化學刺激、熱休克、DNA損傷、pH和饑餓等應激反應[8]。在酵母自溶過程中,、、、、等基因被發現可能有重要的作用[2]。CWI通路是負責細胞壁組織和生物合成的主要系統,在細胞壁應激反應時,轉錄因子Rlm1被招募到和基因的啟動子上,激活這兩個基因啟動細胞的轉錄重編程[4,25-26]。基因的敲除可能導致產生了細胞壁缺陷從而引起了H-Δ細胞對米卡芬凈及熒光增白劑抗性的減弱。該基因作為參與CWI激活自調節反饋環的重要基因[25],其破壞導致反饋環的破壞,進而影響在細胞壁損傷應力下的表達。啤酒酵母在自溶過程中會迅速喪失生存能力,同時激活一系列應激反應對細胞進行改造以適應環境[1]。前期研究中從兩個不同自溶階段的啤酒酵母菌株的蛋白質二維凝膠電泳譜可以清楚地發現,在自溶過程中發現了蛋白質的大規模降解以及蛋白質數量的下降,同時也出現了一些新的蛋白質[2],結合轉錄組學數據我們推測啤酒酵母細胞對其基因組進行了大量的重編程以響應環境變化[8]。酵母細胞作為一種動態變化的結構,在生長和發酵過程中通過氧化劑應激、滲透應激、乙醇應激、饑餓應激等多種應激反應系統來調節細胞壁的組成和結構[1,7,23]。細胞壁完整性通路調節酵母細胞使其在惡劣的環境中能夠遵循嚴格的規律來維持其細胞結構的完整。
酵母在釀造工業中應用廣泛,啤酒酵母更是被認為是啤酒發酵的核心。對啤酒酵母的基因組測序工作證實了其雜交的本質[11,27-28],由于長期的種內及種間的雜交,啤酒酵母的倍性出現了非整倍體等特征,直接對其進行遺傳改造很難清楚地了解某個基因的功能。本研究所采用的啤酒酵母單倍體菌株H5是從中國啤酒廠廣泛使用的lager型啤酒酵母菌株中選育而來的,具有一定的代表性。在單倍體啤酒酵母菌株中研究基因的功能,能夠直觀地了解該基因在啤酒酵母菌中的功能及基因調控對啤酒釀造的影響。基因修飾雖然不影響菌株的生長和發酵性能,但仍然影響酵母菌株的應激反應,導致啤酒發酵結束后菌株細胞形態差異較大。啤酒酵母細胞在發酵過程中經歷了多種壓力脅迫交替出現的歷程,如高滲透壓脅迫、乙醇脅迫、營養消耗、罐體壓力等[1,7],所有這些壓力都會導致細胞的損傷和老化。基因的缺失可能加速了衰老過程,發酵過程結束后也發現了胞內細胞器的損傷。研究發現細胞壁組裝及DNA損傷應答相關基因的表達隨的過表達與敲除而調整,而CWI途徑中其他相關基因的調控方式并沒有明顯的規律,推測可能主要影響了CWI途徑中基因的表達,進而提高啤酒酵母在惡劣環境中的抗逆性。
本研究基于同源重組的方法對進行敲除和過表達,獲得了重組菌H-Δ和H/。研究了重組菌在啤酒發酵過程中的抗自溶性能,發現敲除影響了酵母的抗自溶能力。敲除了使啤酒酵母對細胞壁損傷、氮饑餓、滲透壓的耐受性以及溫度的敏感性受到影響[14]。結合qRT-PCR發現CWI途徑中與涉及細胞壁組裝和DNA損傷應答的在過表達時顯著上調。對基因的調控能夠影響酵母的抗自溶能力,為選育抗自溶能力高的啤酒酵母菌株提供參考依據。
[1] Gibson BR, Lawrence SJ, Leclaire JP, et al. Yeast responses to stresses associated with industrial brewery handling. Fems Microbiol Rev, 2007, 31(5): 535–569.
[2] Xu WN, Wang JJ, Li Q. Comparative proteome and transcriptome analysis of lager brewer’s yeast in the autolysis process. FEMS Yeast Res, 2014, 14(8): 1273–1285.
[3] Rodicio R, Heinisch JJ. Together we are strong-cell wall integrity sensors in yeasts. Yeast, 2010, 27(8): 531–540.
[4] Sanz AB, Garcia R, Rodriguez-Pe?a JM, et al. Cooperation between SAGA and SWI/SNF complexes is required for efficient transcriptional responses regulated by the yeast MAPK Slt2. Nucleic Acids Res, 2016, 44(15): 7159–7172.
[5] Watanabe Y, Irie K, Matsumoto K. Yeast rlm1 encodes a serum response factor-like protein that may function downstream of the Mpk1 (slt2) mitogen-activated protein kinase pathway. Mol Cell Biol, 1995, 15(10): 5740–5749.
[6] García R, Bermejo C, Grau C, et al. The global transcriptional response to transient cell wall damage inand its regulation by the cell integrity signaling pathway. J Biol Chem, 2004, 279(15): 15183–15195.
[7] Gibson BR, Lawrence SJ, Boulton CA, et al. The oxidative stress response of a lager brewing yeast strain during industrial propagation and fermentation. FEMS Yeast Res, 2008, 8(4): 574–585.
[8] Xu WN, Wang JJ, Li Q. Microarray studies on lager brewer's yeasts reveal cell status in the process of autolysis. FEMS Yeast Res, 2014, 14(5): 714–728.
[9] Li XE, Wang JJ, Li Q. Screening and RT-PCR analysis of stress resistant brewing yeast. Food Ferment Ind, 2014, 40(8): 18–23 (in Chinese). 李欣兒, 王金晶, 李崎. 壓力耐受啤酒酵母的篩選及其RT-PCR研究. 食品與發酵工業, 2014, 40(8): 18–23.
[10] Wendland J. Lager yeast comes of age. Eukaryot Cell, 2014, 13(10): 1256–1265.
[11] Nakao Y, Kanamori T, Itoh T, et al. Genome sequence of the lager brewing yeast, an interspecies hybrid. DNA Res, 2009, 16(2): 115–129.
[12] Xu WN. Study on the evaluation and mechanism of autolysis in lager brewer’s yeast[D]. Wuxi: Jiangnan University, 2014 (in Chinese).許維娜啤酒酵母自溶評價及機理研究[D]. 無錫: 江南大學, 2014.
[13] Ding HJ. Physiological and autolysis properties of brewer's yeast during passage [D]. Wuxi: Jiangnan University, 2018 (in Chinese).丁華建. 啤酒酵母在傳代過程中的生理與自溶性能研究[D]. 無錫: 江南大學, 2018.
[14] Ding HJ, Duan HX, Wang JJ, et al. Comparison of physiological performance differences between two lager yeasts during serial re-pitching and autolysis. Food Ferment Ind, 2018, 44(10): 57–64 (in Chinese). 丁華建, 段鴻緒, 王金晶, 等. 兩株lager啤酒酵母在傳代及自溶過程中生理性能差異的比較. 食品與發酵工業, 2018, 44(10): 57–64.
[15] Martínez-Rodríguez AJ, Polo MC, Carrascosa AV. Structural and ultrastructural changes in yeast cells during autolysis in a model wine system and in sparkling wines. Int J Food Microbiol, 2001, 71(1): 45–51.
[16] Dodou E, Treisman R. Themads-box transcription factor Rlm1 is a target for the Mpk1 mitogen-activated protein kinase pathway. Mol Cell Biol, 1997, 17(4): 1848–1859.
[17] Orlean P. Architecture and biosynthesis of thecell wall. Genetics, 2012, 192(3): 775–818.
[18] Venters BJ, Wachi S, Mavrich TN, et al. A comprehensive genomic binding map of gene and chromatin regulatory proteins in. Mol Cell, 2011, 41(4): 480–492.
[19] Nuoffer C, Jen? P, Conzelmann A, et al. Determinants for glycophospholipid anchoring of theGAS1 protein to the plasma membrane. Mol Cell Biol, 1991, 11(1): 27–37.
[20] Eustice M, Pillus L. Unexpected function of the glucanosyltransferase Gas1 in the DNA damage response linked to histone H3 acetyltransferases in. Genetics, 2014, 196(4): 1029–1039.
[21] Niepel M, Molloy KR, Williams R, et al. The nuclear basket proteins Mlp1p and Mlp2p are part of a dynamic interactome including Esc1p and the proteasome. Mol Biol Cell, 2013, 24(24): 3920–3938.
[22] Kim KY, Truman AW, Caesar S, et al. Yeast Mpk1 cell wall integrity mitogen-activated protein kinase regulates nucleocytoplasmic shuttling of the Swi6 transcriptional regulator. Mol Biol Cell, 2010, 21(9): 1609–1619.
[23] Levin DE. Regulation of cell wall biogenesis in: The cell wall integrity signaling pathway. Genetics, 2011, 189(4): 1145–1175.
[24] Wang JJ, Li MQ, Zheng FY, et al. Cell wall polysaccharides: Before and after autolysis of brewer’s yeast. World J Microb Biot, 2018, 34: 137.
[25] Monerawela C, Bond U. Brewing up a storm: The genomes of lager yeasts and how they evolved. Biotechnol Adv, 2017, 35(4): 512–519.
[26] Baker E, Wang B, Bellora N, et al. The genome sequence ofand the domestication of lager-brewing yeasts. Mol Biol Evol, 2015, 32(11): 2818–2831.
[27] Ishihara S, Hirata A, Nogami S, et al. Homologous subunits of 1,3-beta-glucan synthase are important for spore wall assembly in. Eukaryot Cell, 2007, 6(2): 143–156.
[28] Martin-Yken H, Dagkessamanskaia A, Basmaji F, et al. The interaction of Slt2 MAP kinase with Knr4 is necessary for signalling through the cell wall integrity pathway in. Mol Microbiol, 2003, 49(1): 23–35.
Regulations ofgene affect the anti-autolytic ability of lager yeast
Jinjing Wang1,2, Mengqi Li1,2, Dan Hou1,2, Weina Xu1,2, Feiyun Zheng1,2, Chunfeng Liu1,2, Chengtuo Niu1,2, and Qi Li1,2
1 Key Laboratory of Industrial Biotechnology, Ministry of Education, School of Biotechnology, Jiangnan University, Wuxi 214122, Jiangsu, China 2 Laboratory of Brewing Science and Technology, Jiangnan University, Wuxi 214122, Jiangsu, China
The autolysis of brewer’s yeast seriously affects the quality of beer and the quality of yeast is considered as one of the key factors in beer brewing. Previous studies on brewer’s yeast autolysis showed thatgene, an important transcription factor in cell integrity pathway, is closely related to the autolysis of yeast. In this study,was knocked out and overexpressed in a haploid brewer’s yeast.disruption resulted in poor anti-autolysis performance of yeast, whereas overexpression ofcontributed to the anti-autolytic ability of yeast. In addition,gene knockout affected the osmotic stress resistance, cell wall damage resistance, nitrogen starvation resistance and temperature tolerance of yeast strain. The transcriptional level ofinvolved in cell wall assembly and DNA damage response was regulated along with the expression of, whereas other genes in CWI pathway did not show apparent regularity.might mainly affect the expression ofso as to improve the stress resistance of lager yeast in harsh environment. The result from this study help further understand the mechanism of yeast autolysis and lay a foundation for breeding brewer’s yeast strain with better anti-autolytic ability.
lager yeast, anti-autolytic ability,, cell wall integrity
December 24, 2018;
February 28, 2019
National Natural Science Foundation of China (Nos. 31771963, 31571942).
Jinjing Wang. Tel/Fax: +86-510-85918176; E-mail: jjwang@jiangnan.edu.cn
國家自然科學基金 (Nos. 31771963,31571942) 資助。
2019-03-21
http://kns.cnki.net/kcms/detail/11.1998.Q.20190321.1053.001.html
10.13345/j.cjb.180530
王金晶, 李夢琦, 侯丹, 等. Lager啤酒酵母基因調控對其抗自溶性能的影響. 生物工程學報, 2019, 35(6): 1059–1070.
Wang JJ, Li MQ, Hou D, et al. Regulations ofgene affect the anti-autolytic ability of lager yeast. Chin J Biotech, 2019, 35(6): 1059–1070.
(本文責編 郝麗芳)
