基于雙向溫度梯度系統(tǒng)的檳榔萌發(fā)及生理響應(yīng)
2019-07-09 11:37:20黃麗云李東霞陳君劉立云周煥起
熱帶作物學(xué)報 2019年8期
黃麗云 李東霞 陳君 劉立云 周煥起


摘 ?要 ?本研究以‘熱研1號檳榔種子為供試材料,借助雙向溫度梯度系統(tǒng)控制面板設(shè)置49個晝夜溫度處理(最高溫區(qū)48?℃,最低溫區(qū)20?℃),研究其對種子萌發(fā)率、生長狀況及生理響應(yīng)的影響。結(jié)果表明:檳榔種子萌發(fā)最佳晝夜溫度組合為29/25?℃,萌發(fā)率100%,生長態(tài)勢最佳;20/20?℃條件下檳榔種子有萌發(fā),但發(fā)芽率為80%,種子長勢一般;48/48?℃條件下種子不萌發(fā)。根據(jù)長勢狀況,以及酶活(SOD、POD、GR)測定,明確了最適于種子萌發(fā)及生長的溫度范圍為25/20~34/25?℃。另外種子萌發(fā)所需日積溫560~744?℃?h為宜,萌發(fā)總積溫不宜超過24 864?℃?h。實驗還證實了檳榔種子變溫處理比常溫萌發(fā)效果更好。
關(guān)鍵詞 ?雙向溫度梯度系統(tǒng);檳榔;萌發(fā);生理生化
中圖分類號 ?S543.2 ?????文獻(xiàn)標(biāo)識碼 ?A
Abstract ?The effect of germination rate, growth conditions, physiological response of Arecanut was studied using the 2-way grants thermogradient plate which allows 49 temperature combinations (regimes) (20?℃ to 48?℃) of single temperature and alternating temperature combination over a time continuum. Under the best temperature combination 29/25?℃, the seed germination rate was 100% and the growth potential was the best. When temperature was 20/20?℃, the germination rate was 80%, and the growth was general. The 48/48?℃ treatment did not have any effect. According to the germination rate, growth conditions and physiological response (SOD, POD, GR), the most suitable temperature range was 25/2034/25?℃. In addition, the optimum reaction accumulated temperature was 560744?℃?h, and the total germination temperature should not exceed 24?864?℃?h. The experiment also supported that the alternate temperature treatment was better than the normal temperature treatment for the seed germination rate and growth of Arecanut.
Keywords ?2-way grants thermogradient plate; areca; germination; physiological response
DOI ?10.3969/j.issn.1000-2561.2019.08.007
檳榔(Areca catechu L.)是棕櫚科檳榔屬多年生經(jīng)濟(jì)作物,目前已成為海南第一大農(nóng)業(yè)支柱產(chǎn)業(yè)。檳榔是我國四大南藥之一,種子、果皮、花等均可入藥,檳榔中含有的檳榔堿、兒茶素、多糖、膽堿等具有殺蟲、消積、下氣、行水等功效。另外,檳榔嫩果(授粉受精后140~160 d采摘的未成熟果)作為咀嚼嗜好品供不應(yīng)求[1],目前全球約有10億人咀嚼檳榔,是檳榔的主要消費方式。
不同種類植物種子的萌發(fā)溫度要求不同,這是由植物本身特性所決定,也是植物長期適應(yīng)環(huán)境的結(jié)果。檳榔屬頑拗性種子,具有對溫度、干旱等外界環(huán)境較為敏感的特性。檳榔生長于熱帶及南北回歸線以內(nèi)的亞熱帶邊緣地區(qū),種果成熟月份集中在4—5月,恰逢海南高溫季節(jié),如何控制合適的溫度進(jìn)行種果萌發(fā)處理,避免因溫度過高造成大批量種果失活是最為關(guān)鍵的一步。由于種植的局域性與研究滯后的現(xiàn)狀,針對檳榔種果萌發(fā)的研究報道較少。董志國等[2]研究了不同濃度NaCl脅迫對檳榔種果發(fā)芽的影響;陳思婷等[3]實驗證明剝殼與切頭處理對檳榔種果發(fā)芽有顯著的促進(jìn)作用。但探討種果最適宜發(fā)芽溫度研究尚未見報道,本文利用雙向溫度梯度系統(tǒng),在廣泛的溫度范圍內(nèi)對種果進(jìn)行發(fā)芽測試,探討最佳發(fā)芽溫度及生長效應(yīng),比較不同溫度處理的生理生化差異,精準(zhǔn)確定檳榔種果的最佳發(fā)芽溫度,界定發(fā)芽的極限溫度,為檳榔規(guī)模化育苗提供理論依據(jù)。
1 ?材料與方法
1.1 ?材料
試材為中國熱帶農(nóng)業(yè)科學(xué)院椰子研究所國審品種‘熱研1號檳榔(熱品審2014011)[4]同串種果,種子均勻一致,無病蟲害。采回種果人工剝?nèi)ス猓孟礉嵕珜⒎N子上附著的果肉清洗干凈,然后用2.5%次氯酸鈉溶液將種子滅菌5 min,再用滅菌純水將種子清洗3遍。實驗儀器為溫度梯度種子系統(tǒng)(GRD1 Gradient Plate 40674-A)。
1.2 ?方法
1.2.1 ?實驗設(shè)計 ?設(shè)置了7×7矩陣共49個溫度組合(20~48?℃),如表1所示,高溫端為48/48?℃(白天/黑夜溫度),低溫端20/20?℃,第1梯度在黑暗中由左向右移動,每8 h交替一次,第2次由前向后隨光而變(光照強(qiáng)度4000 lx),每16 h交替一次。每個實驗格放置15個種果(去皮洗凈),每個處理3次重復(fù)。
1.2.2 ?測定指標(biāo)及方法 ?生長指標(biāo)觀測:以觀察到種子胚根、胚芽伸出種皮并發(fā)育到一定程度視為發(fā)芽;定期觀測種子發(fā)芽率、胚芽、胚根及發(fā)
根數(shù)的生長動態(tài);發(fā)芽率=n/N×100%(式中,n為實驗結(jié)束后發(fā)芽粒數(shù);N為供試種子總數(shù));積溫T=TN?Nt+TD?Dt(式中,T為積溫值,TD、TN分別為晝周期和夜周期對應(yīng)的平均溫度;Dt、Nt為種子萌發(fā)對應(yīng)的晝夜小時數(shù))。生理生化指標(biāo)測定:取樣點為播種28 d后距離萌發(fā)孔基部0.8~1.5 cm的胚芽部位。蛋白(Pro)、超氧化物歧化酶(SOD)和過氧化物酶(POD)、谷胱甘肽還原酶(GR)等按照南京建成生物工程研究所提供的試劑盒進(jìn)行測定,SOD和POD酶活性按每毫克蛋白中的酶活力單位數(shù)計算(U/mg)。每個指標(biāo)重復(fù)測定3次取平均值。
1.3 ?數(shù)據(jù)處理
采用SPSS 19.0軟件與office Excel軟件對數(shù)據(jù)進(jìn)行統(tǒng)計分析。
2 ?結(jié)果與分析
2.1 ?溫度處理對種子萌發(fā)的影響
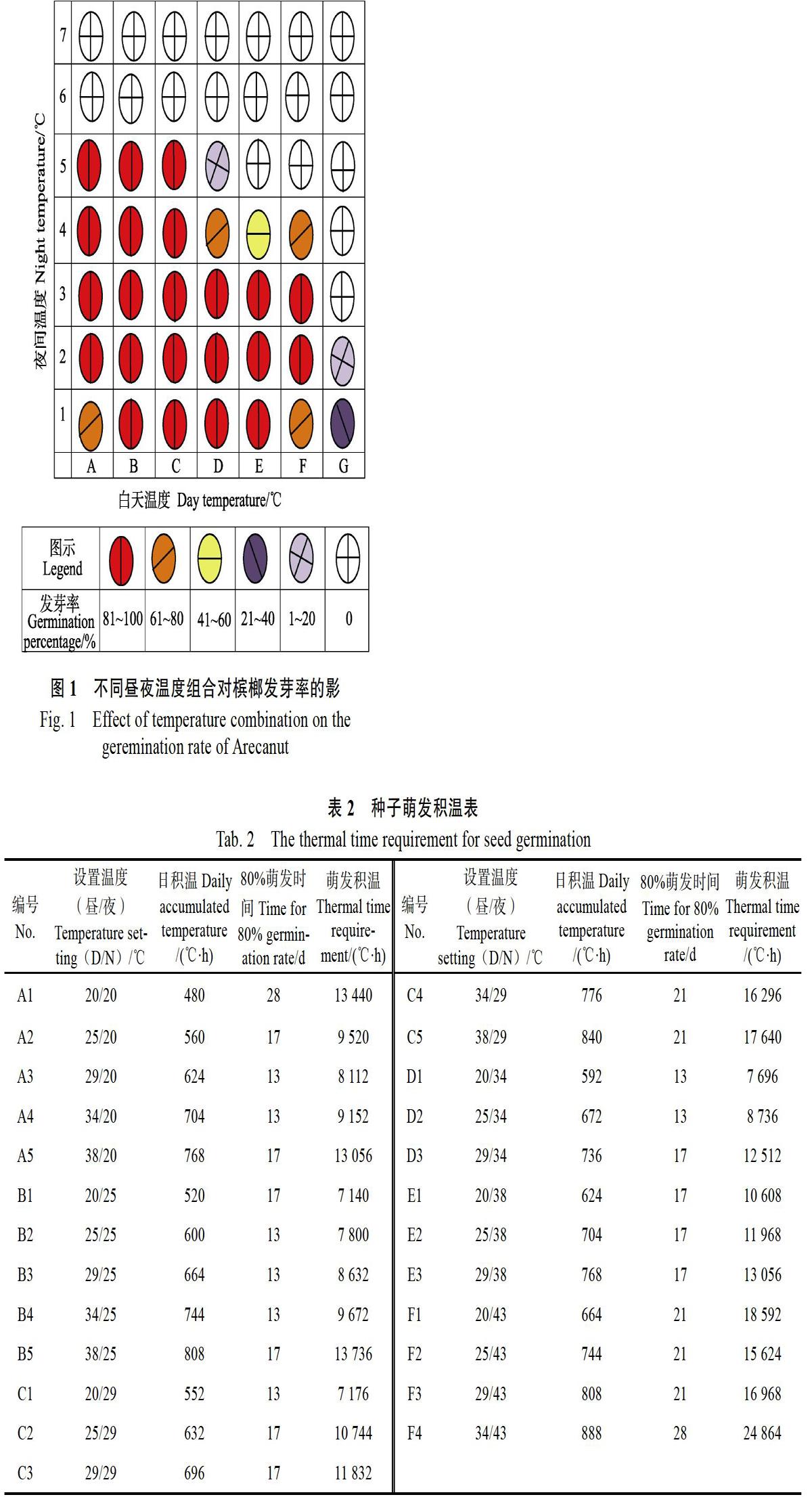
2018年4月20日將種子置于溫度板上,4月28日開始萌發(fā),最早是B2、B3、B4、C2、C3等溫度組合出現(xiàn)萌發(fā)現(xiàn)象。種子發(fā)芽周期約28 d(5月18日)。結(jié)合表1,由圖1可見,交替溫度條件下,在25~43?℃(夜)和25~38?℃(日)范圍內(nèi)種子萌發(fā)速度最快,效果好,發(fā)芽率達(dá)100%。另外,可萌發(fā)的邊緣區(qū)域溫度組合為D5、F4、G1、G2發(fā)芽率分別為93.3%、80%、33.3%、20%。恒溫條件下,A1(20/20?℃)、B2(25/25?℃)、C3(29/29?℃)、D4(34/34?℃)、E5(38/38?℃)、F6(43/43?℃)、G7(48/48?℃)處于面板中軸斜線的位置,發(fā)芽率分別為80%、100%、100%、66.7%、0、0、0,發(fā)芽率呈“低-高-低”的拋物線形。
2.2 ?積溫對種子萌發(fā)的影響
從表2可見,49個溫度組合中25個萌發(fā)率達(dá)80%以上,占比51%。其中A3、A4、B2、B3、B4、C1、D1、D2共8個溫度組合種子萌發(fā)率達(dá)80%僅需13 d,日積溫范圍為552~744?℃?h,總萌發(fā)積溫范圍為7176~9672?℃?h。17 d時種子萌發(fā)率達(dá)80%的有10個溫度組合,日積溫范圍為520~808?℃?h,總萌發(fā)積溫范圍為7 140~13 736?℃?h,涵蓋了13 d萌發(fā)率達(dá)80%的日積溫及總積溫范圍。在28 d方可達(dá)到80%發(fā)芽率的溫度組合20/20、34/43?℃中,日積溫分別為480、888?℃?h,是處理組合達(dá)80%以上的最低值和最高值。
2.3 ?溫度處理對種子生長勢的影響
從圖1可見,檳榔種子發(fā)芽溫度范圍較廣,25~38?℃均可正常發(fā)芽,但在相同發(fā)芽率情況下種子生長狀況差異較大。如圖2所示,檳榔長勢最佳溫區(qū)為A2~A4、B2~B4、C2~C3、D1~D3、E1~E2;其次為A1、B1、C1、E3、F1;A6~G6、A7~G7、G1~G7為不萌發(fā)狀態(tài)。根據(jù)種子萌發(fā)生長的表征,選取發(fā)芽率為100%的3個溫度處理B3(29/25?℃)、C1(20/29?℃)、D4(34/34?℃),在不同的發(fā)育時期(5月8日,5月18日,5月28日)進(jìn)行測量。從圖3、圖4和圖5可見,無論是胚芽、胚根和發(fā)根數(shù),B3處理均顯著高于C1和D4,生長態(tài)勢由高至低為B3(29/25?℃)>C1(20/29?℃)>D4(34/ 34?℃),由此可見,合適的積溫條件下,相同發(fā)芽率情況下,合適的積溫條件種子長勢有顯著差異。
2.4 ?種子萌發(fā)的生理生化響應(yīng)
以B3、C1、D4代表3個不同生長勢區(qū)間材料,開展其生理生化研究。結(jié)果如圖6、圖7和圖8所示。不同溫度組合條件下對種子萌發(fā)過程中SOD、POD、GR活性影響不同,3種溫度組合種子萌發(fā)過程中3種酶活性規(guī)律一致,由高到低為B3(29/25?℃)>C1(20/29?℃)>D4(34/34?℃)。其中抗氧化物酶SOD、POD活性在三者之間差異達(dá)顯著水平(P<0.05)。B3處理的SOD值為80 U/mg,比C1和D4處理顯著高出40.79%和79.70%;POD的表現(xiàn)變化與SOD的較為一致,亦是B3處理POD值最大,比C1與D4分別高出60.04%和82.78%。
3 ?討論
本研究所用儀器雙向溫度梯度系統(tǒng)是高效性的溫度循環(huán)系統(tǒng),是在鋁板平面一側(cè)加熱同時另一側(cè)制冷,從而在板面形成溫度梯度,通過梯度板再次旋轉(zhuǎn)90°,即可形成多孔溫差條件。該系統(tǒng)可同時模擬196個不同的溫差條件,可快速模擬和摸索培養(yǎng)最佳溫度條件,一次性得出大量實驗數(shù)據(jù),替代了光照培養(yǎng)箱溫度條件單一的缺陷。特別是用于探索種植地域優(yōu)化篩選、種子最佳溫度條件、種子活性鑒定等效果顯著,可在保證準(zhǔn)確性的條件下節(jié)省大量時間和實驗測試,Joseph 等[5]和Asomaning等[6]均運用該儀器進(jìn)行了艷麗欖仁、白卡雅楝種子的變溫處理實驗。
種子萌發(fā)溫度具有最高溫與最低溫的范圍需求,超出范圍都會使酶失去催化能力,從而使種子失去萌發(fā)力。種子萌發(fā)是植物生長周期的重要階段,此期間萌發(fā)生長極易受溫度等環(huán)境的影 ?響[7]。理想條件下,種子生長勢旺盛,酶活力強(qiáng),催化能力亦強(qiáng)。本研究中最低溫設(shè)置為20?℃,是因為檳榔為忌低溫的敏感性種子,加之檳榔發(fā)芽季平均溫度為25~30?℃;最高溫設(shè)置為48?℃,筆者在種果檳榔采摘后進(jìn)行裝袋堆漚(一般海南農(nóng)戶的種子處理方法),測定袋內(nèi)溫度,可高達(dá)50?℃左右。溫度高的原因主要是種子的呼吸作用、氣候因素以及微生物發(fā)酵升溫等,當(dāng)種子貯藏置放不當(dāng)極易引起高溫?fù)p傷[8]。
在種子萌發(fā)方面,20?℃常溫處理檳榔種子有萌發(fā),但發(fā)芽率為80%,種子長勢一般;50?℃處理種子萌發(fā)率為0;極限高溫處理組合為34/43?℃(日均溫為37?℃),發(fā)芽率為66.7%,種子長勢差;最適于種子萌發(fā)及生長的溫度范圍為25/20~34/25?℃(日均溫為23.3~31?℃),最佳的溫度組合為29/25?℃,這與棕櫚種子萌發(fā)溫度(30/20?℃)較為接近[9]。由此計算出最佳萌發(fā)日積溫范圍為560~744?℃?h,總萌發(fā)積溫為7280~9672?℃?h。
另外本研究還發(fā)現(xiàn),檳榔種子溫度交替變化處理比常溫條件發(fā)芽效果好。實驗中29/29和34/34?℃日積溫與25/34和38/25?℃相似,但80%種子萌發(fā)率時間與生長勢明顯差別。類似的研究多見報道,恒溫不利于羊草種子發(fā)芽[10],變溫能顯著提高種子萌發(fā)率[11-14],這可能是棕櫚科大多數(shù)植物長期適應(yīng)原生境的結(jié)果。關(guān)于變溫促進(jìn)種子發(fā)芽的機(jī)理目前尚未認(rèn)識清楚,據(jù)分析可能與不同溫度滿足種子萌發(fā)不同生理過程有關(guān)。
POD、SOD和GR是植物體內(nèi)清除自由基的重要保護(hù)酶。通常情況下,正常植株體內(nèi)保護(hù)酶都具有較高的活性,可有效地清除細(xì)胞內(nèi)的各種活性氧自由基。本研究中,SOD、POD、GR等酶協(xié)調(diào)一致,維持在一個相對高的穩(wěn)定水平,從而防止生物自由基對細(xì)胞的毒害。本研究中種子萌發(fā)長勢越好,酶活越強(qiáng),反映了長勢旺盛的種苗具有良好的防御系統(tǒng)。另外溫梯度板的種子培養(yǎng)屬于微逆境環(huán)境,無濕度控制,靠人為手動控濕,種子萌發(fā)相關(guān)酶活性也會隨著環(huán)境條件而變化,曹紅星等[15]、孫程旭等[16]、黃麗云等[17]和李佳等[18]以檳榔幼苗為試材開展低溫、干旱、耐旱等抗逆性研究,表明在逆境條件下,隨著脅迫程度的增加,抗氧化物酶活性均有不同程度的提高。本研究種子萌發(fā)過程會受濕度變化的影響,導(dǎo)致酶活變化,反映出不同溫度處理種子自身生長調(diào)控的能力。
本研究探討了溫度對檳榔種子萌發(fā)的影響,明確了檳榔種子的最佳晝夜溫度組合為29/25?℃,最佳萌發(fā)恒溫為23.3~31.0?℃,日積溫560~744?℃?h,萌發(fā)積溫不宜超過24 864?℃?h。另外證實了檳榔種子變溫交替處理比常溫處理萌發(fā)效果好。
參考文獻(xiàn)
黃麗云, 劉立云, 齊 ?蘭, 等. 基于Logistic模型的檳榔果實生長發(fā)育研究[J]. 熱帶農(nóng)業(yè)科學(xué), 2018, 38(8): 105-108.
董志國, 劉立云. NaCl脅迫對檳榔種果發(fā)芽的影響[J]. 安徽農(nóng)業(yè)學(xué)報, 2008, 14(22): 105-106.
陳思婷, 王 ?萍. 剝殼與切頭對檳榔種子發(fā)芽的影響[J]. 熱帶農(nóng)業(yè)科學(xué), 2011, 31(10): 13-15.
李和帥, 范海闊, 黃麗云, 等. 檳榔新品種“熱研1號”[J]. 園藝學(xué)報, 2011, 38(5): 1011-1012.
Joseph M A, Moctar S, Nana S O. Germination responses of Terminalia superba Engl and Diels seeds on the 2-way grants thermogradient plate[J]. Research Journal of Seed Science, 2011, 4(1): 28-39.
Asomaning J M, Sacande M, Olympio N S. Germination responses of Khaya Anthotheca seeds to a range of temperatures provided by the 2-way grants thermogradient plate[J]. Ghana Journal Forestry, 2010, 26: 74-88.
張曉艷, 楊忠仁, 郝麗珍, 等. 溫度及鹽脅迫對地梢瓜種子萌發(fā)及抗氧化酶活性的影響[J]. 西北植物學(xué)報, 2017, 37(6): 1166-1174.
畢辛華, 戴心維. 種子學(xué)[M]. 北京: 中國農(nóng)業(yè)出版社, 2011: 86-87.
唐安軍, 龍春林, 刀志靈, 等. 棕櫚種子萌發(fā)特性及其貯藏行為的研究[J]. 云南植物研究, 2005, 27(6): 657-662.
楊偉光, 劉盼盼, 袁光孝, 等. 種質(zhì)及變溫對羊草種子萌發(fā)的影響[J]. 草業(yè)學(xué)報, 2018, 27(7): 103-111.
易 ?津, 李青豐, 田瑞華. 賴草屬牧草種子休眠與植物激素調(diào)控[J]. 草地學(xué)報, 1997, 5(2): 93-100.
藺吉祥, 邵 ?帥, 隋 ?丹, 等. 幾種提高羊草種子發(fā)芽率方法的比較[J]. 中國草地學(xué)報, 2014, 36(3): 47-51.
Gu A L, Yi J, Roman H, et a1. Effects of low-temperatures on seed germination of Leymus chinensis and Pascopyrum smithii[J]. Chinese Journal Grassland, 2005, 27(2): 50-54.
Chiwocha S D S, Cutler A J, Abrams S R, et al. The etrl-2 mutation in Arabidopsis thaliana affects the abscisic acid, auxin, cytokinin and gibberellin metabolic pathways during maintenance of seed dormancy, moist-chilling and germination[J]. The Plant Journal, 2005, 42(1): 35-48.
曹紅星, 馮美利, 孫程旭, 等. 低溫及干旱脅迫對檳榔幼苗生理生化特性的影響[J]. 西南農(nóng)業(yè)學(xué)報, 2010, 23(3): 832-335.
孫程旭, 陳良秋, 馮美利, 等. 檳榔不同品種幼苗耐寒性比較初步研究[J]. 西南農(nóng)業(yè)學(xué)報, 2009, 22(6): 1686-1689.
黃麗云, 劉立云, 李 ?艷, 等. 低溫脅迫對“熱研1號”檳榔新品種生理特性的影響[J]. 熱帶作物學(xué)報, 2015, 36(11): 2015-2018.
李 ?佳, 劉立云, 李 ?艷, 等. 保水劑對干旱脅迫檳榔幼苗生理特征的影響[J]. 南方農(nóng)業(yè)學(xué)報, 2018, 49(1): 104-108.
