基于轉錄-代謝聯合分析哈茨木霉ACCC32527對NaCl脅迫的分子調節
2019-07-06 03:00:50向杰陳敬師夏鑫鑫劉快李世貴顧金剛
中國農業科學 2019年12期
關鍵詞:差異
向杰,陳敬師,夏鑫鑫,劉快,李世貴,顧金剛
(中國農業科學院農業資源與農業區劃研究所農業農村部農業微生物資源收集保藏重點實驗室,北京100081)
0 引言
【研究意義】土壤鹽漬化是由自然因素或人類活動引起的環境風險之一,是世界范圍內農業發展所面臨的嚴重威脅。土壤過度鹽漬化不僅抑制土壤有機質的積累,改變土壤滲透勢和基質潛力,土壤團聚體減少[1],還可使植物吸水困難,導致作物生長不良,病蟲害發生加劇,嚴重阻礙農業生產[2]。研究表明,通過耐鹽堿微生物作用可改善植物根際環境,減輕鹽分對作物生長的抑制作用[3-5]。木霉菌(Trichodermaspp.)是一類能有效抑制植物病害的生防真菌,部分木霉菌株可定殖于植物根際形成共生體,一定程度上增加植物根系養分吸收利用率,促進植物生長,提高農作物產量并誘導植物增強抗逆性[6]。RAWAT等[7]從分離得到的54株Trichodermaspp.中發現,耐鹽能力較強的木霉菌具有更快的產孢速度和更強的拮抗尖鐮孢(Fusarium oxysporum)能力;HASHEM等[8]研究發現,接種鉤狀木霉(T. hamatum)能顯著降低NaCl脅迫下沙漠灌木槳果木犀草(Ochradenus baccatus)葉片中丙二醛含量,進一步提高部分抗氧化酶的活力。因此,研究木霉菌對鹽離子的耐受機理,對于提高木霉菌的耐鹽性、提高對植物病害的防治效果和改善植物根系的微生態環境具有重要意義。【前人研究進展】高濃度鹽離子脅迫下,微生物易產生滲透脅迫,并出現氧化損傷、細胞膜通透性改變、代謝活動停滯,菌落生長受到明顯抑制[9-11]。在長期的進化過程中,微生物也形成了一定的防御機制,主要包括離子轉運系統和胞內滲透調節系統等[12]。DURAN等[13]認為絲狀真菌(如Aspergillussp.)響應滲透脅迫的主要機制包括由MAPKs激活HOG(high osmolarity glycerol)通路合成甘油等滲透保護性物質、細胞骨架的重組和細胞壁再生等。哈茨木霉(T. harzianum)hog1是釀酒酵母(Saccharomyces cerevisiae)控制高滲應激反應的HOG1同源基因,沉默哈茨木霉hog1后,發現該菌對高滲環境的抵抗力降低,并對甜菜莖點霉(Phoma betae)和炭疽病菌(Colletotrichum acutatum)等植物病原菌拮抗能力下降[14]。超氧化物歧化酶(SOD)是一種能有效催化超氧化物陰離子降解的金屬蛋白酶,通過農桿菌介導法向哈茨木霉導入SOD相關基因可以提高木霉對高溫(40℃)和NaCl(2 mol·L-1)的耐受性[15]。棘孢木霉(T. asperellum)SOD相關編碼基因敲除后,突變株在1.2 mol·L-1NaCl脅迫下的滲透調節能力減弱,并對新月彎孢(Curvularia lunata)、擬輪枝鐮孢(Fusarium verticillioides)和立枯絲核菌(Rhizoctonia solani)的拮抗能力降低[16]。【本研究切入點】微生物的耐鹽機理是一個極其復雜的過程,僅通過部分抗鹽相關基因的克隆與功能研究不能更好地詮釋耐鹽機理。多層組學的發展使微生物逆境生理研究獲得更加豐富與完整的基因表達和代謝產物信息。木霉菌是生防領域中應用最廣泛的真菌之一,但其在鹽脅迫下的轉錄組和代謝組差異分析尚未見報道。本課題組前期自鹽堿地中篩選得到哈茨木霉菌株ACCC32527,該菌可在8% NaCl脅迫條件下生長,并對煙草黑脛病菌(Phytophthora parasiticavar.nicotianae)具有一定的防治效果[17]。【擬解決的關鍵問題】采用RNA-seq和GC-TOF-MS技術,對耐鹽哈茨木霉ACCC32527在0(T1)、0.4 mol·L-1(T2)、0.6 mol·L-1(T3)NaCl脅迫條件下的差異轉錄組和次級代謝產物進行分析,篩選與耐鹽機制調控相關的功能基因及次級代謝產物,為探討木霉菌的耐鹽機理提供依據。
1 材料與方法
試驗于2018年3—8月在中國農業微生物菌種保藏與管理重點實驗室完成。
1.1 材料
供試哈茨木霉菌株ACCC32527,保藏于中國農業微生物菌種保藏管理中心(ACCC)。
1.2 方法
1.2.1 NaCl脅迫處理 無菌水收集 PDA平板上ACCC32527孢子,并稀釋至1.0×106孢子/mL,按1%接種量分別接種于含 0、0.2、0.4、0.6 mol·L-1NaCl液體培養基(葡萄糖 40 g·L-1、蛋白胨 20 g·L-1、K2HPO4·3H2O 8 g·L-1、MgSO4·7H2O 2.5 g·L-1),28℃、180 r/min避光連續培養。培養4 d后,用8層紗布過濾,ddH2O洗滌收集菌絲,凍干并稱重。參照 DNS法[18]分別檢測培養結束后發酵液中葡萄糖剩余量,計算形成單位質量菌絲量所消耗的葡萄糖量,葡萄糖利用量(g/生物量)=(對照液葡萄糖含量-發酵液葡萄糖含量)/生物量。每組樣品3個生物學重復。
1.2.2 NaCl脅迫下ACCC32527轉錄差異分析 cDNA文庫構建及測序:收集的各組樣品委托北京百邁客生物科技有限公司進行后續處理,包括使用Trizol法提取樣本總RNA,EBNext?UltraTMRNA Library Prep Kit試劑盒完成 cDNA文庫構建,并使用 Illumina HiseqXTen高通量測序平臺對cDNA文庫進行測序。每組樣品3個生物學重復。
基因注釋和表達定量分析:使用 Illumina HiseqXTen測序平臺對各樣品cDNA文庫進行測序,得到原始數據(Raw Data),對Raw Data進行過濾,去除其中的接頭序列及低質量序列得到Clean Data。通過 Trinity軟件對 Clean Data進行組裝,獲得ACCC32527的unigene庫。使用BLAST軟件將unigene序列與NR、Swiss-Prot、GO、COG、KOG、eggNOG4.5和KEGG數據庫比對,使用KOBAS2.0得到unigene在 KEGG(Kyoto Encyclopedia of Genes and Genomes)中的KEGG Orthology分析結果,預測完unigene的氨基酸序列之后使用HMMER軟件與Pfam數據庫比對,獲得unigene的注釋信息。采用Bowtie將測序得到的序列與 unigene庫進行比對,根據比對結果,結合RSEM軟件進行表達量水平估計。利用FPKM(每百萬 Reads中來自比對到某一基因每千堿基長度的Reads數目)值表示對應unigene的表達豐度。
差異表達基因(DEG)篩選:T1、T2、T3樣品間DEG定量分析通過DEseq2完成。在篩選過程中,采用Benjamini-Hochberg方法對原有假設檢驗得到的顯著性P值(P-value)進行校正,并最終得到校正后的P值,即FDR(false discovery rate,錯誤發生率),作為DEG篩選的關鍵指標。將得到的FDR<0.01且差異倍數FC(fold change,兩樣品間表達量的比值)≥2作為篩選標準。
1.2.3 NaCl脅迫下ACCC32527差異代謝組分析 樣品準備:取凍干后菌體樣本(50±1)mg置于2 mL EP管中,加入450 μL提取液(甲醇∶氯仿=3∶1),再加入10 μL L-2-氯苯丙氨酸,渦旋30 s;加入瓷珠,45 Hz研磨儀處理4 min,超聲5 min(冰浴),重復3次;將樣本4℃,12 000 r/min離心15 min;移取350 μL上清液于1.5 mL EP管中;在真空濃縮器中干燥提取物;向干燥后的代謝物加入60 μL甲氧胺鹽試劑(甲氧胺鹽酸鹽溶于吡啶20 mg·mL-1),輕輕混勻后,放入烘箱中 80℃孵育 30 min;向每個樣品中加入 80 μL BSTFA(含有 1% TMCS,v/v),將混合物 70℃孵育1.5 h;準備上機檢測。每組樣品3個生物學重復。數據處理:使用LECO-Fiehn Rtx5數據庫對質譜數據進行匹配,采取將 student’st-test的P值和OPLS-DA模型的VIP值相結合的方法來篩選差異代謝物,篩選的標準為P-value≤0.05,VIP>1。
1.2.4 實時熒光定量 PCR(RT-qPCR) 為檢測轉錄組數據的準確性,從NaCl脅迫下DEG中選取7個差異表達顯著的基因,用Primer 5.0設計引物(表1)。使用PrimeScript 1st strand cDNA Synthesis Kit反轉錄試劑盒合成cDNA模板。以cDNA鏈為模板,UCE(泛素結合酶)為內參基因[19],使用 TB GreenTMPremix EX TaqTM熒光定量試劑盒、QuantStudioTMreal-time PCR軟件(Applied Biosystems公司)進行熒光定量檢測,采用-ΔΔCt法計算相對表達量,每個樣品3個生物學重復。

表1 RT-qPCR驗證基因及引物設計Table 1 Gene selection and primer design for RT-qPCR
2 結果
2.1 NaCl脅迫對ACCC32527生物量的影響
不同濃度NaCl脅迫對哈茨木霉ACCC32527生物量的影響如圖 1所示,0.2、0.4、0.6 mol·L-1NaCl脅迫對ACCC32527生長抑制率分別為5.66%、24.08%和 47.24%。其中,0.4、0.6 mol·L-1NaCl脅迫下,ACCC32527單位質量生物量消耗的葡萄糖分別為4.61 g和4.65 g,分別較對照提高32.84%和34.16%。
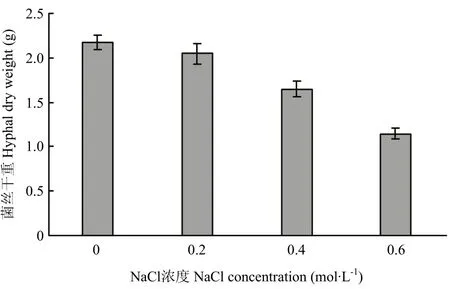
圖1 不同濃度NaCl脅迫對ACCC32527生長的影響Fig. 1 Effects of different concentrations of NaCl stress on the growth of ACCC32527
2.2 NaCl脅迫下ACCC32527差異轉錄組分析
2.2.1 差異基因共表達趨勢分析及GO功能注釋 經篩選,從T1 vs T2比較組中共獲得637個差異表達基因(DEG),包括 389個上調表達基因和248個下調表達基因;從T1 vs T3比較組中共獲得1 570個DEG,包括900個上調表達基因和670個下調表達基因。為觀察DEG在不同處理下各樣品表達量變化,根據log2(FPKM+1)值進行DEG共表達趨勢分析。趨勢分析結果顯示,1 921個DEG共聚類為16種基因表達模式,其中950個DEG共聚類為9種基因持續上調表達模式(圖2-A—C、2-E、2-G—H、2-J—L),662個DEG共聚類為3種基因持續下調表達模式(圖2-F、2-I、2-P),309個DEG共聚類為4種不規則表達模式(圖2-D、2-M—O)。
對16種表達模式的基因分別進行GO功能注釋,共包括分子功能、細胞組分和生物學進程3個基本分類和42個小類(圖2)。16種表達模式所包含基因共有的功能分類是催化活性、結合、細胞器、細胞膜部分、細胞膜、細胞部分、細胞、單組織過程和代謝過程,且占主要比例,約為61%—94%,說明這9種功能分類所涉及的基因主要響應 ACCC32527的脅迫應答過程。除此之外,每個趨勢圖基因還包含轉運活性和生物調節等幾個甚至幾十個功能小類。

圖2 差異表達基因GO分類Fig. 2 Go classification of DEGs
2.2.2 KEGG代謝通路分析 KEGG通路數據庫包含了各生物通路中分子相互作用網絡和生物體所特有的變化形式,從而確定蛋白質參與的最重要的信號轉導通路和生化代謝通路。T2、T3處理下ACCC32527差異表達基因KEGG通路分析表明,分別有225和535個差異基因富集于20條KEGG通路,主要包括真菌核糖體生物合成、嘌呤代謝、嘧啶代謝和剪切體等(圖3)。其中,分別有 12和 19個基因極顯著(CorrectP-valueT1vsT2=1.9×10-4,P-valueT1vsT3=1.6×10-5)富集于真菌核糖體生物合成途徑。T2處理下,該通路共有12個核糖體蛋白相關基因差異表達下調;T3處理下,共有DKC1、NHP2、UTP15等18個核糖體蛋白相關基因下調和CK2A蛋白相關編碼基因表達上調。
2.2.3 NaCl對ACCC32527抗逆相關基因的影響 與T1相比,T2或T3處理下ACCC32527大量氧化還原蛋白發生差異表達,包括 8個過氧化物酶、1個谷胱甘肽過氧化物酶、26個單加氧酶、5個過氧化氫酶和4個硫還原蛋白等,且多數為上調表達。另外,共發現有11個ABC轉運蛋白相關基因發生差異表達,其中 8個基因上調表達,3個基因下調表達。本研究檢測到1個編碼真菌classⅡ疏水蛋白基因(c60770.graph_c0)在T2和T3處理下表達量均發生上調。離子轉運方面,篩選得到 1個上調表達的Na+/K+-ATPase合成相關基因(圖4)。
2.3 代謝組分析
2.3.1 偏最小二乘法判別分析 為從各樣品中檢測并篩選出具有重要生物學意義和統計顯著差異的代謝物,并以此為基礎闡明NaCl脅迫下ACCC32527的代謝變化機制,分別對T1、T2、T3處理后樣品進行轉錄組學分析。基于GC-TOF-MS數據對T1、T2和T3各樣品數據進行偏最小二乘法判別分析(partial least square method-discrimination analysis,PLS-DA)。本試驗中R2X、R2Y>0.6,且Q2>0.3,T1和T3組內聚類效果較好,組間差異明顯,但T2處理下3個樣品散點分散,重復性較差,與T1、T3組間差距較小(圖5)。因此,將重點分析T1和T3的差異代謝產物。
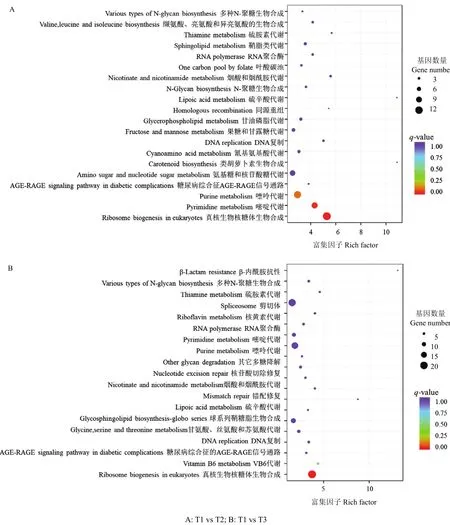
圖3 差異表達基因KEGG通路富集Fig. 3 Enriched KEGG terms of DEGs
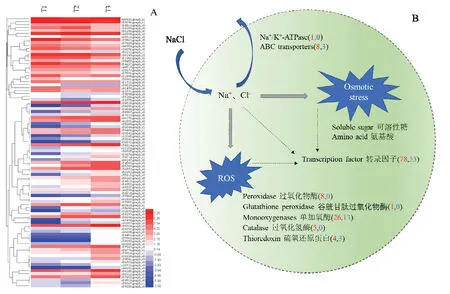
圖4 推測哈茨木霉ACCC32527耐鹽機理Fig. 4 Speculated mechanism of T. harzianum ACCC32527 resistance to NaCl
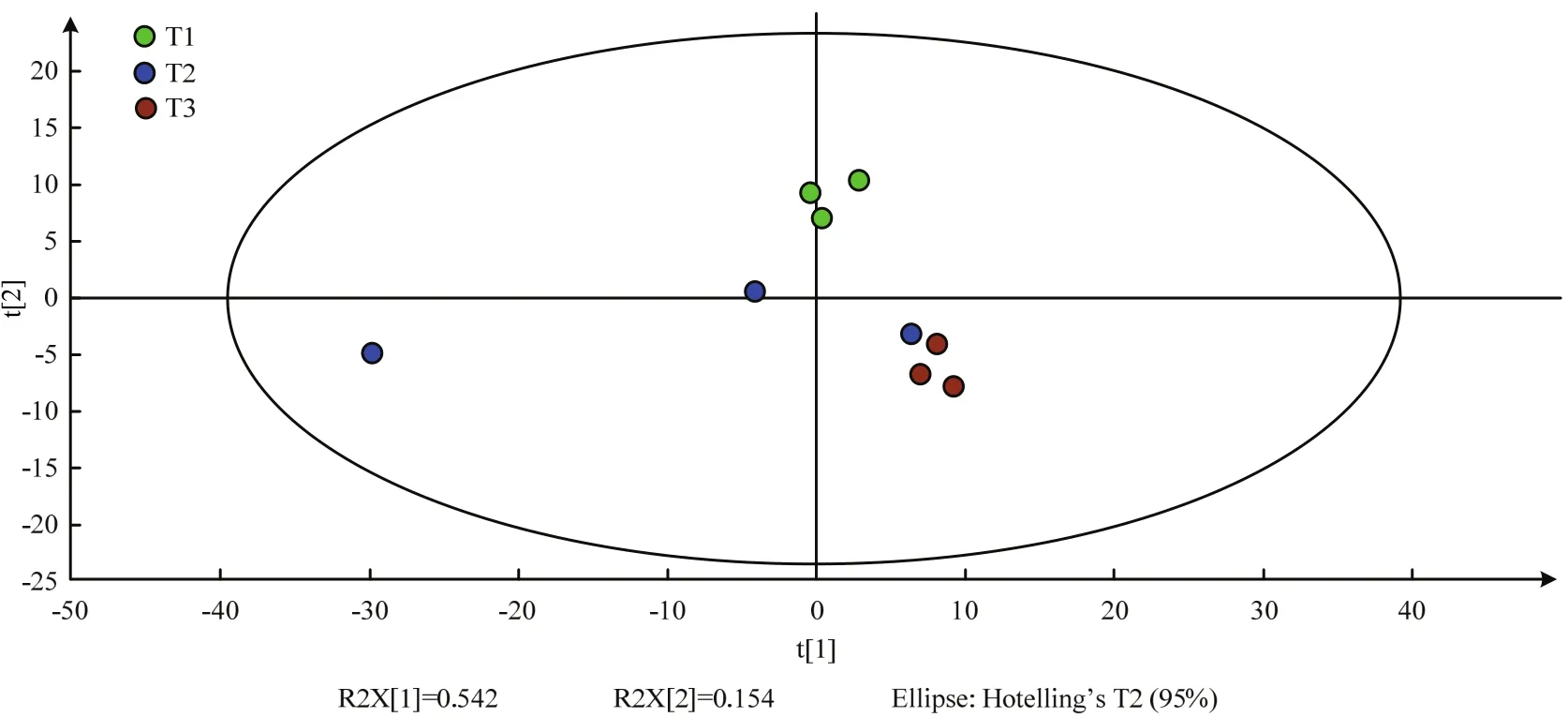
圖5 各樣本PLS-DA得分圖Fig. 5 PLS-DA score map of samples
2.3.2 差異代謝產物篩選及注釋 以 VIP>1,P-value≤0.05作為篩選標準,共得到336種代謝產物信息。與T1相比,T3處理后ACCC32527累積量增加的代謝產物有30種,減少的代謝產物有53種,而253個代謝產物含量保持不變。
在 83種差異物質中,只有 34種物質能夠在LECO-Fiehn Rtx5數據庫中找到匹配物質(表2),主要包括以下幾類:(1)氨基酸及其衍生物,如天冬氨酸和N-甲酰-L-蛋氨酸;(2)糖類及其衍生物,如赤蘚糖和葡萄糖-1-磷酸;(3)脂肪酸,如豆蔻酸和花生四烯酸;(4)有機酸,如檸檬酸和馬來酸;(5)醇類,如丙三醇和丁三醇;(6)核酸及其衍生物,如2-脫氧尿苷和尿嘧啶。鹽脅迫下,真菌普遍利用的滲透調節物質包括多元醇、糖類和部分氨基酸及其衍生物。在累積量上調的差異代謝物中,發現10種含量上調的氨基酸,包括L-組氨酸甜菜堿、酪氨酸和鳥氨酸等。甘油是絲狀真菌和酵母菌的重要滲透保護物質,T3處理下ACCC32527胞內甘油累積量發生上調(FC=1.504),維持胞內滲透平衡。

表2 哈茨木霉ACCC32527差異代謝產物Table 2 The differential metabolites of T. harzianum ACCC32527
2.3.3 差異代謝物KEGG通路富集分析 對T1和T3組的差異表達代謝物進行KEGG通路富集分析,主要涉及的通路包括代謝、遺傳信息過程和環境信息過程3類,包括代謝途徑、抗生素的合成、次級代謝產物的合成、ABC轉運蛋白和氨基酸的生物合成等40個通路發生變化。其中,有85%的通路中的差異代謝產物以累積量上調為主,下調的代謝產物集中在酪氨酸代謝和代謝通路(圖6)。
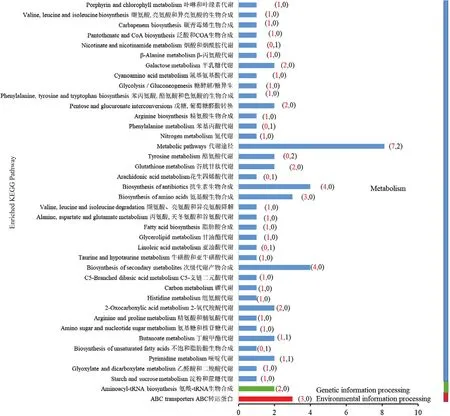
圖6 KEGG富集柱狀圖Fig. 6 Enriched column map of KEGG
2.4 RT-qPCR驗證
為驗證RNA-seq數據的準確性,從T3處理下的DEG中隨機選取7個差異表達顯著的基因,包括3個上調表達基因和4個下調表達基因,設計引物并利用RT-qPCR進行對照組和NaCl脅迫下基因的表達量變化分析。DEG的RT-qPCR結果與轉錄組測序的結果在基因表達幅度上有一定的差異,但基因的表達趨勢一致(圖7),說明轉錄組測序的結果是可信的。
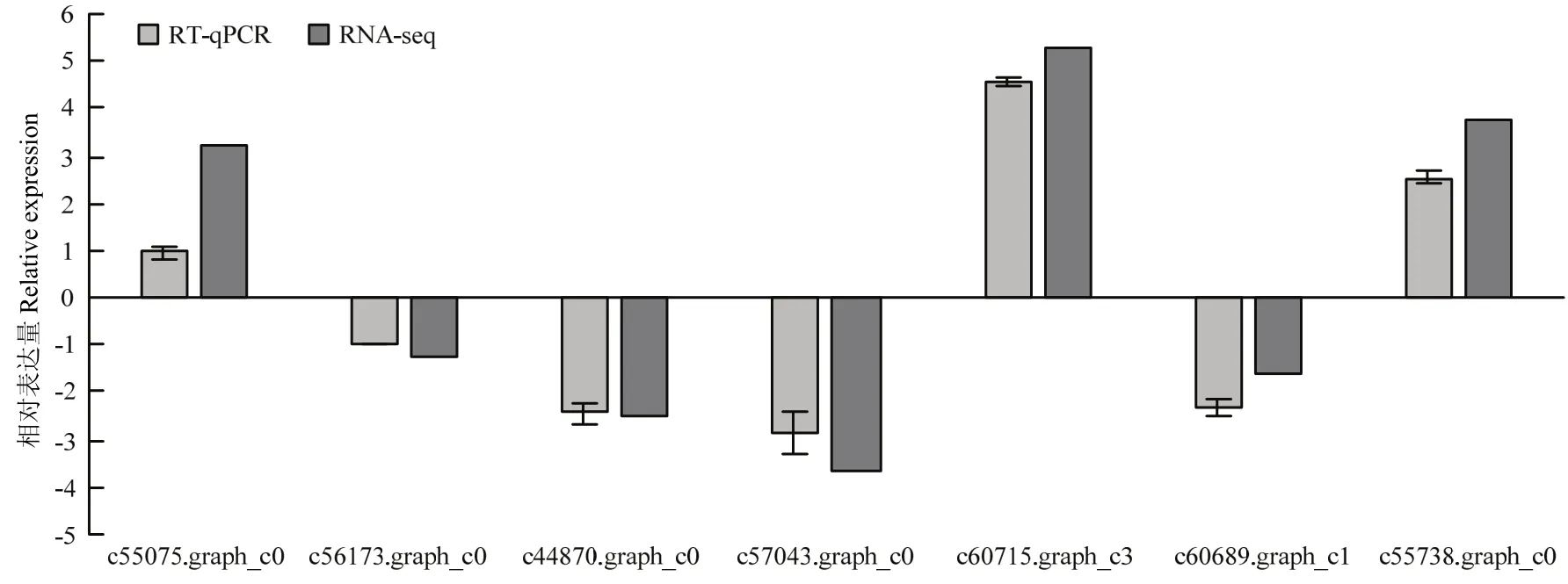
圖7 差異表達基因的RT-qPCR驗證Fig. 7 Quantitative real-time PCR validation of relative expression levels of DEGs
3 討論
多層組學技術的發展使對環境脅迫下的細胞抗逆生理研究從生理代謝和形態結構適應的層面發展到細胞的內在適應機制及大分子物質網絡互作、轉錄水平和翻譯水平調控的分子層面上來[20]。為深入了解鹽脅迫下木霉的分子調控機制,本研究對不同 NaCl脅迫濃度下的耐鹽哈茨木霉 ACCC32527進行轉錄和代謝差異分析,共獲得44 594條unigene和336個代謝產物信息,通過與GO、COG、KEGG、Swiss-Prot、NR等數據庫比對,發現多條與鹽脅迫相關的功能基因及代謝產物。
T2和T3處理下的DEG通過共表達趨勢分析,獲得1 921個DEG的16種基因表達模式,基因的GO功能注釋表明,NaCl對細胞的影響主要集中在催化、結合、細胞器、細胞膜、細胞膜部分、細胞和細胞部分,但上調、下調或不規則變化趨勢基因在主要 GO功能分類上并無明顯差異,表明每部分功能區域所涉及的基因并非全部積極響應 NaCl脅迫,而是其中一部分基因發生上調或下調表達變化。
由于鹽離子的毒性作用,使細胞產生大量ROS,ROS造成的氧化損傷是破壞生物大分子結構和細胞穩態的重要因素[21]。核糖體 RNA(rRNA)是細胞內含量最多的一類RNA,約占RNA總量的80%,更易遭受ROS的攻擊,形成穩定的氧化基損傷[22]。有研究表明,環境脅迫下細胞核糖體的生物合成受到明顯抑制,如LI等[23]研究TDCPP(磷酸三(1,3-二氯-2-丙基)酯)對低等生物毒性試驗時發現,TDCPP能夠抑制四膜蟲(Tetrahymena Thermophila)21個核糖體蛋白編碼基因,并猜測該基因是 TDCPP作用的靶標基因;HASE等[24]發現E. coli核糖體成熟因子(RsgA、RbfA、RimM和RrmJ)或核糖體蛋白S6的缺失可以提高細胞的耐鹽能力,外源添加翻譯抑制劑,如春雷霉素和氯霉素也可以提高細胞的抗逆特性,因此推斷E. coli極可能存在一條由核糖體調節的應急響應通路。本研究中,T2和T3處理下分別有12和18個下調表達基因極顯著富集于核糖體生物合成通路中,表明 NaCl脅迫下 ACCC32527核糖體合成受到明顯抑制,該現象是否與細胞的耐鹽能力相關,目前仍未可知。
根據葡萄糖利用情況和轉錄組數據可知,NaCl脅迫下 ACCC32527對氨基酸、糖類和脂類代謝能力加強。高濃度NaCl(T3)脅迫下,氨基酸代謝相關上調基因比例下降 7%,但對糖類和脂類的代謝分別上調3%和9%。細胞膜的基本結構是磷脂雙分子層,與脂類的代謝密不可分,T3處理下,篩選到一個編碼溶血磷脂酰乙醇胺酰基轉移酶(lysophosphatidylethanolamine acyltransferase,LPEAT)(log2FC=1.01),該蛋白在細胞膜脂肪酸組成的不對稱性和多樣性以及三酰甘油(TAG)中長鏈多不飽和脂肪酸(PUFA)等特殊脂肪酸的引入、改變細胞膜的通透性中起著非常重要的作用[25]。另外,甘油激酶能夠催化甘油生成3-磷酸甘油,與胞內甘油的積累密切相關。T3處理下ACCC32527編碼甘油激酶合成的兩個相關基因均發生下調表達,代謝組數據表明胞內甘油含量上升,可能與細胞累積甘油作為滲透調節物質有關。
微生物的耐鹽機制研究主要集中在:(1)離子轉運系統;(2)活性氧清除系統;(3)細胞滲透調節系統;(4)細胞壁結構。這些機制共同作用使細胞能在高鹽環境下生存[12],如Synechocystissp. PCC 6803編碼K+通道蛋白相關基因(kdpA、ntpJ)的敲除降低細胞對 NaCl的耐受性[26];TEJERA等[27]研究發現,25 mmol·L-1NaCl脅迫下,根瘤菌HB10 SOD活性提高40%—70%,50 mmol·L-1NaCl脅迫下,根瘤菌HB12和HB13 SOD活性提高200%;高鹽環境下,MEENA等[28]發現不動桿菌(Acinetobacter baylyi)能通過提高胞內甘露醇和谷氨酸含量來平衡細胞滲透壓,并發現一個受高鹽誘導,參與甘露醇合成的甘露醇-1-磷酸脫氫酶基因(mltD)。NaCl脅迫下,ACCC32527差異表達基因中篩選到60個氧化還原蛋白基因,其中54個為上調表達基因,包括過氧化物酶、谷胱甘肽過氧化物酶、單加氧酶、過氧化氫酶和硫還原蛋白,參與胞內活性氧的清除。離子轉運調節方面,篩選得到 1個上調表達Na+/K+-ATPase相關基因和8個上調表達的ABC轉運蛋白,維持胞內鹽離子的平衡,調節滲透壓。幾丁質是真菌細胞壁的重要組分之一,在真菌抵御滲透壓、機械力和重金屬等脅迫中起到關鍵作用[29]。KUNCIC等[30]指出細胞壁增厚是真菌Wallemia ichthyophaga在極端高鹽環境下生存的重要機制。木霉幾丁質酶在自身細胞壁的重塑及循環過程中起著極其重要的作用[31]。NaCl脅迫下,ACCC32527共有9個幾丁質酶合成相關基因發生差異表達,且8個基因為上調表達。因此,推測 NaCl脅迫條件下細胞壁幾丁質的降解十分活躍,可能與細胞壁的重塑、再生與加厚密切相關。王麗榮等[19]在0.6 mmol·L-1Cu2+脅迫下哈茨木霉Th-33差異表達基因中篩選發現5個上調表達和3個下調表達的幾丁質酶合成相關基因,并推測是因為Cu2+促進了細胞壁不同幾丁質間的合成與轉化,以維持細胞壁的穩定性。疏水蛋白是細胞壁的延伸結構,是細胞壁外層的一類小分子蛋白,覆蓋在真菌表面,能夠保護真菌抵御惡劣環境[32-33]。本研究中僅檢測到1個上調表達的Ⅱ型疏水蛋白編碼基因,可能一方面形成的疏水表面有利于營養菌絲的生長和孢子擴散,另一方面與細胞壁中的幾丁質β-1,3/1,6-糖苷鍵結合,保護細胞壁結構的完整性。
微生物在外部壓力下會導致基因表達和蛋白質產生的變化,這兩種變化都受到胞內各種穩態控制和反饋機制的影響,這些變化在代謝組的水平上被放大。代謝組學反應的是生物體內小分子物質的含量變化,如氨基酸、有機酸、單糖等,是對轉錄組結果的有效補充[34]。T3處理下,共檢測到 ACCC32527胞內 83種代謝產物含量發生變化,這些物質一部分作為代謝網絡的中間產物,參與細胞生長調節;另一部分為細胞合成的脅迫響應物質,作為滲透調節物質,維持胞內滲透壓穩定,增強菌體耐鹽能力。在差異的代謝產物中,只有34種物質能在LECO-Fiehn Rtx5得到注釋,其中氨基酸及其衍生物變化最為明顯,且含量多為上升,包括谷氨酸、谷氨酸鹽和酪氨酸等。葡萄糖-6-磷酸是各個糖代謝途徑的交叉點,可以進一步在磷酸葡萄糖變位酶作用下產生葡萄糖-1-磷酸。葡萄糖-1-磷酸是細胞內低聚糖和多聚糖通過底物磷酸化降解的產物,在合成代謝中是糖原和核苷酸合成的前體[35]。T3處理下,葡萄糖-6-磷酸含量下降而葡萄糖-1-磷酸含量上升,表明ACCC32527在NaCl脅迫下可能加速糖原的合成,為細胞儲存能源和碳源。6-脫氧-D-葡萄糖作為葡萄糖類似物,通過在細胞內大量積累可以抑制糖酵解作用[36]。NaCl脅迫下,ACCC32527的6-脫氧-D-葡萄糖的含量上升,結合轉錄組數據,可能意味著糖酵解途徑受到抑制而磷酸戊糖途徑等其他途徑得到加強。
此外,本研究仍有近50%差異表達基因被注釋為假定蛋白和約59%未知的差異小分子代謝產物,表明在 NaCl脅迫下,存在許多未知的基因及代謝產物待挖掘。
4 結論
通過轉錄組分析獲得T1 vs T2比較組中637個DEG、T1 vs T3比較組中1 570個DEG和T1 vs T3比較組中的336種差異代謝產物。T2或T3處理下,哈茨木霉 ACCC32527核糖體合成途徑受到明顯抑制,而抗氧化酶、轉運蛋白及胞外結構相關基因多數發生上調表達。結合代謝轉錄組及代謝組數據,發現甘油在胞內累積量上升,推測可能參與胞內的滲透調節。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
