野生和栽培大豆種質油脂組成特點及其與演化的關系
2019-06-25 02:15:20張晟瑞王連錚孫君明
作物學報 2019年7期
陳 影 張晟瑞 王 嵐 王連錚 李 斌 孫君明
?
野生和栽培大豆種質油脂組成特點及其與演化的關系
陳 影**張晟瑞**王 嵐 王連錚 李 斌*孫君明*
中國農業科學院作物科學研究所/ 作物分子育種國家工程實驗室/ 農業部農村大豆生物學重點實驗室, 北京 100081
以58份不同類型(野生、半野生和栽培)大豆種質為材料, 利用32對SSR標記分析大豆種質間的遺傳多樣性和進化關系, 采用NIRS和GC方法分別分析大豆脂肪含量和脂肪酸組分含量, 研究不同類型大豆種質油脂組成特點及其與演化的關系。結果顯示, 野生大豆和栽培大豆的油脂組成存在顯著差異, 栽培大豆脂肪含量(平均20.8%)顯著高于野生大豆(平均10.49%), 油酸含量(平均28.5%)顯著高于野生大豆(平均14.37%), 而亞麻酸含量卻顯著低于野生大豆; 由相關性分析可知, 大豆種子中的脂肪與油酸含量顯著正相關(= 0.85**), 而與其他脂肪酸組分極顯著負相關; 油酸與所有其他脂肪酸組分均負相關, 特別是與亞麻酸和亞油酸呈極顯著負相關(=-0.90**和-0.89**); 油脂組成和SSR標記對不同類型大豆種質的聚類和主成分分析表明, 2種分類結果基本一致, 可分為栽培和野生2個亞群, 半野生大豆則分布于2個亞群中。由此可見, 大豆油脂組成與大豆種質的馴化程度有關, 脂肪含量和亞麻酸含量可以作為大豆演化分類的參考指標。
野生大豆; 半野生大豆; 栽培大豆; 脂肪含量; 脂肪酸組分
大豆(L. Merr.)是世界上四大油料作物之一, 為食用植物油的主要來源。大豆油中含有豐富的脂肪酸, 主要包括棕櫚酸、硬脂酸、油酸、亞油酸和亞麻酸, 其中棕櫚酸和硬脂酸為飽和脂肪酸, 油酸、亞油酸和亞麻酸為不飽和脂肪酸[1-2]。不同脂肪酸組成決定大豆油的品質和最終用途[3]。多不飽和脂肪酸可以促進體內膽固醇的代謝, 對預防心血管疾病有著顯著的作用[4]。另一方面, 多不飽和脂肪酸亞油酸和亞麻酸中含有較多的不飽和鍵, 易被脂肪氧化酶氧化, 從而降低大豆油的保存期限和品質[5-6]。飽和脂肪酸含量升高可以提高豆油的穩定性, 但因其很難被人體所吸收, 攝入量過高易引發高血脂、冠心病、肥胖癥等疾病, 不利于人體的健康[7-8], 因此, 依據不同的需求選育不同脂肪酸組成的大豆品種是十分必要的。根據系統進化關系, 大豆種質可分為野生()、半野生()、栽培大豆() 3種類型。前人利用大豆形態學分類方法研究種質間馴化關系, 并對不同類型大豆種質的脂肪含量和脂肪酸組分分析發現, 栽培豆中的脂肪含量顯著高于野生豆[9]; 且不同類型大豆間的油酸、亞油酸和亞麻酸組成存在顯著差異[1]。隨著分子生物學技術的迅速發展, AFLP、SSR和SNP等分子標記被廣泛應用于大豆種間親緣關系和演化關系的研究[10-12]。本文采用近紅外光譜和氣相色譜分析方法分別分析野生、半野生和栽培大豆籽粒中的脂肪含量和脂肪酸組成, 結合大豆形態學和分子標記分類方法, 研究其油脂組成特點, 探討在大豆馴化過程中油脂組成的演化規律, 為大豆品質改良育種提供理論參考。
1 材料與方法
1.1 大豆試驗材料
選用不同類型的大豆種質58份, 根據形態學特征將其分成3種類型, 即27份野生大豆、7份半野生大豆和24份栽培大豆(表1)均由本單位大豆基因資源創新研究組邱麗娟研究員提供。2017年6月將材料種植于中國農業科學院作物科學研究所北圃場基地, 采用完全隨機區組設計, 單行區, 行長2.0 m, 行距0.5 m, 株距0.1 m, 按株行收獲脫粒, 4℃保存待用。

表1 58份不同類型的大豆種質
1.2 大豆葉片DNA的提取
于苗期取每份大豆種質0.1~0.2 g幼葉放入研缽, 加液氮充分研磨, 取100 mg樣品放入1.5 mL離心管。使用TIANGEN新型植物基因組DNA提取試劑盒(離心柱型)提取大豆葉片基因組DNA。利用1%瓊脂糖凝膠電泳檢測DNA質量, 使用NanoDrop 2000紫外-可見分光光度計檢測DNA的濃度和純度。
1.3 大豆種質基因型分型
參照王彪等[13]的研究方法選用32對均勻分布在大豆20個遺傳連鎖群上的SSR引物, 根據SoyBase網站(https://soybase.org/)提供的SSR引物序列, 由北京梓熙生物科技有限公司合成, 并使用FAM (藍光)和HEX (紅光) 2種熒光基團修飾上游引物。PCR主要試劑來自全式金生物技術有限公司。PCR反應體系總體積15 μL, 其中1.5 μL 10×PCR buffer、1.5 μL 2.5 mmol L-1dNTP、正反向引物各0.5 μL、0.2 μLpolymerase (5 U μL-1)、大豆基因組DNA 50 ng、9.8 μL ddH2O。PCR擴增條件為95℃預變性3 min; 95℃變性30 s, 50℃退火30 s, 72℃延伸30 s, 擴增35個循環; 72℃延伸10 min。
利用毛細管電泳法對58份大豆種質材料的PCR擴增產物進行基因型檢測。取100~200 ng的PCR產物放入96孔板中, 加入50 μL無水乙醇, 震蕩混勻, 4℃, 3000 ×離心30 min, 棄上清液; 加入70 μL預冷的(-20℃) 70%乙醇, 震蕩混勻, 4℃, 2000 ×離心15 min, 棄上清液; 室溫避光放置10~20 min; 加入10 μL Hi-Pi甲酰胺(混有0.3% ROX內標), 震蕩混勻, 95℃, 變性5 min; 冰浴5 min, 1000 ×瞬時離心后利用ABI 3730XL測序儀進行毛細管電泳。
1.4 大豆脂肪和脂肪酸組分含量測定
1.4.1 大豆脂肪含量檢測 采用傅里葉近紅外光譜分析儀(MATRIX-1, Bruker, 德國)掃描樣品, 每個樣品掃描3次, 利用OPUS 5.0軟件對大豆種質的脂肪含量進行預測, 取3次檢測結果的平均值為脂肪含量。
1.4.2 大豆脂肪酸提取 采用加熱甲酯化法提取脂肪酸, 每份大豆種質取20粒種子, 用磨樣機(Retsch ZM100,= 1.0 mm, Rheinische, 德國)磨成細粉, 準確稱取150.0 mg豆粉于10.0 mL試管中, 加入5.0 mL正己烷, 60℃浸提20 min; 然后加入5.0 mL 0.5 mol L-1甲醇鈉溶液, 充分搖勻10 min, 使其充分甲酯化, 靜置, 取上清液置色譜專用樣品瓶中檢測[14]。
1.4.3 大豆脂肪酸氣相色譜檢測條件 采用島津GC-2010氣相色譜儀對大豆脂肪酸含量進行定性定量檢測。用色譜柱RTX-Wax (30 m × 0.25 m × 0.25 m); 自動注射l μL; 分流比40∶1; 注入口溫度250℃, 載氣氮氣54 mL min-1, 氫氣40 mL min-1, 空氣400 mL min-1, 采用程序升溫模式(180℃保持1.5 min, 以10℃ min-1升到210℃, 保持2 min, 然后以5℃ min-1升到220℃, 保持5 min)。檢測器FID溫度為300℃; 采用面積歸一化法計算5種主要脂肪酸(棕櫚酸、硬脂酸、油酸、亞油酸和亞麻酸)的百分比作為大豆脂肪酸組分含量[15]。
1.5 數據分析
利用Microsoft Excel 2007和SAS 9.2軟件進行數據統計與方差分析。利用PowerMarker V3.25軟件進行標記的遺傳多樣性和系統進化樹分析。使用Mcquitty方法[16]對脂肪酸含量進行聚類和主成分分析等。
2 結果與分析
2.1 58份不同類型大豆種質脂肪和脂肪酸組成特點
58份不同類型大豆種質的脂肪和脂肪酸組分含量存在顯著差異。由表2可知, 大豆種質脂肪含量的變異系數高達34.64%, 其中, 栽培大豆品種合豐50的脂肪含量最高(24.71%), 野生大豆ZYD01694的脂肪含量最低(6.97%); 5種脂肪酸組分含量的變異系數也各不相同(8.59%~42.82%), 其中亞麻酸的變異系數最大(42.82%), 野生大豆BY019的亞麻酸含量最高(15.97%), 栽培品種合豐50含量最低(3.45%); 油酸的變異系數次之(38.16%), 其中野生大豆ZYD3263油酸含量最低(11.00%), 半野生大豆ZYD0001油酸含量最高(39.61%)。總體上看, 飽和脂肪酸含量的變異系數為10.23%, 不飽和脂肪酸含量的變異系數為2%, 由此可見, 大豆種子中飽和、不飽和脂肪酸比例是相對比較穩定。對58份大豆資源材料的油脂特征頻率分布分析發現, 脂肪含量和脂肪酸組分均呈現連續分布。盡管5種脂肪酸含量在58份材料中不是典型的正態分布, 但飽和脂肪酸和不飽和脂肪酸含量卻呈現典型的正態分布(圖1)。
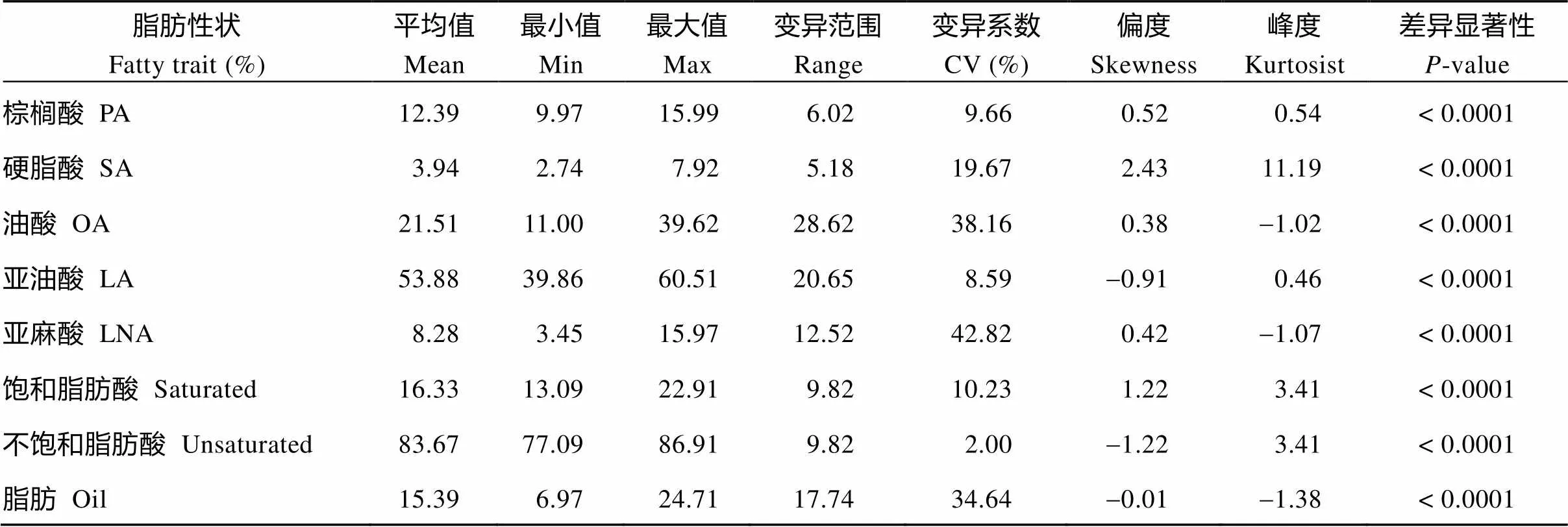
表2 58份大豆種質的脂肪和脂肪酸組成含量變異分析
PA: palmitic acid; SA: stearic acid; OA: oleic acid; LA: linoleic acid; LNA: linolenic acid.

圖1 58份材料油脂性狀的頻率分布
縮寫同表2。Abbreviations are the same as those given in Table 2.
2.2 不同類型大豆種質脂肪和脂肪酸組分的差異比較分析
按照形態學分類方法將58份不同類型大豆種質分為野生、半野生和栽培大豆3種類型(表1)。通過對3種類型大豆種質油脂組成的方差分析發現, 野生大豆的脂肪含量和脂肪酸組分與栽培大豆均存在顯著差異, 與半野生大豆除硬脂酸外也均存在顯著差異。野生大豆脂肪含量(平均10.49%)顯著低于栽培大豆(平均20.8%), 半野生大豆位于二者之間(平均15.71%); 野生大豆不飽和脂肪酸含量(平均82.71%)顯著低于栽培大豆(平均84.7%), 而飽和脂肪酸含量(平均17.29%)顯著高于栽培大豆(平均15.3%)。半野生大豆飽和脂肪酸和不飽和脂肪酸含量均與野生大豆差異不顯著。野生大豆的油酸含量(平均14.37%)顯著低于栽培大豆(平均28.5%), 其他脂肪酸組分含量高于栽培大豆。野生大豆亞麻酸含量(平均11.38%)顯著高于栽培大豆(平均4.97%), 而半野生大豆同樣位于二者之間(平均7.64%)。因此, 脂肪和亞麻酸含量在不同類型大豆中差異顯著(圖2)。
2.3 大豆種質的脂肪含量和脂肪酸組分相關性分析
由圖3可知, 大豆種子中的脂肪含量與油酸含量存在極顯著的正相關(= 0.85**), 而與其他脂肪酸組分存在極顯著負相關, 特別是與亞麻酸含量呈極顯著負相關(=-0.92**)。油酸與所有其他脂肪酸組分均存在負相關, 特別是與亞麻酸、亞油酸呈極顯著負相關(=-0.90**和-0.89**); 而亞麻酸除了與油酸含量呈極顯著負相關(=-0.90**)外, 與其他組分均存在顯著正相關。另外, 2種飽和脂肪酸棕櫚酸和硬脂酸呈極顯著正相關(= 0.41**)。
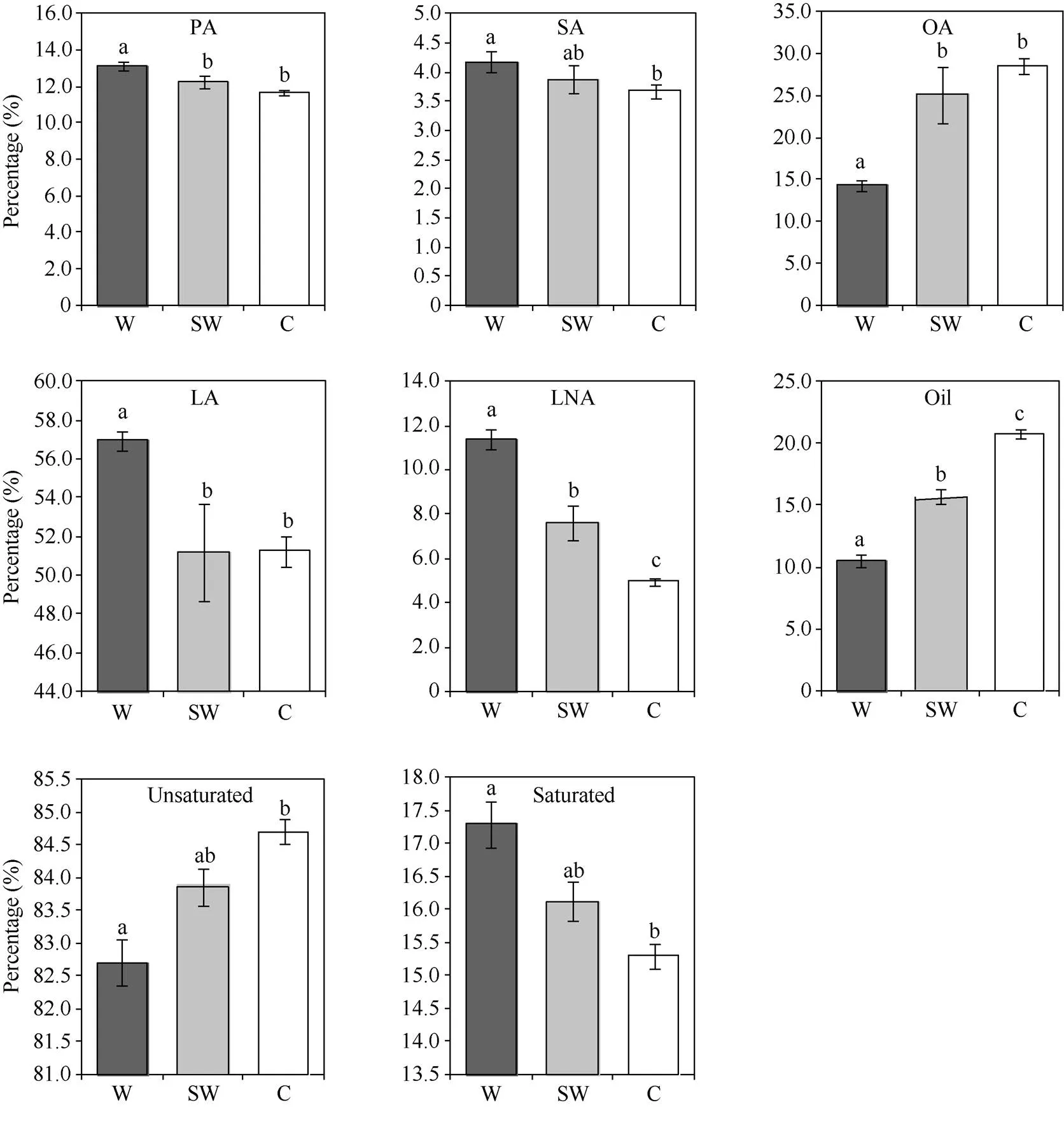
圖2 基于形態學分類的野生、半野生和栽培大豆種質的油脂組成的方差分析
圖中符號W、SW、C分別代表野生大豆()、半野生大豆()和栽培大豆()。標以不同字母的柱值在0.05水平上差異顯著。縮寫同表2。
W, SW, and C represent wild soybean (), semi-wild soybean (), and cultivated soybean (), respectively. Bars superscripted by different letters are significantly different at the 0.05 probability level. Abbreviations are the same as those given in Table 2.

圖3 58份大豆種質的脂肪含量和脂肪酸組分間的Pearson相關系數
*和**分別表示0.05和0.01水平差異顯著; 圖中圓圈越大, 表示相關性越強, 圓圈越小, 表示相關性越弱。縮寫同表2。
*and**represent the significant level at the 0.05 and 0.01 probabi-lity levels, respectively. The larger the circle, the stronger the correlation, the smaller the circle, the weaker. Abbreviations are the same as those given in Table 2.
2.4 基于油脂組成參數的不同類型大豆種質的聚類分析
以大豆種質的脂肪含量和5種脂肪酸組分含量為參數, 聚類分析(圖4)表明, 58份大豆種質可分為3個亞群, 第1亞群僅由半野生大豆種質SW5組成; 第2亞群主要由野生大豆種質組成, 包含25個野生和2個半野生大豆種質, 第3亞群主要由栽培大豆種質組成, 包含24個栽培、4個半野生和2個野生大豆種質。野生大豆與栽培大豆明顯分成2個亞群, 說明2種類型的大豆種質的油脂組成明顯不同, 同一類型大豆種質的油脂組成分布規律較為統一, 不同類型大豆種質間存在較大差異; 而半野生大豆無明顯的聚集群, 分布在野生和栽培大豆亞群中。
2.5 不同類型大豆種質基于油脂組成的主成分分析
以大豆種質的脂肪含量和5種脂肪酸組分含量為參數, 對58份大豆種質進行主成分分析(圖5), 共提取解釋變異最高的前3個主成分因子, 可累計反映總變異的93%, 第一主成分(PC1)主要由油酸、亞麻酸和脂肪含量組成, 第二主成分(PC2)主要由硬脂酸和亞油酸組成, 第三主成分(PC3)主要由棕櫚酸組成, 3個主成分因子分別揭示總變異的65%、19.7%和8.5%。野生大豆和栽培大豆被分別聚集在2個亞群中, 半野生大豆則在2個亞群中均有分布。主成分分析與聚類分析結果基本一致, 58份大豆種質中, 野生大豆與栽培大豆脂肪含量、脂肪酸含量差異較明顯, 半野生大豆則處在野生和栽培大豆之間, 無明顯油脂組成性狀的差異。
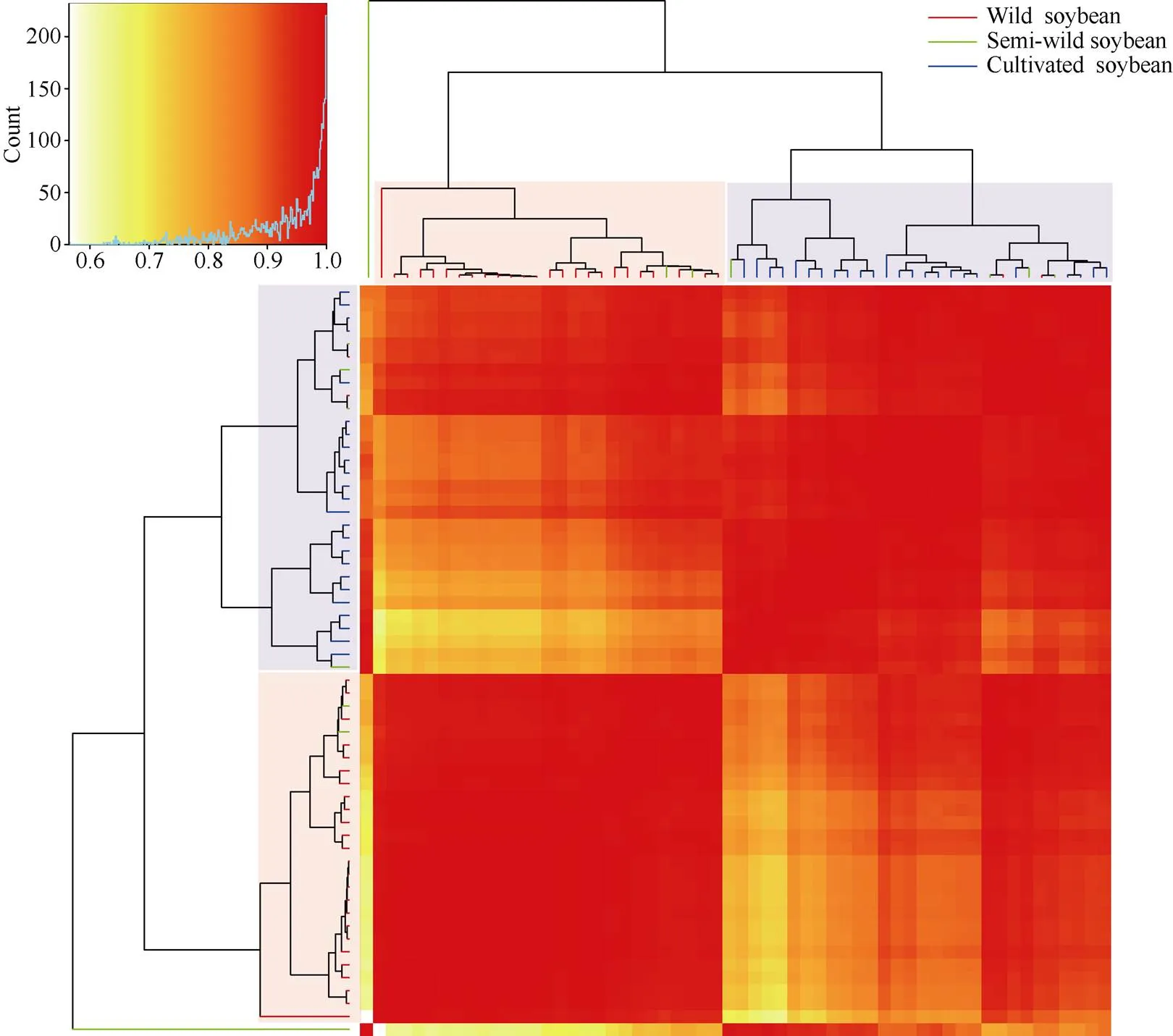
圖4 58份大豆種質基于油脂組成的聚類分析
聚類結果中紅色分支代表野生大豆, 綠色分支代表半野生大豆, 藍色分支代表栽培大豆。
The red branches represent wild soybean, the green branches represent semi-wild soybean, and the blue branches represent cultivated soybean in the clustering results.

圖5 58份大豆種質基于油脂組成的主成分分析
PC1~PC3代表基于脂肪和5種脂肪酸組成含量的前3個主成分。紅色圓點代表野生大豆, 綠色三角形代表半野生大豆, 藍色正方形代表栽培大豆。
PC1-PC3 represent the first three principal components based on the oil and five fatty acid contents. The red circles, green triangles, and blue squares represent wild, semi-wild, and cultivated soybean accessions, respectively.
2.6 基于SSR標記的不同類型大豆種質的遺傳多樣性分析
利用32對SSR引物對大豆種質進行遺傳多樣性分析。由表3可知, 32對SSR引物在58份大豆種質中共擴增出等位基因368個, 平均每對引物擴增出11.5個等位基因, 等位基因數目變異系數為5~18個, 引物Satt 390僅擴增出5個等位基因, 引物Satt 281和Satt 590分別擴增出多達18個等位基因。
遺傳多樣性代表生物種群之內和種群之間遺傳結構變異程度。基因多樣度和多態性信息含量(PIC)是常用來表示種群的遺傳多樣性的指標, 本研究結果表明, 基因多樣性平均值為0.80, 各引物基因多樣性值變異系數為0.61~0.91, 其中引物Satt 590基因多樣性最高, 引物Satt 281次之, 引物Satt 442最低。PIC值與基因多樣性呈現的結果較為一致, 其平均值為0.78, 變異系數為0.53~0.90, 引物Satt 590與Satt 281的PIC值最大, 均為0.90, 引物Satt 442的PIC值最低(0.53)。
(續表3)

引物Primer連鎖群Linkage group染色體Chr.等位基因數目Allele number基因多樣性Gene diversity多態性信息含量PIC Satt 586F13110.830.81 Satt 309G1860.730.69 Satt 442H1280.610.53 Satt 239I20120.870.85 Satt 571I2080.790.76 Sct 189I20120.830.81 Satt 431J16150.890.89 Satt 596J16100.860.84 Satt 242K9110.870.86 Satt 588K9110.800.78 Satt 373L19120.800.79 Satt 308M7130.850.84 Satt 346M790.730.69 Satt 590M7180.910.90 Satt 530N3120.820.80 平均Mean——11.50.800.78
2.7 基于SSR標記的不同類型大豆種質的聚類分析
基于Nei (1983)氏距離, 采用鄰位相連算法(Neighbor-Joining tree, NJ)[17]對58份大豆種質進行聚類分析。由圖6可以看出, 58份大豆種質可明顯分為兩大亞群, 第一亞群(CI)主要由野生大豆組成, 包含23份野生和5份半野生大豆; 第二亞群(CII)主要由栽培大豆組成, 包含24份栽培、2份半野生和4份野生大豆; 半野生大豆分布于兩大亞群之中, 表明利用32對SSR標記, 可以大致區分栽培大豆和野生大豆類群, 但不能準確區分半野生大豆類群。第一亞群中的G1類群(全部為野生大豆類型)與第二亞群中的G2類群(全部為栽培大豆類型)遺傳距離最遠, 表明這2個類群的大豆親緣關系較遠, 可作為典型的野生和栽培大豆類型。本研究從這2個典型類群中發現了14個特異性標記, 分別為Satt 236、Satt 346、Satt 586、Satt 409、Satt 300、Satt 596、Satt 308、Satt 590、Satt 286、Sct_189、Satt 556、Satt 530、Satt 242和Satt 431。
2.8 基于SSR標記的不同類型大豆種質的主成分分析
基于32對SSR標記對58份大豆種質進行主成分分析發現, 前3個解釋變異最高的主成分因子PC1a、PC2a、PC3a分別揭示總變異的17.83%、9.42%和7.76% (此處PC1a~PC3a代表不同SSR標記組合所構成的主成分因子)。由圖7可以看出, 58份不同類型的大豆種質可明顯分為2個亞群, 野生大豆和半野生大豆聚集在一個亞群中, 栽培大豆聚集在另外一個亞群, 其中有少量野生大豆。主成分與聚類分析結果均表明, 58份大豆種質中, 野生大豆與栽培大豆親緣關系較遠, 而半野生大豆與兩者均存在一定的親緣關系。
2.9 基于SSR標記分類的典型野生和栽培大豆種質的油脂組成特點
SSR標記遺傳分類結果(圖6)及典型野生大豆(9份)和栽培大豆(12份)脂肪含量和脂肪酸組成含量的方差分析(圖8)顯示, 野生大豆脂肪含量(平均9.02%)極顯著低于栽培大豆(平均21.6%); 野生大豆油酸含量(平均12.1%)顯著低于栽培大豆(平均29.07%), 其他脂肪酸組分野生大豆均高于栽培大豆, 特別是野生大豆的亞麻酸含量(平均12.26%)顯著高于栽培大豆(平均4.71%); 野生大豆飽和脂肪酸含量高于栽培大豆, 不飽和脂肪酸正相反。這與基于形態學區分的野生和栽培大豆種質的油脂組成規律基本一致。

圖6 基于SSR標記的大豆種質的NJ聚類圖
圖中紅色、綠色和藍色分支分別代表野生、半野生和栽培大豆; CI和CII表示2個主要亞群, G1和G2分別表示聚類結果中的親緣關系較遠且形態差異明顯的典型野生大豆()和栽培大豆()。
The red, green, and blue branches represent the wild, semi-wild, and cultivated soybean accessions, respectively; CI and CII indicate two major subgroups, G1 and G2 represent typical wild soybean () and cultivated soybean () with distant genetic relationships and obvious morphological differences in clustering results.
3 討論
3.1 野生大豆和栽培大豆的油脂組成存在顯著差異
大豆起源于中國, 早在5000年前野生大豆()就被馴化為栽培大豆()[12], 二者存在緊密的親緣關系, 但油脂組成特點不同。徐豹等[18]分析了中國1595份野生大豆的脂肪含量, 平均為9.94%, 低于栽培大豆(平均19.05%)。王連錚等[19]針對東北地區收集的大量野生大豆資源分析了野生大豆和栽培大豆的脂肪酸組成差異, 發現野生大豆的油酸含量(平均15.58%)低于栽培大豆(平均28.86%), 亞麻酸含量(平均18.69%)高于栽培大豆(平均7.35%)。本研究針對分子數據分類的典型野生大豆的平均脂肪含量為9.02%, 典型的栽培大豆的平均脂肪含量為21.61%; 野生大豆油酸含量(12.10%)同樣低于栽培大豆(29.08%), 亞麻酸含量(12.26%)同樣高于栽培大豆(4.71%), 表明野生大豆和栽培大豆的油脂組成存在顯著差異, 這與前人的大樣本研究結果吻合。
3.2 通過分子標記方法可以對野生和栽培大豆種質進行準確分類
大豆屬種間遺傳關系的研究有多種方法, 利用形態學特征區分大豆不同類型最為常用。然而, 大豆形態學特征易受環境條件影響, 不能準確用于判定種質材料的種屬關系[20]。本研究利用大豆形態學特征、品質特性及種間分子親緣關系, 對58份大豆種質進行了種屬分類, 并鑒定種質間的進化關系, 獲得了9份典型野生大豆和12份典型栽培大豆類型, 針對2種類型的大豆種質篩選出了特異性標記和特征條帶。趙洪餛等[21]研究表明BARC-sat39標記在野生大豆和栽培大豆之間有特異譜帶, 推斷該SSR標記是栽培大豆和野生大豆有關的一個等位基因點。本研究篩選出的14個野生大豆和栽培大豆的特異性SSR標記, 未見相關報道, 可能是新的特異標記, 下一步將在包含栽培和野生大豆群體的更大范圍內進一步驗證其有效性。
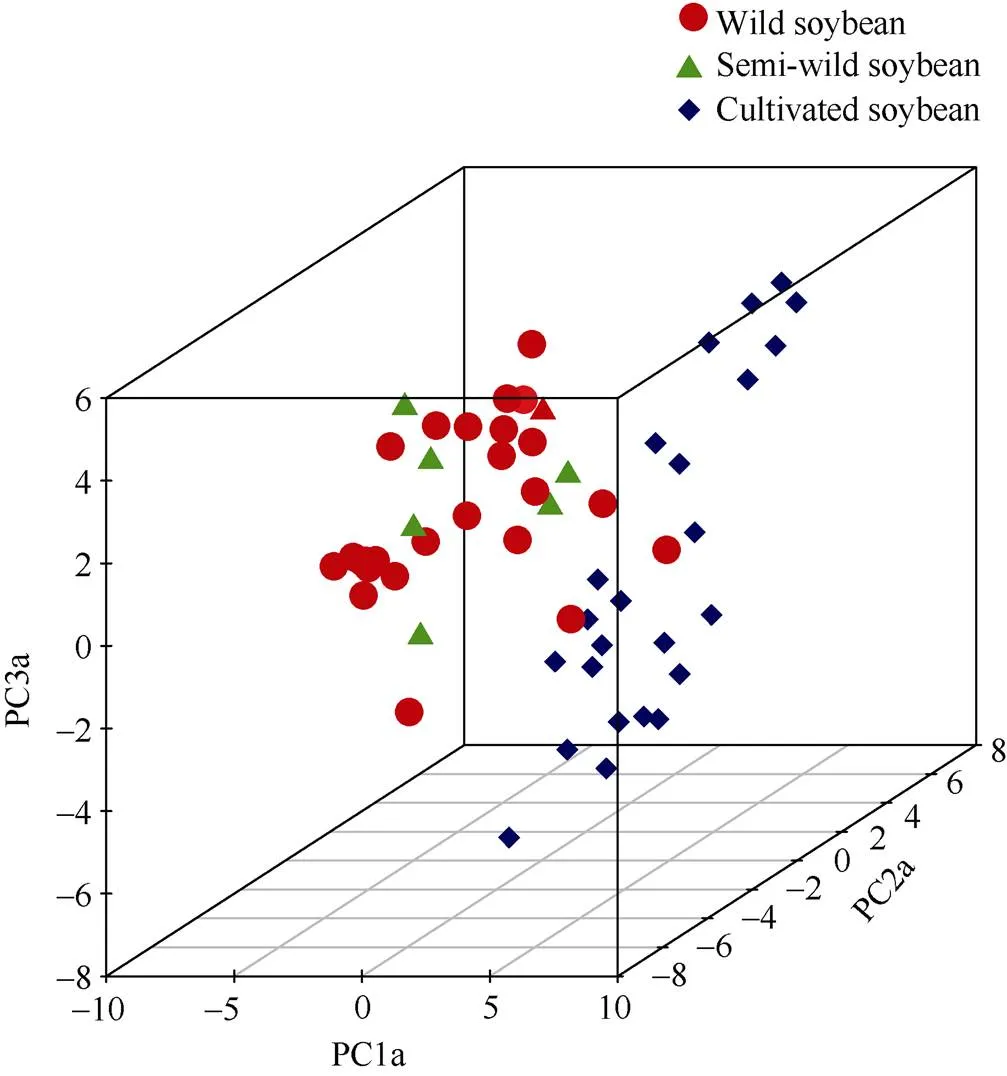
圖7 58份大豆材料基于32對SSR遺傳標記的主成分分析
PC1a~PC3a代表基于32對SSR引物的前3個主成分因子。紅色圓點代表野生大豆, 綠色三角形代表半野生大豆, 藍色正方形代表栽培大豆。
PC1a-PC3a represent the first three principal components based on the 32 pairs of SSR markers. The red circle, green triangle, and blue square represent wild, semi-wild and cultivated soybean accessions, respectively.
3.3 大豆油脂組成特點與大豆的馴化與分類有關
栽培大豆是野生大豆在栽培條件下通過人工定向選擇而來[22], 半野生豆是野生大豆和栽培大豆中間的過渡類型[23]。本文利用分子標記方法結合大豆形態學特征對58份大豆種質進行聚類分析和主成分分析, 發現野生大豆與栽培大豆可以明顯分為2個亞群, 而半野生大豆則同時分布于2個亞群, 這可能是本研究選取的分子標記數目較少, 不能完全區分野生和半野生大豆, 也可能是半野生大豆無論親緣關系還是油脂組成特征均居于栽培大豆和野生大豆之間。
方差分析表明, 大豆從野生大豆到栽培大豆的演化過程中, 脂肪含量顯著增加; 在脂肪酸組成上, 主要趨勢是油酸含量大幅增加, 亞油酸和亞麻酸含量顯著降低。食用過多的飽和脂肪酸不利于人體健康, 而不飽和脂肪酸有益于心血管疾病的預防[24], 同時由于多不飽和脂肪酸亞麻酸容易被氧化造成豆油的保存期縮短, 因此提高油酸含量可以保證大豆油健康品質, 是大豆品質育種的主要研究方向。由此推測, 栽培大豆從野生的演化過程中, 脂肪含量和脂肪酸組成可能受到了人工選擇。近年來對302份野生和栽培大豆的重測序分析也表明, 控制油脂合成的多個重要位點和基因(等)均受到了顯著的選擇[12], 這些位點和基因在馴化過程中的定向選擇可能是大豆油脂組成演化的重要遺傳基礎。同時, 發掘這些重要基因的優異等位變異, 并結合常規育種技術、分子標記輔助選擇技術、以及轉基因技術將這些基因的優異等位變異聚合是進一步優化大豆油脂組成的有效途徑。
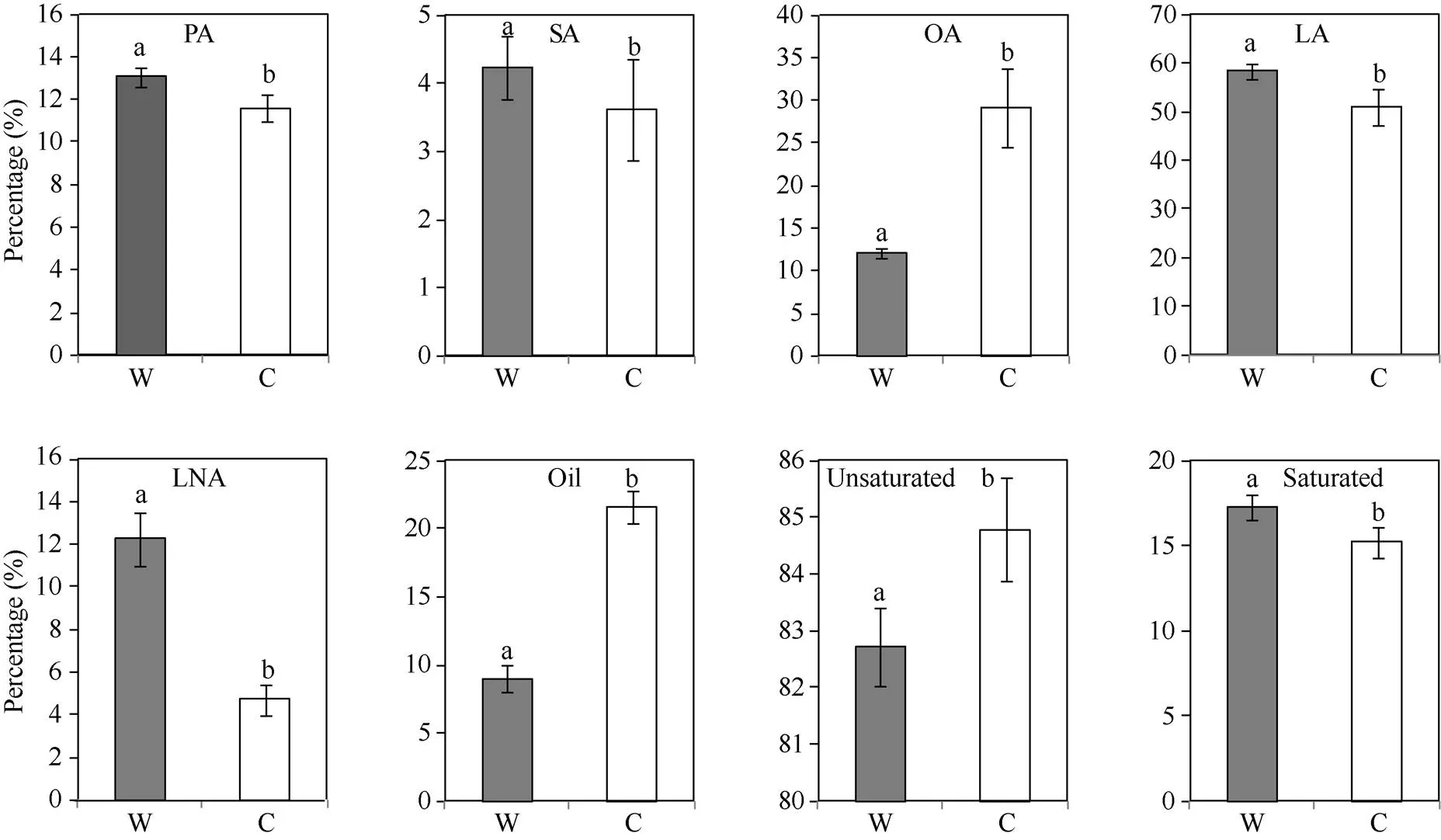
圖8 基于遺傳距離分析結果對野生和栽培大豆種質油脂組成的方差分析
W、C分別代表野生大豆()和栽培大豆(); 標以不同字母的柱值在0.05水平上差異顯著。縮寫同表2。
W and C represent wild soybean () and cultivated soybean (), respectively. Bars superscripted by different letters are significantly different at the 0.05 probability level. Abbreviations are the same as those given in Table 2.
α-亞麻酸是2種重要n-3長鏈脂肪酸EPA和DHA的合成前體, 這2種脂肪酸在大腦發育、心血管健康以及炎癥反應中具有重要的作用[25], 而野生大豆中的亞麻酸含量高達15%以上, 因此, 在大豆品質育種中可以利用野生大豆資源, 通過雜交組配, 提高栽培大豆中的亞麻酸含量, 用于專用營養大豆品種的選育。
3.4 脂肪含量和亞麻酸含量可以作為野生和栽培大豆進化分類的參考指標
莊無忌等[1]、呂景良等[2]和徐豹等[18]研究發現亞麻酸含量在栽培和野生大豆間存在顯著差異, 并提出了將亞麻酸含量作為大豆種或類型鑒定的一個參考指標。本文從油脂特征和親緣關系2個方面對58份大豆種質聚類分析和主成分分析顯示, 野生大豆與栽培大豆被分別聚集在2個亞群內, 半野生大豆則在2個聚集群內均有分布; 其中, 51份(88%)大豆種質基于油脂性狀聚類結果與基于分子親緣關系的聚類結果一致, 說明從野生大豆到栽培大豆的演化過程中油脂組成發生了顯著的改變, 特別是, 脂肪含量和亞麻酸含量在不同類型大豆種質中差異十分顯著, 而且隨著演化程度的提高而呈規律性的變化, 因此可將其作為大豆演化與分類研究的參考指標。
4 結論
大豆油脂組成與大豆種質的馴化程度有關, 栽培大豆脂肪含量及油酸含量均顯著高于野生大豆, 而亞麻酸含量顯著低于野生大豆。依據油脂組成和分子標記作為分類參數的聚類結果基本一致, 野生和栽培大豆明顯分為2個亞群, 而半野生大豆分布在2個亞群中。栽培和野生大豆的脂肪含量和亞麻酸含量存在明顯分布規律, 可以作為大豆演化分類的參考指標。
[1] 莊無忌, 韓華瓊, 謝發明, 張喬, 李福山, 舒世珍, 常汝鎮. 栽培、野生、半野生大豆脂肪酸組成的初步分析研究. 大豆科學, 1984, 3: 223–230. Zhuang W J, Han H W, Xie F M, Zhang Q, Li F S, Shu S Z, Chang R Z. Composition of fatty acid in cultivated, semi-cultivated and wild soybean.1984, 3: 223–230 (in Chinese with English abstract).
[2] 呂景良, 邵榮春, 吳百靈,梁岐, 吳桂榮. 東北地區大豆品種資源脂肪酸組成的分析研究. 作物學報, 1990, 16: 349–356. Lyu J L, Shao R C, Wu B L, Liang Q, Wu G R. Studies on the fatty acid composition of soybean germplasm resources in northeast China.,1990, 16: 349–356 (in Chinese with English abstract).
[3] Thelen J J, Ohlrogge J B. Metabolic engineering of fatty acid biosynthesis in plants., 2002, 4: 12–21.
[4] 李志香, 沈翠平. 多不飽和脂肪酸對人體的作用. 生物學通報, 1998, 33(1): 9–10. Li Z X, Shen C P. The effect of polyunsaturated fatty acids on the human health., 1998, 33(1): 9–10 (in Chinese).
[5] Willett W C, Stampfer M J, Manson J E, Hu F B, Oh K. Dietary fat intake and the risk of coronary heart disease in women., 1997, 337: 1491–1499.
[6] Mounts T L, Warner K, List G R,Kleiman R,Fehr E G,Hammond E G, WilcoxJ R. Effect of altered fatty acid composition on soybean oil stability., 1988, 65: 624–628.
[7] Judd J T, Clevidence B A, Muesing R A, Wittes J, Sunkin M E, Podczasy J J. Dietary trans fatty acids: effects on plasma lipids and lipoproteins of healthy men and women., 1994, 59: 861–868.
[8] Krisetherton P M. Trans-fats and coronary heart disease., 2010, 50: 29–30.
[9] 李福山. 中國野生大豆資源的地理分布及生態分化研究. 中國農業科學, 1993, 26(2): 47–55. Li F S. Studies on the ecological and geographical distribution of the Chinese resources of wild soybean()., 1993, 26(2): 47–55 (in Chinese with English abstract).
[10] Zhao H K, Zhuang B C, Wang Y M, Li Q J. AFLP analysis of wild () and cultivated soybean () in China., 2000, 10: 32–35.
[11] 吳曉雷, 賀超英, 陳受宜, 莊炳昌, 王克晶, 王學臣. 用SSR分子標記研究大豆屬種間親緣進化關系. 遺傳學報, 2001, 28: 359–366. Wu X L, He C Y, Chen S Y, Zhuang B C, Wang K J, Wang X C. Phylogenetic analysis of interspecies in genusthrough SSR markers., 2001, 28: 359–366 (in Chinese with English abstract).
[12] Zhou Z K, Jiang Y, Wang Z, Gou Z H, Lyu J, Li W Y, Yu Y J, Shu L P, Zhao Y J, Ma Y M, Fang C, Shen Y T, Liu T F, Li C C, Li Q, Wu M, Wang M, Wu Y S, Dong Y, Wan W T, Wang X, Ding Z L, Gao Y D, Xiang H, Zhu B G, Lee S H, Wang W, Tian Z X. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean., 2015, 4: 408–414.
[13] 王彪, 常汝鎮, 陶莉, 關榮霞, 閆麗, 張明恢, 馮忠孚, 邱麗娟. 分析中國栽培大豆遺傳多樣性所需SSR引物的數目. 分子植物育種, 2003, 1: 82–88. Wang B, Chang R Z, Tao L, Guan R X, Yan L, Zhang M H, Feng Z F, Qiu L J. Identification of SSR primer numbers for analyzing genetic diversity of Chinese soybean cultivated soybean., 2003, 1: 82–88 (in Chinese with English abstract).
[14] 范勝栩, 李斌, 孫君明, 韓粉霞, 閆淑榮, 王嵐, 王連錚. 氣相色譜方法定量檢測大豆5種脂肪酸. 中國油料作物學報, 2015, 37: 548–553. Fan S X, Li B, Sun J M, Han F X, Yan S R, Wang L, Wang L Z. A quantitative gas chromatographic method for determination of soybean seed fatty acid components., 2015, 37: 548–553 (in Chinese with English abstract).
[15] Ma L, Li B, Han F X, Yan S R, Wang L Z, Sun J M. Evaluation of the chemical quality traits of soybean seeds, as related to sensory attributes of soymilk., 2015, 173: 694–701.
[16] McQuitty L L. Similarity analysis by reciprocal pairs for discrete and continuous data., 1996, 26: 825– 831.
[17] Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees., 1987, 4: 406–425.
[18] 徐豹, 莊炳昌, 路琴華, 王玉民, 胡傳璞, 梁歧, 鄭惠玉, 呂景良. 中國野生大豆()脂肪及其脂肪酸組成的研究. 吉林農業科學, 1993, (2): 1–6. Xu B, Zhuang B C, Lu Q H, Wang Y M, Hu C P, Liang Q, Zheng H Y, Lyu J L. A study of fat content and fatty acid composition of wild soybean () in China., 1993, (2): 1–6 (in Chinese with English abstract).
[19] 王連錚, 吳和禮, 姚振純, 林紅. 黑龍江省野生大豆的考察和研究. 植物研究, 1983, 3(3): 116–130. Wang L Z, Wu H L, Yao Z C, Lin H. Investigation and research of the wild soybean in Heilongjiang province., 1983, 3(3): 116–130 (in Chinese with English abstract).
[20] 姚利坡. 野生大豆與栽培大豆遺傳關系研究. 河北師范大學碩士學位論文, 河北石家莊, 2009. Yao L P. Studies on the Genetic Relationship betweenand. MS Thesis of Hebei Normal University, Shijiazhuang, Hebei, China, 2009 (in Chinese with English abstract).
[21] 趙洪錕, 王玉民, 李啟云, 張明, 莊炳昌. 中國不同緯度野生大豆和栽培大豆SSR分析. 大豆科學, 2001, 20: 172–720. Zhao H K, Wang Y M, Li Q Y, Zhang M, Zhuang B C. SSR analysis of wild soybean () and cultivated soybean from different latitude in China., 2001, 20: 172–176 (in Chinese with English abstract).
[22] 王金陵. 大豆的進化與其分類栽培及育種的關系. 中國農業科學, 1962, 3(1): 11–15.Wang J L. The evolution of soybean and its relationship with cultivating and breeding., 1962, 3(1): 11–15 (in Chinese).
[23] Han Y P, Zhao X, Liu D Y, Li Y H, Lightfoot D A, Yang Z J, Zhao L, Zhou G, Wang Z K, Huang L, Zhang Z W, Qiu L J, Zheng H K, Li W B. Domestication footprints anchor genomic regions of agronomic importance in soybeans., 2016, 209: 871–884.
[24] 宋萬坤, 于妍, 高運來, 姜威, 劉春燕, 孫殿君, 陳慶山, 胡國華. 大豆飽和脂肪酸組分改良研究進展. 生物技術通報, 2008, (增刊1): 18–21.Song W K, Yu Y, Gao Y L, Jiang W, Liu C Y, Sun D J, Chen Q S, Hu G H. Genetic modification of saturated fatty acid composition of soybean., 2008, (suppl-1): 18–21 (in Chinese with English abstract).
[25] 吳俏槿, 杜冰, 蔡尤林, 梁鉆好, 林志光, 邱國亮, 董立軍.a-亞麻酸的生理功能及開發研究進展. 食品工業科技, 2016, 37(10): 386–390.Wu Q J, Du B, Cai Y L, Liang Z H, Lin Z G, Qiu G L, Dong L J. Research development of alpha-linolenic acid., 2016, 37(10): 386–390 (in Chinese with English abstract).
Characteristics of oil components and its relationship with domestication of oil components in wild and cultivated soybean accessions
CHEN Ying**, ZHANG Sheng-Rui**, WANG Lan, WANG Lian-Zheng, LI Bin*, and SUN Jun-Ming*
Institute of Crop Sciences, Chinese Academy of Agricultural Sciences / National Engineering Laboratory for Crop Molecular Breeding / Key Laboratory of Soybean Biology, the Ministry of Agriculture and Rural Affairs, Beijing 100081, China
In this study, 58 soybean accessions composed of wild, semi-wild and cultivated soybean were used to analyze the oil and fatty acid contents by the NIRs and GC methods. Their genetic diversity and domestication were also analyzed based on 32 pairs of SSR markers. There was a significant difference in oil content and fatty acid compositions between wild and cultivated soybeans. The oil content of cultivated soybean (an average of 20.8%) was significantly higher than that of wild soybean (an average of 10.49%). As regards fatty acid compositions, the content of oleic acid in cultivated soybean (an average of 28.5%) was significantly higher than that of wild soybean (an average of 14.37%), on the contrary, the content of linolenic acid was lower. In cultivated soybean, the oil content positively correlated with oleic acid content (= 0.85**), and negatively correlated with other fatty acids. Moreover, the oleic acid content had a negative correlation with all other fatty acids, especially, with linoleic acid (=-0.90**) and linolenic acid (=-0.89**). Similar clustering results were observed in the clustering and principal component analysis based on oil compositions and SSR molecular markers showing that the soybean accessions were clustered into two main subgroups of wild and cultivated soybean, and semi-wild soybean distributed into both subgroups. Therefore, we suggest that contents of oil and fatty acid compositions are related with the domestication level in soybean, and may be used as a reference index for the evolutionary classification in soybean.
;;; oil content; fatty acid components
2018-08-24;
2019-01-19;
2019-03-11.
10.3724/SP.J.1006.2019.84114
李斌, E-mail: libin02@caas.cn; 孫君明, E-mail: sunjunming@caas.cn
**同等貢獻(Contributed equally to this work)
E-mail: 623882855@qq.com
本研究由國家重點研發計劃項目(2016YFD0100201), 北京市科技計劃項目(Z16110000916005)和中國農業科學院科技創新工程項目資助。
This study was supported by the National of Research and Development Program of China (2016YFD0100201), the Beijing Science and Technology Project (Z16110000916005), and the Innovation Project of Chinese Academy of Agricultural Sciences.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20190308.1044.006.html
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
