控制高粱分蘗與主莖株高一致性的基因定位
2019-05-31 06:45:38詹鵬杰于紀(jì)珍楚建強(qiáng)平俊愛張福耀
作物學(xué)報(bào) 2019年6期
關(guān)鍵詞:關(guān)聯(lián)
王 瑞 凌 亮 詹鵬杰 于紀(jì)珍 楚建強(qiáng) 平俊愛,*張福耀,*
1山西省農(nóng)業(yè)科學(xué)院高粱研究所 / 高粱遺傳與種質(zhì)創(chuàng)新山西省重點(diǎn)實(shí)驗(yàn)室, 山西榆次 030600; 2山西省農(nóng)業(yè)科學(xué)院食用菌研究所, 山西太原030031
高粱[Sorghum bicolor(L.) Moench]是全球農(nóng)業(yè)生態(tài)系統(tǒng)中重要的糧食和飼料作物。全球年種植面積 4000多萬(wàn)公頃, 總產(chǎn)約 6000萬(wàn)噸。高粱抗旱且耐鹽堿和瘠薄土壤, 具有在惡劣環(huán)境下生長(zhǎng)的能力,被視為干旱和鹽堿土壤農(nóng)業(yè)區(qū)農(nóng)業(yè)可持續(xù)發(fā)展的一種重要作物[1-3]。隨著對(duì)淡水資源需求的增加、邊際農(nóng)田利用的增多和全球氣候變暖, 耐干旱、鹽堿作物——高粱將在全球糧食需求不斷增加的今天顯的越來(lái)越重要。高粱也是我國(guó)的重要旱糧作物之一,全國(guó)年種植面積約80萬(wàn)公頃。目前, 我國(guó)的農(nóng)業(yè)生產(chǎn)進(jìn)入新的轉(zhuǎn)型期, 勞動(dòng)力成本逐年提高, 農(nóng)民對(duì)機(jī)械化栽培高粱的渴求與日俱增, 但推廣的高粱品種基本不適宜機(jī)械化栽培, 一是株高本身偏高, 不適宜機(jī)械化作業(yè), 二是分蘗高度與主莖不一致, 高粱穗不在一個(gè)層面, 造成收獲時(shí)丟損嚴(yán)重, 極大地制約了高粱機(jī)械化生產(chǎn)的發(fā)展[4-5]。因此, 高粱分蘗和主莖株高一致的研究, 對(duì)指導(dǎo)高粱育種和選育適宜機(jī)械化生產(chǎn)高粱品種具有十分重要的理論意義和應(yīng)用價(jià)值。
國(guó)內(nèi)外對(duì)機(jī)械化生產(chǎn)高粱品種的研究主要集中在株高和分蘗力這2個(gè)性狀。Quinby等[6]研究確定了植株高度由4個(gè)基因位點(diǎn)控制, 依此命名為Dwl、Dw2、Dw3和Dw4; Pereira 等[7]以 CK60 和 PI229828為親本雜交得到 F2群體, 計(jì)算出 4個(gè)控制株高的QTL; Lin等[8]以雜交F2代群體鑒定出6個(gè)控制株高的QTL; Rami等[9]以IS2807分別與種質(zhì)庫(kù)中2種高粱雜交, 得到2個(gè)RIL群體, 以此為研究對(duì)象, 得到3個(gè)與株高相關(guān)的 QTL; Klein等[10]以 Sureno和RTx430為親本雜交, 選取125株F2鑒定得到2個(gè)控制株高性狀的QTL; Upadhyaya等[11]用242份微核心種質(zhì)與39個(gè)標(biāo)記關(guān)聯(lián)分析, 得出5個(gè)標(biāo)記與熟期或株高關(guān)聯(lián); Upadhyaya等[12]開發(fā)了14,739個(gè)SNP標(biāo)記評(píng)估株高, 結(jié)果發(fā)現(xiàn), 6個(gè) SNP位點(diǎn)與株高關(guān)聯(lián);Harris-Shultz等[13]以 F2群體發(fā)現(xiàn)了 1個(gè)新的株高QTL位于第 3染色體上; 蘇舒等[14]以 T70和 P607為親本, 雜交得到F6代RIL檢測(cè)到6個(gè)控制株高的QTL; 劉娟等[15]利用‘河農(nóng) 16’與‘千三’的 F2:3遺傳作圖群體的后代材料建立的F8代的132個(gè)重組自交系, 檢測(cè)到2個(gè)與株高相關(guān)的QTL。分蘗數(shù)表現(xiàn)出數(shù)量性狀的遺傳特點(diǎn), 受多個(gè)數(shù)量基因位點(diǎn)和環(huán)境因子的共同作用[16]。Feltus等[17]利用BTx623 ×IS3620C的F6-8代作為構(gòu)圖群體, 檢測(cè)到9個(gè)控制高粱分蘗數(shù)的 QTL; Shehzad等[18]對(duì)來(lái)自亞洲和非洲27個(gè)國(guó)家的代表性高粱品種多樣性研究發(fā)現(xiàn)7個(gè)影響分蘗數(shù)的 QTL; Shiringani等[19]利用甜高粱 SS79和粒用高粱M71雜交得到的F6代重組自交系群體為定位群體, 檢測(cè)到 6個(gè)影響高粱分蘗數(shù)的 QTL; 董維等[20]以T70 × P607雜交得到的F6代RIL群體構(gòu)建遺傳連鎖圖, 檢測(cè)到7個(gè)與分蘗數(shù)相關(guān)的QTL。對(duì)高粱分蘗和主莖株高一致的研究尚未見報(bào)道。本研究通過(guò)雜交、自交等方法構(gòu)建 F2分離群體, 基于SLAF-seq技術(shù), 利用F2遺傳分離群體和集群分離法(BSA)對(duì)高粱分蘗與主莖株高一致性狀進(jìn)行關(guān)聯(lián)分析, 定位該基因在染色體上的位置, 以期提高我國(guó)適宜機(jī)械化高粱品種選育水平以及選擇的準(zhǔn)確性和科學(xué)性。
1 材料與方法
1.1 材料
本實(shí)驗(yàn)田間部分在山西省農(nóng)業(yè)科學(xué)院高粱研究所試驗(yàn)基地(山西榆次和海南三亞)進(jìn)行, 選用矮桿多分蘗且分蘗整齊一致的品種 K35-Y5為母本, 中高桿有分蘗且分蘗明顯高于主莖的恢復(fù)系1383為父本雜交得F1代, 冬季在海南三亞種植F1代, 第 2年夏在山西榆次種植 F2代, 株行距為 25 cm × 50 cm, 每公頃留苗約8萬(wàn)株, 保證每個(gè)單株充分表達(dá)其性狀。開花后調(diào)查分蘗與主莖株高的一致性, 分蘗株高與主莖株高同等或株高差≤ 5 cm, 記為株高一致, 分蘗株高與主莖株高差> 5 cm記為不一致。
選取F2分離群體中分蘗與主莖株高一致和分蘗明顯高于主莖的植株各45株, 構(gòu)建分蘗與主莖株高一致與分蘗明顯高于主莖的 2個(gè)極端性狀混池, 連同2個(gè)親本作為定位群體進(jìn)行關(guān)聯(lián)分析。
1.2 SLAF-seq文庫(kù)構(gòu)建
選擇已經(jīng)測(cè)序完成的高粱基因組作為參考基因組 ( 下 載 地 址 : ftp://ftp.ensemblgenomes.org/pub/plants/release-25/fasta/sorghum_bicolor/, 組裝出的基因組大小738.61 Mb, GC含量為43.93%), 根據(jù)基因組大小以及 GC含量等信息作為參考基因組進(jìn)行酶切預(yù)測(cè), 根據(jù)選定的最適酶切方案, 對(duì)檢測(cè)合格的各樣品基因組DNA分別進(jìn)行酶切。對(duì)得到的酶切片段(SLAF標(biāo)簽)進(jìn)行3′端加A處理、連接Dual-index[21]測(cè)序接頭、PCR擴(kuò)增、純化、混樣、切膠選取目的片段, 文庫(kù)質(zhì)檢合格后用Illumina HiSeq 2500測(cè)序。為評(píng)估酶切實(shí)驗(yàn)的準(zhǔn)確性, 選用水稻(Oryza sativa)[22]作為對(duì)照進(jìn)行測(cè)序。
1.3 SNP位點(diǎn)的開發(fā)與關(guān)聯(lián)分析
利用Dual-index對(duì)測(cè)序得到的原始數(shù)據(jù)進(jìn)行識(shí)別, 得到各個(gè)樣品的 reads。過(guò)濾測(cè)序 reads的接頭后, 進(jìn)行測(cè)序質(zhì)量和數(shù)據(jù)量的評(píng)估。通過(guò)Control數(shù)據(jù)評(píng)估酶切效率, 以此判斷實(shí)驗(yàn)過(guò)程的準(zhǔn)確性和有效性。通過(guò)將reads與參考基因組比對(duì), 在親本和混池中開發(fā) SLAF標(biāo)簽, 尋找在親本中存在多態(tài)性的SLAF標(biāo)簽和有 reads覆蓋區(qū)域的 SNP。將得到的SNP利用 SNP-index方法[23]和ED方法[24]進(jìn)行關(guān)聯(lián)分析, 獲得與性狀緊密關(guān)聯(lián)的位點(diǎn)。
1.4 關(guān)聯(lián)分析方法
1.4.1 SNP-index方法原理 SNP-index是通過(guò)混池間的基因型頻率差異進(jìn)行標(biāo)記關(guān)聯(lián)分析的方法[23],主要是尋找混池之間基因型頻率的顯著差異。
利用兩親本的 SNP數(shù)據(jù), 分別計(jì)算兩混池的SNP-index, 并通過(guò) ΔSNP-index觀測(cè)可能與性狀分離相關(guān)的位點(diǎn)。
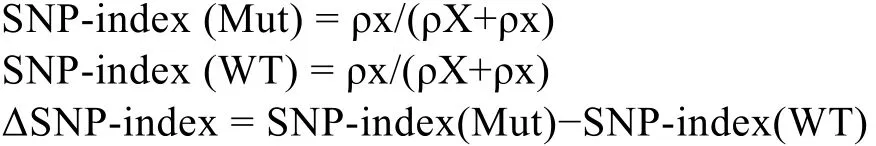
式中, Mut和WT分別為子代的突變池與野生池, ρX和ρx分別為野生型親本的等位基因, 以及突變型親本的等位基因在各自池中出現(xiàn)的 read數(shù)目。通過(guò)ΔSNP-index可以觀察每個(gè)位點(diǎn)在突變池與野生池之間的差異。
為了消除假陽(yáng)性的位點(diǎn), 利用標(biāo)記在基因組上的位置, 對(duì)同一條染色體上標(biāo)記的ΔSNP-index值進(jìn)行擬合, 并根據(jù)關(guān)聯(lián)閾值, 選擇閾值以上的區(qū)域作
為與性狀相關(guān)的區(qū)域。
1.4.2 歐氏距離方法原理 歐式距離(Euclidean distance, ED)是利用測(cè)序數(shù)據(jù)尋找混池間存在的顯著差異標(biāo)記, 并以此評(píng)估與性狀關(guān)聯(lián)區(qū)域的方法[24]。ED值越大, 表明該標(biāo)記在兩混池間的差異越大。
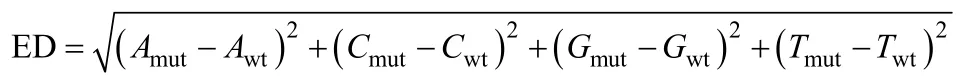
式中,Amut、Cmut、Gmut和Tmut分別為 A、C、G和 T堿基在突變混池中的頻率,Awt、Cwt、Gwt和Twt分別為A、C、G和T堿基在野生型混池中的頻率。
利用兩混池間基因型存在差異的SNP位點(diǎn), 統(tǒng)計(jì)各個(gè)堿基在不同混池中的深度, 并計(jì)算每個(gè)位點(diǎn)ED值, 為消除背景噪音, 對(duì)原始 ED值進(jìn)行乘方處理[24]。
為了消除假陽(yáng)性的位點(diǎn), 利用標(biāo)記在基因組上的位置, 對(duì)同一條染色體上標(biāo)記的ED值進(jìn)行擬合[24],并根據(jù)關(guān)聯(lián)閾值, 選擇閾值以上的區(qū)域作為與性狀相關(guān)的區(qū)域。
2 結(jié)果與分析
2.1 親本及群體分蘗高度的遺傳
K35-Y5是從美國(guó)引進(jìn)的種質(zhì), 通過(guò)在山西和海南多年種植鑒定, 表現(xiàn)為矮桿多分蘗且分蘗整齊一致, 是適宜機(jī)械化生產(chǎn)品種選育的優(yōu)良親本系。1383是我國(guó)主干高粱雜交種‘晉雜 12號(hào)’的恢復(fù)系, 中高桿有分蘗且明顯高于主莖, 由它配制的晉雜12號(hào)在生產(chǎn)中一直表現(xiàn)分蘗高、弱苗高、整齊度差。
選用K35-Y5與1383雜交, F1植株表現(xiàn)為有分蘗且分蘗比主莖高, 說(shuō)明分蘗與主莖株高一致性狀為隱形, 而分蘗高于主莖性狀為顯性。F2分離群體中分蘗與主莖株高不一致與一致的分離比例為239∶72, 經(jīng)卡方(χ2)檢驗(yàn), χ2=0.57<χ20.05=3.84, 分離符合 3∶1, 說(shuō)明分蘗與主莖株高一致性狀受一對(duì)隱性核基因控制。
2.2 酶切方案與建庫(kù)評(píng)估
對(duì)高粱參考基因組序列進(jìn)行電子酶切預(yù)測(cè), 確定限制性內(nèi)切酶為RsaI +HaeIII, 酶切片段長(zhǎng)度在364~414 bp的序列定義為 SLAF標(biāo)簽, 預(yù)測(cè)可得到103,902個(gè)SLAF標(biāo)簽。統(tǒng)計(jì)SLAF標(biāo)簽在各染色體上的數(shù)量(表1), 繪制SLAF標(biāo)簽在染色體上的分布圖(圖1)。由圖可知, SLAF標(biāo)簽在基因組各染色體上分布基本均勻, 酶切方案可行。
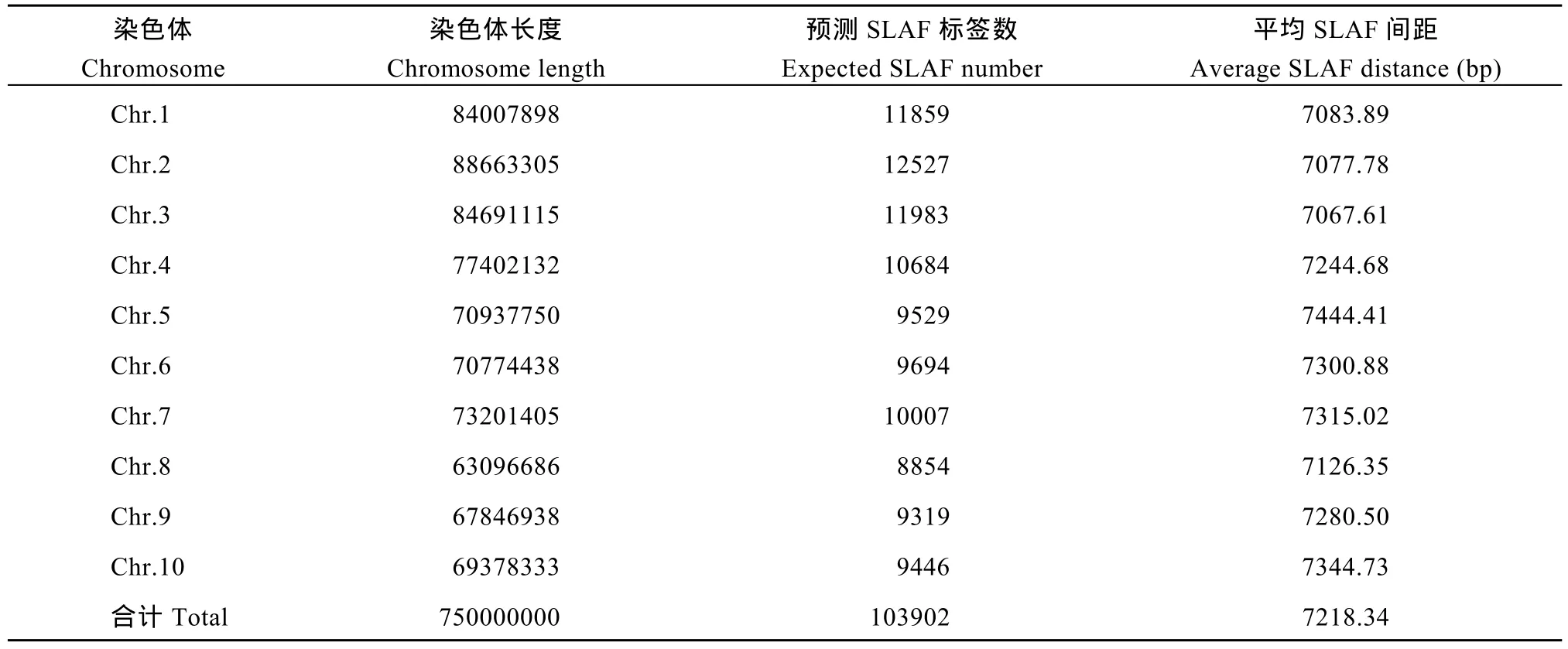
表1 SLAF標(biāo)簽在各染色體上的數(shù)量統(tǒng)計(jì)Table 1 Number of SLAF labels on each chromosome
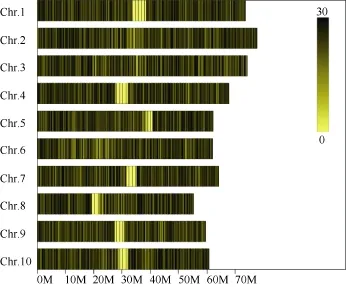
圖1 SLAF標(biāo)簽在參考基因組各染色體上的分布Fig. 1 Distribution of SLAF on reference genome
為進(jìn)一步評(píng)估酶切方案的有效性, 以水稻(Oryza sativa)作對(duì)照, 其基因組大小為374.31 Mb (下載地址:http://rice.plantbiology.msu.edu), 通過(guò) BWA[25]軟件將對(duì)照的測(cè)序 reads與參考基因組比對(duì)(表 2和圖 2)顯示, 本次實(shí)驗(yàn)雙端比對(duì)效率在 93.35%, 酶切效率為 90.60%, 對(duì)照測(cè)序 reads插入片段的長(zhǎng)度均集中分布在360~410 bp之間, 說(shuō)明SLAF建庫(kù)正常。
2.3 測(cè)序數(shù)據(jù)統(tǒng)計(jì)與評(píng)估
采用100 bp×2作為后續(xù)使用的數(shù)據(jù), 進(jìn)行數(shù)據(jù)評(píng)估和分析。測(cè)序質(zhì)量值(Q)是評(píng)估高通量測(cè)序單堿基錯(cuò)誤率的重要指標(biāo), 測(cè)序質(zhì)量值越高對(duì)應(yīng)的堿基測(cè)序錯(cuò)誤率越低。共獲得30.80 M reads數(shù)據(jù), 測(cè)序平均Q30為91.70%, 平均GC含量為45.79% (表3)。用于評(píng)估實(shí)驗(yàn)建庫(kù)的準(zhǔn)確性的水稻(Control)測(cè)序獲得0.19 M reads的數(shù)據(jù)量。
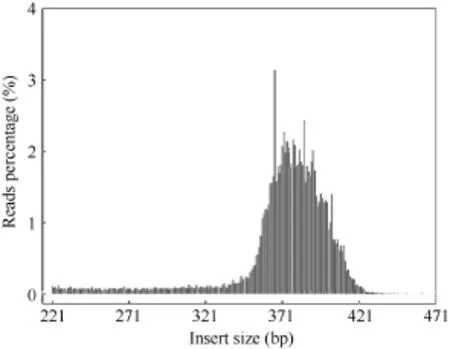
圖2 對(duì)照reads插入片段分布Fig. 2 Distribution of observed control insert size

表2 水稻測(cè)序reads比對(duì)分析Table 2 Alignment analysis of obtained reads in Oryza sativa
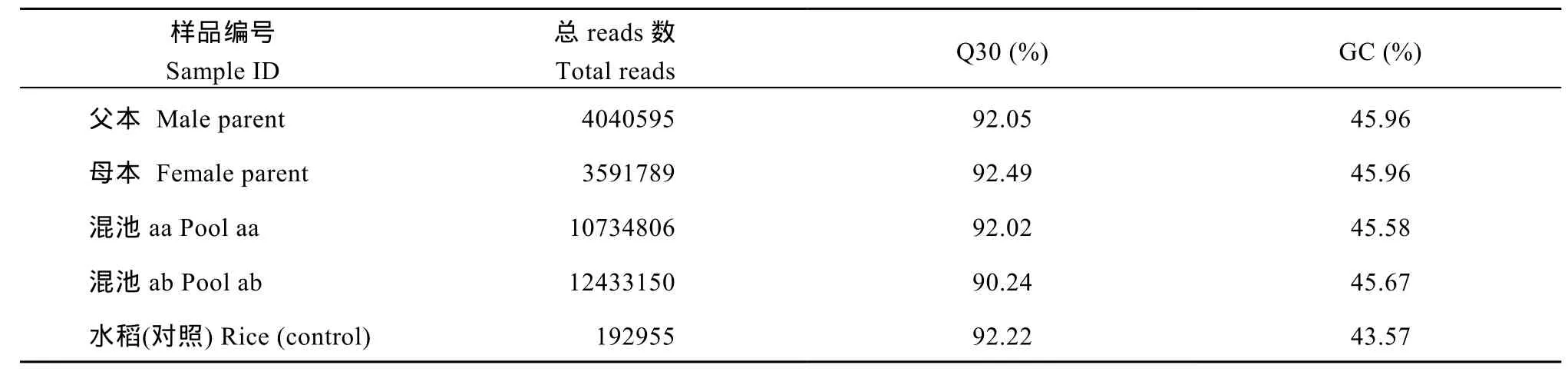
表3 各樣品測(cè)序數(shù)據(jù)統(tǒng)計(jì)表Table 3 Statistics of sequencing for each sample
2.4 SLAF標(biāo)記開發(fā)
利用參考基因組共開發(fā)133,246個(gè)SLAF標(biāo)簽,SLAF標(biāo)簽親本平均測(cè)序深度為 22.68×, 混池平均測(cè)序深度為 61.94× (表 4); SNP 的檢測(cè)主要使用GATK軟件工具包實(shí)現(xiàn), 所有樣品的SNP統(tǒng)計(jì)信息見表5。統(tǒng)計(jì)不同染色體上的SLAF標(biāo)簽與SNP標(biāo)記的分布(表 6); 根據(jù) SLAF在染色體上的分布, 繪制SLAF標(biāo)簽和多態(tài)性SLAF標(biāo)簽的染色體分布圖,由圖3可以看出, 開發(fā)的SLAF標(biāo)記分布較均勻。
2.5 關(guān)聯(lián)分析
2.5.1 SNP-index方法關(guān)聯(lián)結(jié)果 在 SNP-index關(guān)聯(lián)分析前, 先從319,428個(gè)SNP中, 過(guò)濾掉有多重突變的SNP位點(diǎn)、混池中read支持度小于4的位點(diǎn)、親本中不存在的 SNP位點(diǎn), 最終獲得 59,407個(gè)SNP。利用 SNP-index方法[23]計(jì)算關(guān)聯(lián)值, 并采用SNPNUM方法對(duì)ΔSNP-index進(jìn)行擬合, 取每個(gè)SNP附近200個(gè)SNP的ΔSNP-index的中值作為該位點(diǎn)擬合后的關(guān)聯(lián)值。兩個(gè)混池分別的 SNP-index及ΔSNP-index的分布如圖4所示:
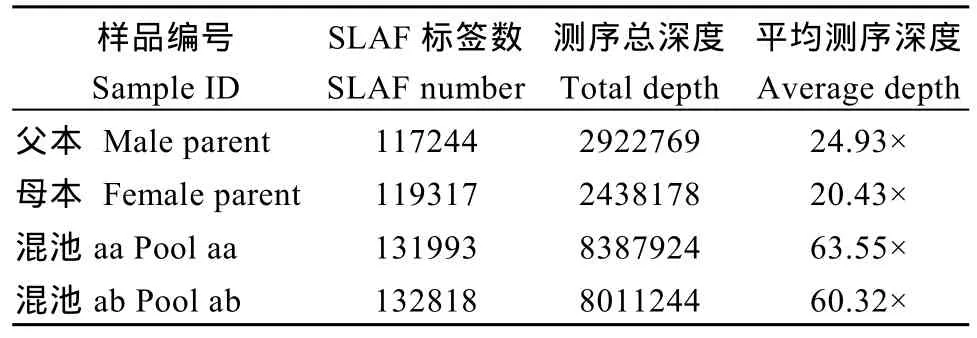
表4 SLAF標(biāo)簽統(tǒng)計(jì)Table 4 Statistics of SLAF
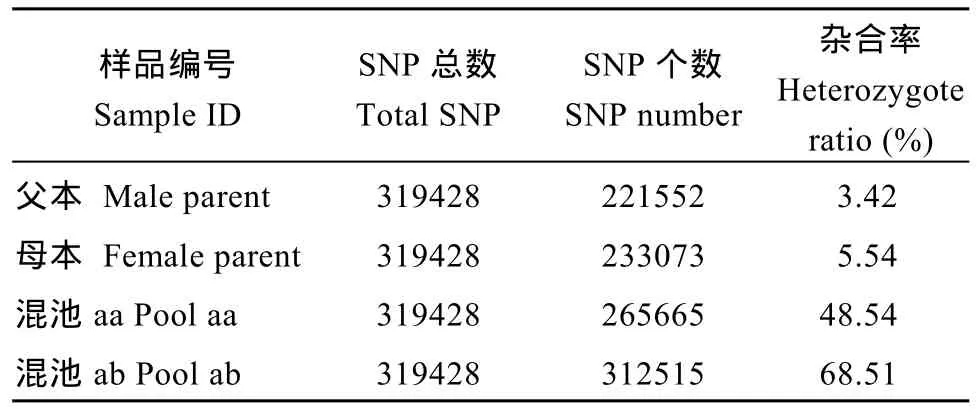
表5 SNP信息統(tǒng)計(jì)Table 5 Statistics of SNP information
根據(jù)計(jì)算機(jī)模擬實(shí)驗(yàn)[26]計(jì)算結(jié)果, 當(dāng)置信度為0.90時(shí), 定位區(qū)域在第 9染色體上的 54,788,026~56,740,873 (1.95 M)區(qū)間內(nèi)(表 7)。
2.5.2 ED方法關(guān)聯(lián)結(jié)果 在ED法關(guān)聯(lián)分析前,先從319,428個(gè)SNP中過(guò)濾任一混池中read支持度小于 4的位點(diǎn) 212,186個(gè), 得到高質(zhì)量的可信SNP位點(diǎn)共107,242個(gè), 并在此基礎(chǔ)上識(shí)別兩混池間差異的位點(diǎn)共100,261個(gè)。利用ED方法[24]計(jì)算關(guān)聯(lián)值, 并取原始ED的3次方作為關(guān)聯(lián)值以達(dá)到消除背景噪音的功能, 然后采用局部線性回歸LOESS方法對(duì) ED值進(jìn)行擬合, 關(guān)聯(lián)值分布見圖 5。
取所有位點(diǎn)擬合值的 median+3SD作為分析的關(guān)聯(lián)閾值[24], 計(jì)算得 0.0457, 根據(jù)關(guān)聯(lián)閾值判定,定位區(qū)域在第 9染色體的 30,964,929~38,410,608(7.45 M)和 43,824,552~59,633,348 (15.81 M)區(qū)間內(nèi)(表 8)。
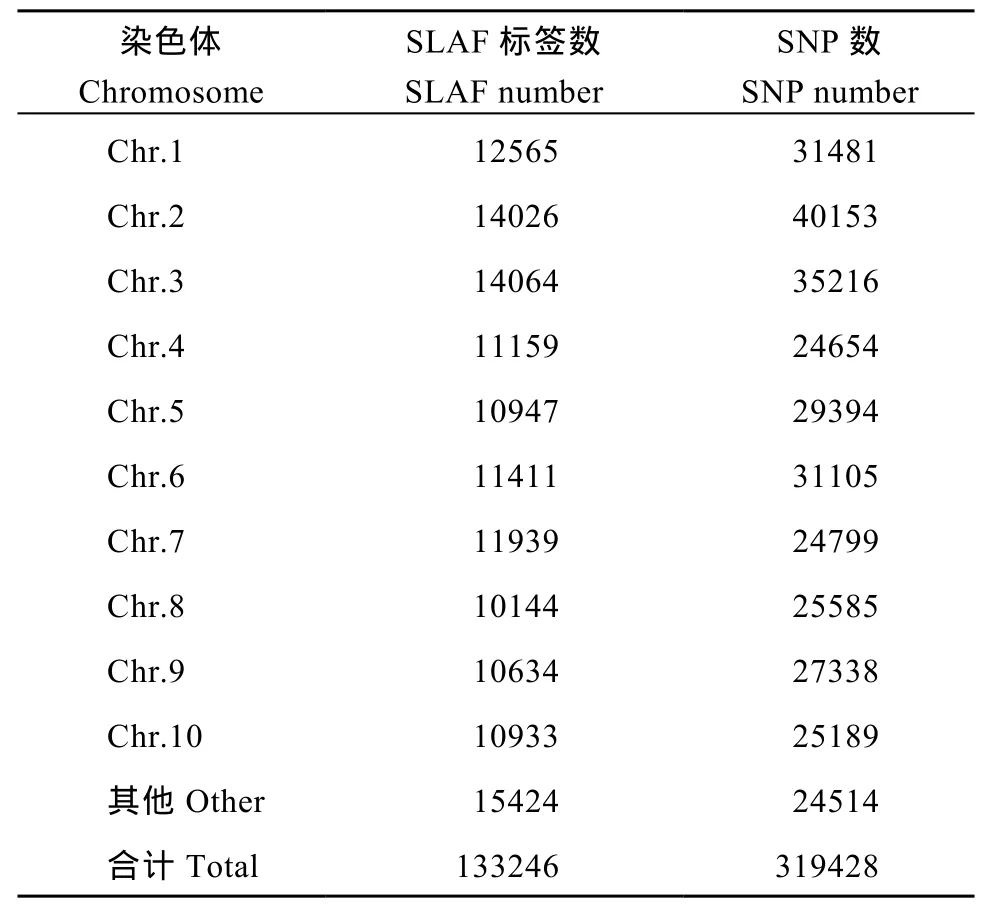
表6 SLAF標(biāo)簽和SNP標(biāo)記在染色體上的分布統(tǒng)計(jì)Table 6 Distribution statistics of SLAF and SNP on genome
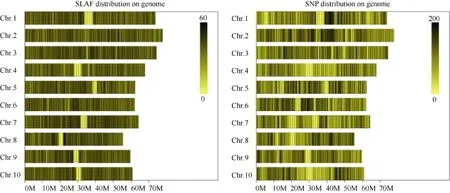
圖3 SLAF標(biāo)簽和SNP標(biāo)記在染色體上的分布Fig. 3 Distribution of SLAF and SNP on genome
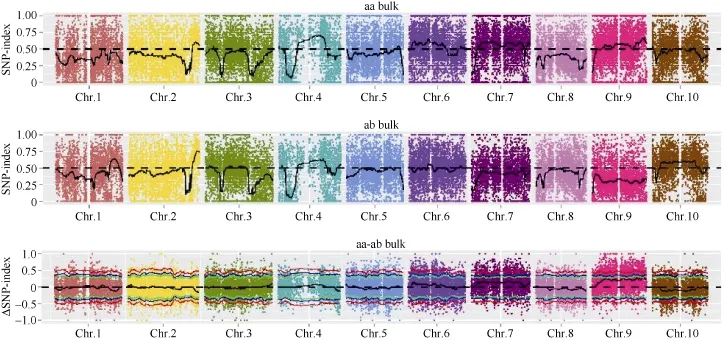
圖4 SNP-index關(guān)聯(lián)值在染色體上的分布Fig. 4 Distribution of SNP-index associated values on genome
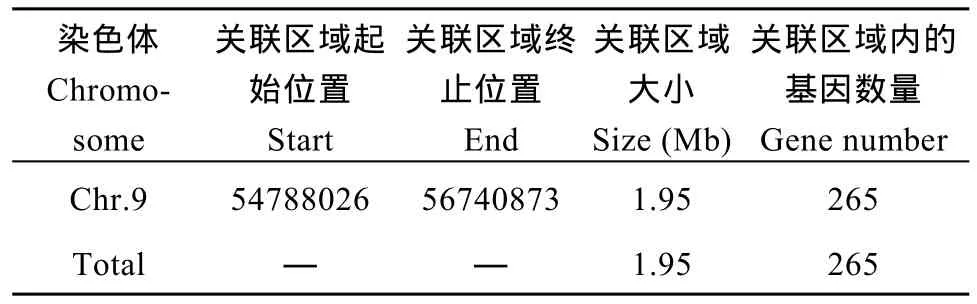
表7 關(guān)聯(lián)區(qū)域信息統(tǒng)計(jì)表Table 7 Statistics of associated region information
2.5.3 候選關(guān)聯(lián)區(qū)域篩選與功能分析 對(duì)這2種方法取交集, 以期得到更準(zhǔn)確的結(jié)果, 得到的交集是定位區(qū)域在第 9染色體上的 54,788,026~56,740,873區(qū)間內(nèi), 該關(guān)聯(lián)區(qū)域長(zhǎng)度1.95 Mb, 在關(guān)聯(lián)區(qū)域內(nèi)共有 265個(gè)候選基因。針對(duì)關(guān)聯(lián)區(qū)域內(nèi)的基因, 分析在外顯子區(qū)域兩個(gè)親本之間差異的 SNP信息, 對(duì) SNP進(jìn)行變異的注釋(表 9), 共發(fā)現(xiàn)存在非同義突變的 SNP4個(gè), 對(duì)應(yīng)到Sobic.009G197901.1、Sobic.009G213300.1和Sobic.009G221200.1三個(gè)基因,初步確定這些基因是與性狀直接相關(guān)的功能基因。
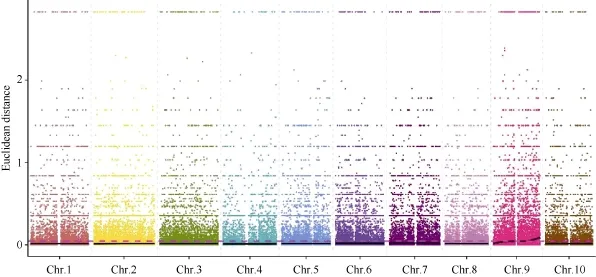
圖5 ED關(guān)聯(lián)值在染色體上的分布Fig. 5 Distribution of ED associated values on genome
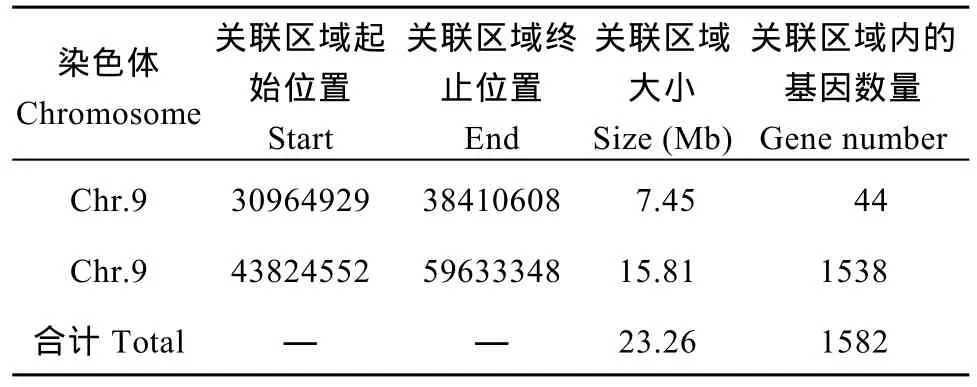
表8 關(guān)聯(lián)區(qū)域信息統(tǒng)計(jì)表Table 8 Statistics of associated region information
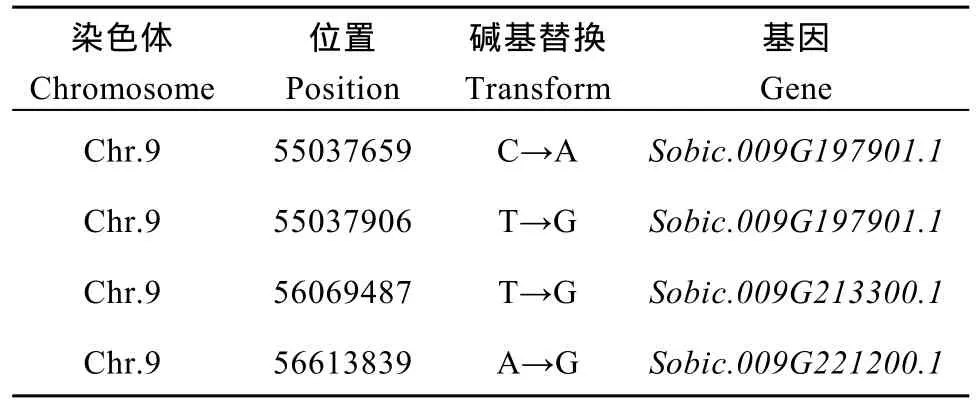
表9 SNP標(biāo)記信息Table 9 Information of SNP
2.6 相關(guān)連鎖分子標(biāo)記的開發(fā)及驗(yàn)證
在4個(gè)非同義突變的SNP兩端設(shè)計(jì)引物(表10),對(duì)雙親的基因組 DNA進(jìn)行 PCR擴(kuò)增, 用瓊脂糖凝膠電泳檢測(cè)擴(kuò)增產(chǎn)物。將擴(kuò)增產(chǎn)物回收, 測(cè)序; 對(duì)測(cè)序結(jié)果進(jìn)行多序列對(duì)比, 獲得基因序列特異差異(圖6)。分蘗與主莖株高不一致親本1383和分蘗與主莖株高一致親本K35-Y5在第55,037,659 bp處分別為A和C; 在第 55,037,906 bp處分別為 G和 T; 在第56,069,487 bp處分別為G和T; 在第56,613,839 bp處分別為G和A。說(shuō)明這4個(gè)SNP是與分蘗與主莖株高一致性狀直接相關(guān)的。
3 討論
對(duì)高粱分蘗性已有廣泛研究, 影響分蘗性的主要因素有水肥條件、溫度、密度等[27]。水肥條件充分, 分蘗期早且長(zhǎng), 分蘗早且多; 不同品種分蘗性具有隨播種至分蘗期土壤溫度的升高而降低的趨勢(shì),溫度高, 生長(zhǎng)快, 迅速進(jìn)入拔節(jié)期, 縮短分蘗時(shí)期;在北方地區(qū), 隨著種植密度的增加單株分蘗數(shù)明顯減少, 高粱分蘗性受環(huán)境影響較大。但高粱分蘗與主莖的高度差這一性狀基本不受環(huán)境的影響, 分蘗與主莖高度一致這一性狀遺傳穩(wěn)定, 已在適宜高粱機(jī)械化育種中應(yīng)用, 育成一批適宜機(jī)械化生產(chǎn)的高粱品種[28]。Upadhyaya等[11]用 242份微核心種質(zhì)與39個(gè)標(biāo)記關(guān)聯(lián)分析, 得出5個(gè)標(biāo)記與熟期或株高關(guān)聯(lián), 分別位于第 6、第 9、第 10染色體上。Brown等[29]用包含119個(gè)單株的RIL群體鑒定得到與株高相關(guān)的4個(gè)QTL, 分別位于第3、第6、第7、第9染色體上。Shiringani等[30]建立了包含188個(gè)單株的RIL群體, 利用 157對(duì)引物, 得到 5個(gè)關(guān)于株高的QTL, 分別位于第1、第2、第5、第6、第9染色體上。Feltus等[17]利用 BTx623×IS3620C 的 F6-8代作為構(gòu)圖群體, 共檢測(cè)到 9個(gè)控制高粱分蘗數(shù)的 QTL,多數(shù)集中于第1和第6染色體上, 第5、第7和第9染色體上各檢測(cè)到1個(gè)QTL。不同的研究者研究結(jié)果不盡相同, 但大多研究結(jié)果都關(guān)聯(lián)到第9染色體。本研究對(duì)高粱分蘗與主莖株高一致性基因的定位與前人研究結(jié)果一致, 所定位到的基因也位于高粱第9染色體上, 說(shuō)明高粱第9染色體不僅存在分蘗性狀的基因, 而且還有分蘗與主莖株高一致性的基因。

表10 引物序列和信息Table 10 Sequence and information of the primers

圖6 引物SNP1(A)、SNP2(B)、SNP3(C)、SNP4(D)對(duì)雙親的測(cè)序結(jié)果Fig. 6 Sequencing results of the parents with primer SNP1(A), SNP2(B), SNP3(C), and SNP4(D)
基于 SLAF-seq技術(shù)的基因定位, 是集 BSA技術(shù)、生物信息學(xué)和高通量測(cè)序技術(shù)為一體的一種簡(jiǎn)單高效的基因定位方法。目前, 利用此類方法定位的有番茄雄性不育基因[31], 甘藍(lán)型油菜抗霜霉病基因[32], 油菜與種子重量高度相關(guān)的基因[33]等。本研究利用SLAF-seq技術(shù), 將高粱分蘗與主莖株高一致基因定位于第 9染色體上的 54,788,026~56,740,873區(qū)間內(nèi), 關(guān)聯(lián)區(qū)域長(zhǎng)度1.95 Mb。
4 結(jié)論
分蘗與主莖株高一致性狀由1對(duì)隱性核基因控制。利用SNP-index法和Euclidean distance法及取兩者交集進(jìn)行關(guān)聯(lián)分析, 得到一個(gè)關(guān)聯(lián)區(qū)域, 位于第 9染色體上的 54,788,026~56,740,873區(qū)間內(nèi), 關(guān)聯(lián)區(qū)域長(zhǎng)度1.95 Mb。在該關(guān)聯(lián)區(qū)域內(nèi)發(fā)現(xiàn)4個(gè)非同義突變的 SNP, 它們和分蘗與主莖株高一致性狀相關(guān)。對(duì)應(yīng)到Sobic.009G197901.1、Sobic.009G213300.1
和Sobic.009G221200.1三個(gè)基因上, 這些基因可能是與性狀直接相關(guān)的功能基因。通過(guò)進(jìn)一步精細(xì)定位, 可以確定該目標(biāo)基因。
猜你喜歡
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:30
當(dāng)代陜西(2021年17期)2021-11-06 03:21:36
原道(2020年2期)2020-12-21 05:47:06
當(dāng)代陜西(2019年15期)2019-09-02 01:52:00
中國(guó)非營(yíng)利評(píng)論(2018年2期)2018-06-18 10:48:50
學(xué)苑創(chuàng)造·A版(2018年11期)2018-02-01 06:29:20
自動(dòng)化學(xué)報(bào)(2017年1期)2017-03-11 17:31:17
讀者(2017年5期)2017-02-15 18:04:18
西藏科技(2016年5期)2016-09-26 12:16:39
振動(dòng)工程學(xué)報(bào)(2015年1期)2015-03-01 01:15:42
