植物三萜皂苷代謝中糖基轉移酶蛋白特性的生物信息學分析
2019-05-13 08:27:28趙志新
陜西農業科學 2019年4期
趙志新, 王 通
(商洛學院 生物醫藥與食品工程學院,陜西 商洛 726000)
三萜皂苷是植物界中一類重要的植物次生代謝產物,具有多種生物活性,其可增強植物抗病、抗蟲害的能力,同時還具有抗炎、抗腫瘤、抗凝血、抗病毒、降低膽固醇、保肝等作用[1]。目前人們已較為明確地認識了三萜皂苷的合成途徑,第一步先通過甲羥戊酸等途徑生成異戊二烯焦磷酸,再經過催化酶鯊烯環氧酶等的催化,生成2,3-氧化鯊烯環化酶,環化形成三萜類骨架,然后通過細胞色素p450、糖基轉移酶、β-糖苷酶的修飾作用,最后形成糖苷化合物-三萜皂苷[2]。糖基轉移酶作為三萜皂苷生物合成的最后一步,其機理及代謝調控機制已深入到分子水平,因此為在分子水平研究藥用植物中三萜皂苷生物合成機理起到了重要作用[3]。
糖基轉移酶(Glycosyltransferase, GT)對植物的生長發育過程和代謝工程應用方面起著重要作用,催化植物次生代謝物合成[4]。糖基轉移酶具有多種生物活性, 包括參與植物次生代謝、脫毒反應、激素平衡、信號轉導和防御反應等[5]。糖基轉移酶在植物次生代謝過程晚期對底物選擇靈活,當進化趨勢變化或外界環境變化時,可使植物能夠較快適應不同底物。最近研究顯示,植物糖基轉移酶有2個羅斯曼折疊(Rossmann fold,βαβ結構),在N末端是受體結合區,C末端為活性供體結合區;同時該結構表現出C末端保守區的功能,以及特定氨基酸殘基合成的結合糖供體的口袋結構[6]。
筆者研究利用生物信息學工具,首先搜集植物中已知的糖基轉移酶序列,然后使用生物信息學工具對其理化性質及蛋白質結構功能進行分析,為以后研究糖基轉移酶蛋白特性提供借鑒。
1 材料與方法
1.1 材料
查找植物糖基轉移酶相關文獻,并搜索NCBI數據庫,下載已收錄的植物糖基轉移酶基因序列,共獲得以下16種植物(見表1)。
1.2 方法
使用在線分析軟件ProtParam分析糖基轉移酶氨基酸序列的理化性質;SignalP4.1Server在線工具分析糖基轉移酶序列中包含的信號肽;SOPMA在線工具分析糖基轉移酶的二級結構(詳細見表2)。
2 結果與分析
2.1 糖基轉移酶氨基酸序列組成和理化性質分析
使用在線分析軟件ProtParam分析16種植物糖基轉移酶的氨基酸序列的組成和理化性質(見表3)。

表1 植物糖基轉移酶GenBank登錄號及所屬科

表2 糖基轉移酶蛋白特性分析工具及網址

表3 糖基轉移酶氨基酸序列的組成和理化性質
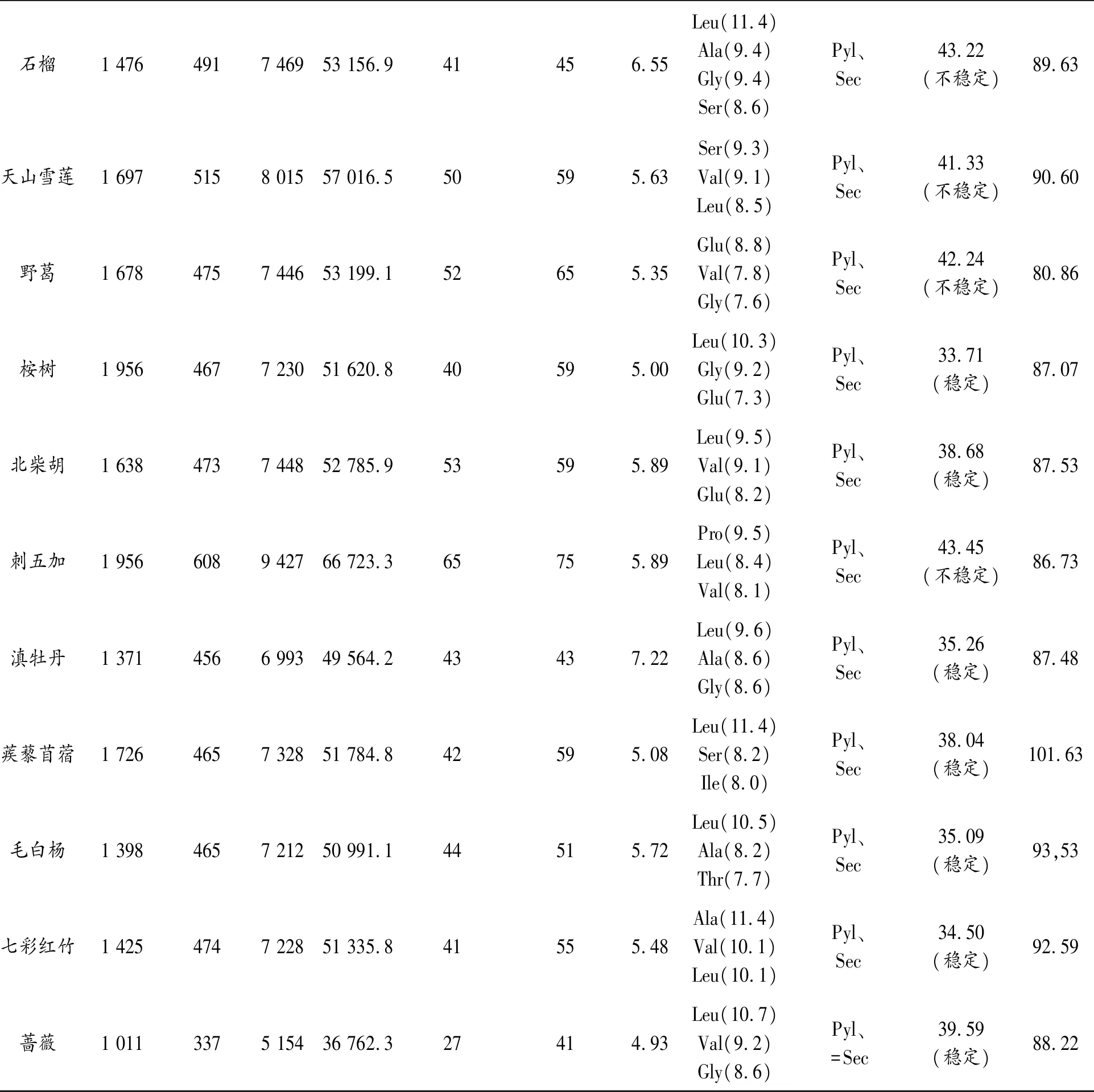
續表3 糖基轉移酶氨基酸序列的組成和理化性質
從表3中可以看出,這16種植物的基因全長均在1 000 bp到2 000 bp之間,變異較小;氨基酸數目大部分在450到500之間,除了薔薇(337)、白蘇子(447)、天山雪蓮(515)和刺五加(608);原子數除刺五加(9427)和薔薇(5154)外,均為7 000左右;分子量除薔薇外(36762.3),都在50 000至67 000之間;滇牡丹pI值為7.22,石榴為6.55,其余植物均小于6,顯示糖基轉移酶氨基酸大多為酸性氨基酸;不穩定指數在40左右,桉樹、北柴胡、滇牡丹、蒺藜苜蓿、毛白楊、七彩虹葉、薔薇和紅景天糖基轉移酶屬于穩定類蛋白,其余為不穩定類蛋白;所選植物的糖基轉移酶序列中含量最豐富的氨基酸是Leu、Val、Ser、Gly、Glu,而Sec和Pyl則是所有植物序列中均不含有的氨基酸;脂肪指數除蒺藜苜蓿(101.63)和野葛(80.86)外,均在90左右。
2.2 糖基轉移酶蛋白質信號肽的預測分析
使用SignalP4.1Server在線工具對糖基轉移酶序列中包含的信號肽進行分析,圖1和圖2分別為紅景天和三七的分析結果。

圖1 紅景天糖基轉移酶的信號肽分析結果
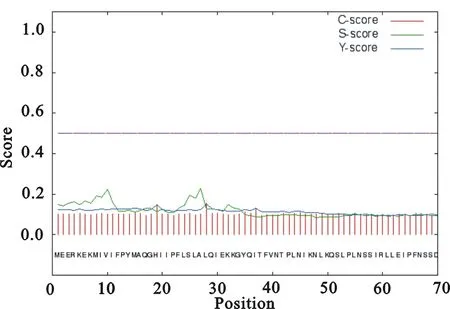
圖2 三七糖基轉移酶的信號肽分析結果
圖中每個位置的氨基酸對應一個S值,若S值較高表明該區域可能有信號肽;C值是剪切位點值,在剪切位點處C值最高;Y值是在不只有一個較高C值時,綜合考慮S值和C值的一個參
數;粉色的虛橫線為閾值線(Threshold value),在該線上方表明具有顯著性。由圖1和圖2可以看出,紅景天的糖基轉移酶序列40-50位置之間可能含有信號肽,而三七的糖基轉移酶序列則不存在信號肽。同時從圖1中可以看出,紅景天的Y值比較低,但S值較高;而三七的S值和Y值均比較低,因此可以推測三七的糖基轉移酶通過核糖體合成之后,生成的蛋白屬于非分泌蛋白。除過紅景天的糖基轉移酶蛋白經核糖體生成的蛋白可能屬于分泌蛋白外,其余14種植物信號肽分析結果到與三七相近,因此沒有展示。
2.3 糖基轉移酶二級結構預測分析
以天山雪蓮為例,使用SOPMA在線工具分析糖基轉移酶氨基酸序列的二級結構(圖3)以及各元件比例結果(表3)。
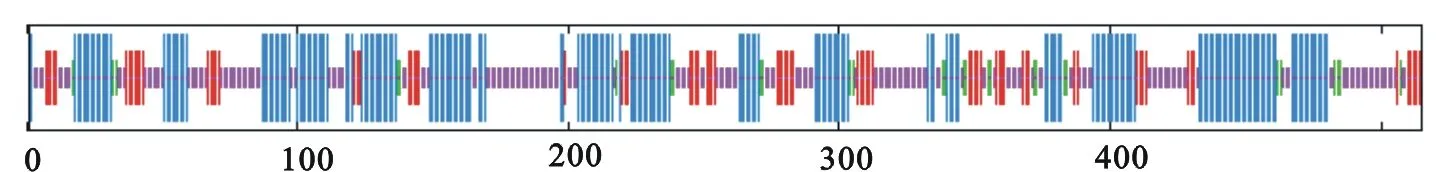
圖3天山雪蓮GT氨基酸序列的二級結構預測
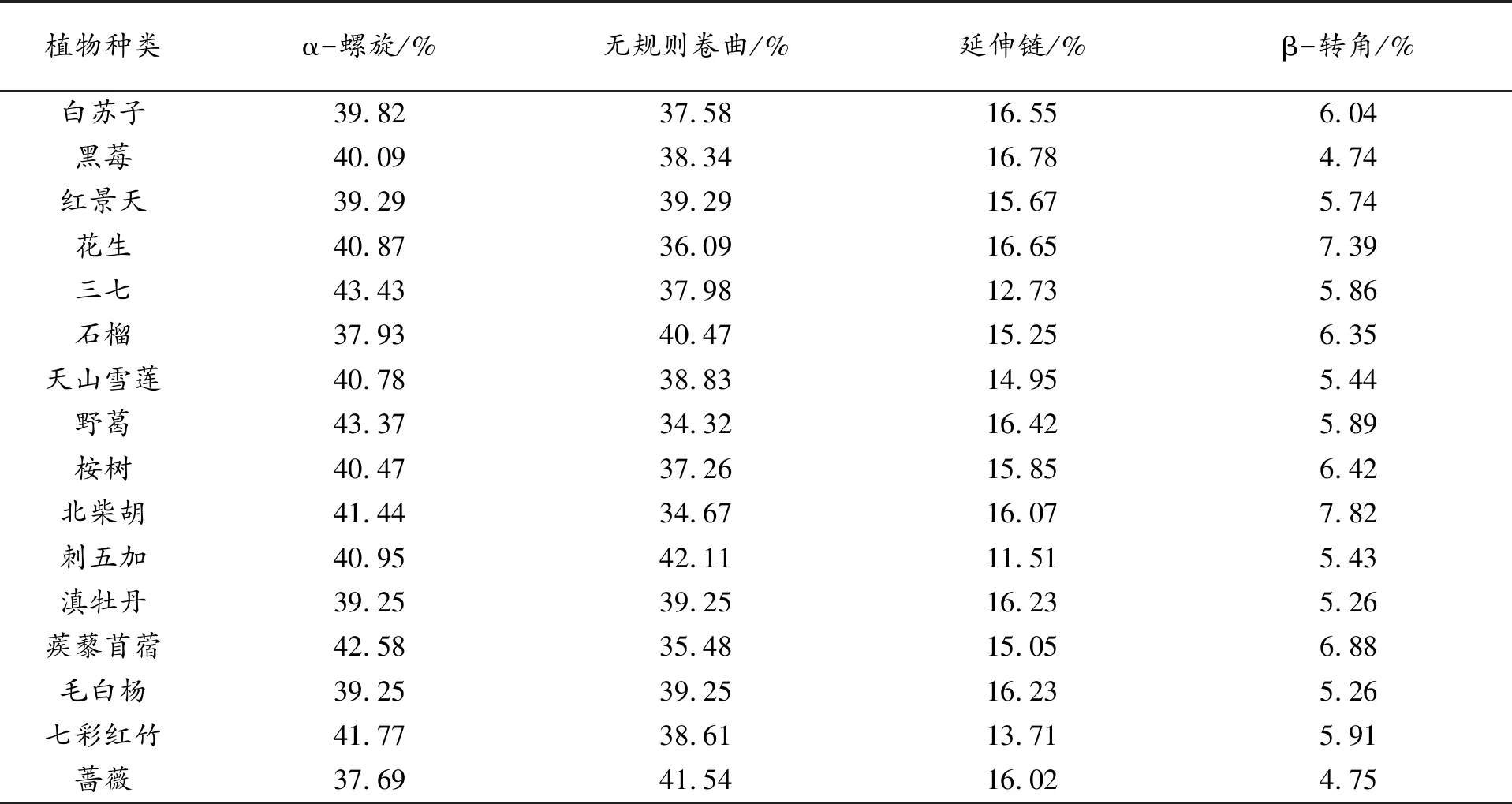
植物種類α-螺旋/%無規則卷曲/%延伸鏈/%β-轉角/%白蘇子39.8237.5816.556.04黑莓40.0938.3416.784.74紅景天39.2939.2915.675.74花生40.8736.0916.657.39三七43.4337.9812.735.86石榴37.9340.4715.256.35天山雪蓮40.7838.8314.955.44野葛43.3734.3216.425.89桉樹40.4737.2615.856.42北柴胡41.4434.6716.077.82刺五加40.9542.1111.515.43滇牡丹39.2539.2516.235.26蒺藜苜蓿42.5835.4815.056.88毛白楊39.2539.2516.235.26七彩紅竹41.7738.6113.715.91薔薇37.6941.5416.024.75
由表3可知,在天山雪蓮的二級結構中α-螺旋出現的概率最高為40.78%,無規則卷曲比例接近于α-螺旋為38.83%,延伸鏈較少為14.95%,β-轉角最少為5.44%。同時可以看出,α-螺旋和無規則卷曲是糖基轉移酶多肽鏈中大量存在的結構,高達79.61%。其余15種植物中的糖基轉移酶二級結構具有類似的比例結構。
3 討論
糖基轉移酶催化的糖基化反應是三萜皂苷生物合成的最后一步,使得單糖與三萜皂苷苷元由糖苷鍵相連[10]。對植物的生長發育過程和代謝工程應用方面起著重要作用,催化植物次生代謝物合成[11]。本文通過使用生物信息學方法,對16種植物的糖基轉移酶蛋白特性進行分析。結果表明,在氨基酸理化性質的分析中顯示植物糖基轉移酶含量最豐富的氨基酸為Leu、Gly、Val、Ser和Ala等,桉樹、北柴胡、滇牡丹、蒺藜苜蓿、毛白楊、七彩紅竹、薔薇和紅景天糖基轉移酶屬于穩定類蛋白,其余則屬于不穩定類蛋白;蛋白質信號肽預測分析中顯示紅景天糖基轉移酶可能含有信號肽,其余植物均不含有信號肽;糖基轉移酶蛋白二級結構均由α-螺旋、無規則卷曲、延伸鏈和β-轉角組成,且比例相似,α-螺旋和無規則卷曲為主要結構,占總的80%左右,延伸鏈次之,為15%左右,β-轉角最少,為5%左右。
綜合三種不同分析工具對16種植物中糖基轉移酶的研究表明,這16種植物的糖基轉移酶蛋白特性差別不大,顯示糖基轉移酶具有較高的保守性和穩定性。同時許多基因組數據表明,糖基轉移酶基因在進化上不是獨立的,而是與其他基因有較強的關聯性;同時代謝途徑中存在基因共調節和蛋白相互作用[12]。糖基轉移酶基因進化主要依賴基因重復,特別是串聯重復,而不是片段重復[13]。對蛋白特性的解析能夠更清楚的揭示酶的保守結構域及作用機制,為進行糖基轉移酶的基因工程改造和分子進化研究提供借鑒,本文章對糖基轉移酶的生物信息學分析,也可為研究糖基轉移酶酶學特征及三萜皂苷代謝提供幫助。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
