常見乳酸菌降解亞硝酸鹽機理探討
2019-05-07 08:00:02王一茜榮金誠王曉輝遲乃玉張慶芳李美玉
食品與發酵工業 2019年8期
王一茜,榮金誠,王曉輝,遲乃玉,張慶芳*,李美玉
1(大連大學 生命科學與技術學院,遼寧 大連,116622)2(遼寧省海洋微生物工程技術研究中心,遼寧 大連,116622)
利用方差分析、相關性分析等數據分析方法,張慶芳等[19]推導出了乳酸菌對亞硝酸鹽的降解分為NiR降解和酸降解 2個階段。在發酵的前期,培養液pH>4.5時,乳酸菌對亞硝酸鹽降解以NiR降解為主;發酵后期,由于乳酸菌本身產生酸,使培養液pH值降低,pH<4.0后,亞硝酸鹽的降解主要以酸降解為主。由于乳酸桿菌產酸能力強于乳酸球菌,乳酸桿菌降解亞硝酸鹽能力大于乳酸球菌,而在發酵前期(pH>4.5),桿菌與球菌降解亞硝酸鹽并無差別。

1 材料與方法
1.1 亞硝酸鹽降解研究
1.1.1 試驗菌種
Lactobacillusbrevis1508;Lactobacillusplantarum1407;Lactobacillusrhamnosus1.11,以上菌株為本研究室保藏。
1.1.2 蔬菜原料
大連市郊區甘藍。
1.1.3 培養基及培養液配方
MRS固體培養基(g/L):葡萄糖20、蛋白胨10、酵母粉4、K2HPO42、牛肉粉8、MgSO40.1、乙酸鈉5、檸檬酸三銨2、MnSO40.05、吐溫-80 1、瓊脂20、pH 7.2。
液體培養基:不加瓊脂,其他成分同MRS固體培養基。
1.1.4 菌種活化
菌種活化培養基為MRS液體培養基。一次活化:取純化后穿刺保藏的各菌種試管,分別穿刺3針接于10 mL液體培養基試管中,重復3次,30 ℃培養48 h。二次活化:取一次活化各菌種菌液,按10%接種量接種于液體培養基擴大培養,30 ℃培養48 h。
1.1.5 儀器與設備
HD-1360超凈工作臺,北京悅泰行科技發展有限公司;THZ-312臺式恒溫振蕩器,上海精勝科學儀器設備有限公司;280型輕便手提壓力蒸汽消毒器,成都一科儀器設備有限公司;BZ-01顯微鏡,德國徠卡;UV-120-02紫外可見分光光度計,上海珂淮儀器有限公司;pHB-4p酸度計,上海升隆公司。
1.1.6 蔬菜發酵工藝流程
甘蘭→清洗→燙漂→瀝水→切段→裝瓶→壓石→注鹽水→接種→密封→發酵
1.1.7 測定指標及方法

1.2 參考序列
菌株的16S rDNA序列下載自NCBI核酸數據庫,包括L.bifermentans[M58809.1],L.brevis[NR_044704.1],L.composti[NR_041509.1],L.coryniformis[NR_044705.2],L.plantarum[D79210.1],L.rhamnosus[M58815.1],L.similis[KJ547681.1]。
乳桿菌的亞硝酸鹽還原酶蛋白序列下載自NCBI蛋白數據庫,包括L.bifermentans[WP_057904912.1],L.brevis[WP_035444188.1],L.composti[WP_057003017.1],L.coryniformis[EJN56181.1],L.plantarum[AKC01517.1],L.similis[WP_057151639.1]。此外,經詳細搜索比對發現,菌株L.rhamnosus的亞硝酸鹽還原酶基因在網上數據庫中無序列信息。
1.3 數據庫和分析軟件
GenBank數據庫:核酸序列數據庫,包含基因核酸與蛋白序列、非編碼序列、基因組序列等,由美國國立生物技術信息中心(NCBI)建立并維護;BLAST(序列局部比對工具,有blastp、blastn、blastx、tblastn、tblastx五種比對方法);Clustal X2(多序列比對工具),MEGA5(系統發育分析軟件),BioEdit(序列比對與分析工具),蛋白保守結構域由NCBI的CDD預測。
2 結果與分析
2.1 乳酸菌發酵產酸特性
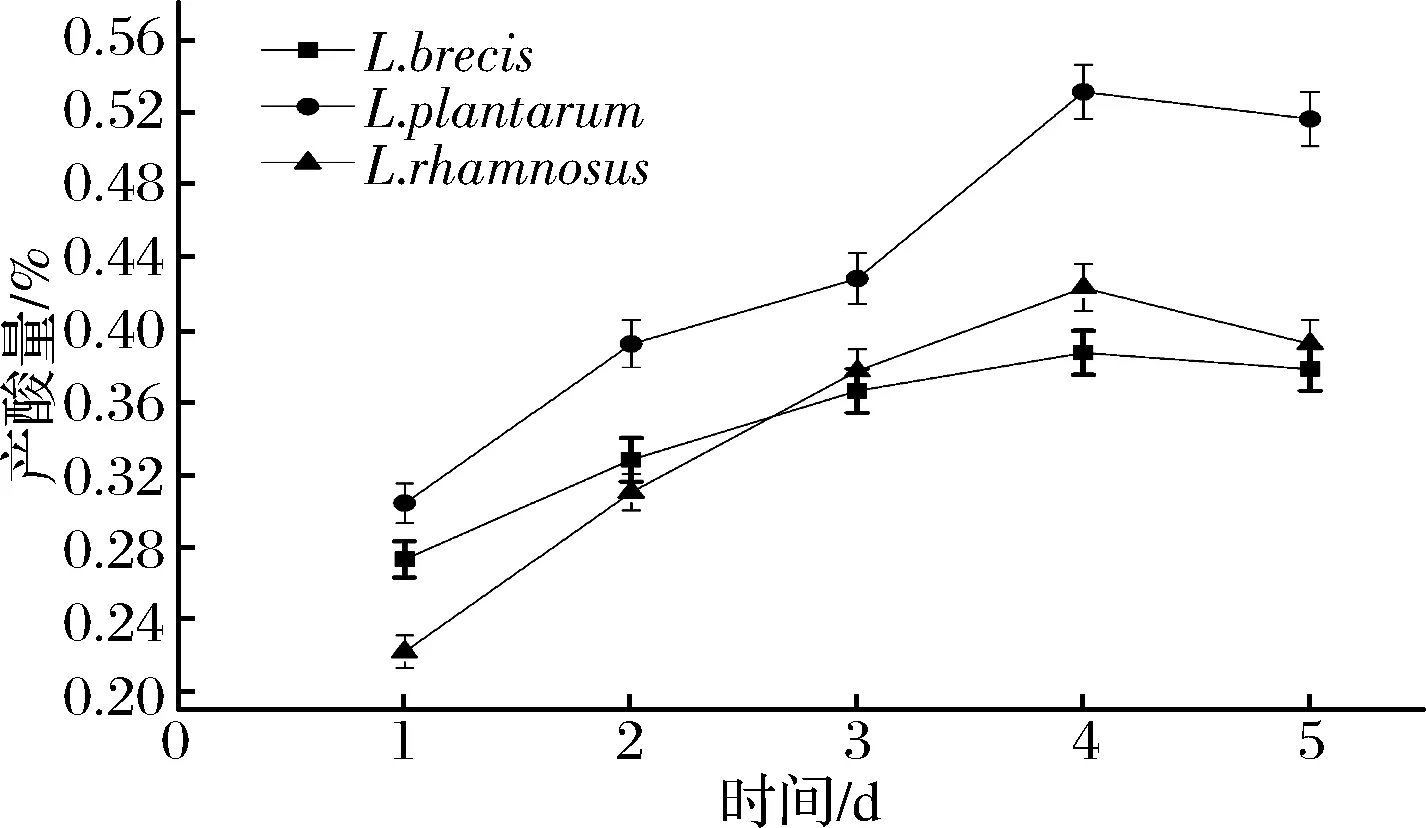

圖1 無培養液中總酸的變化Fig.1 The change of culture medium (without total acid produced
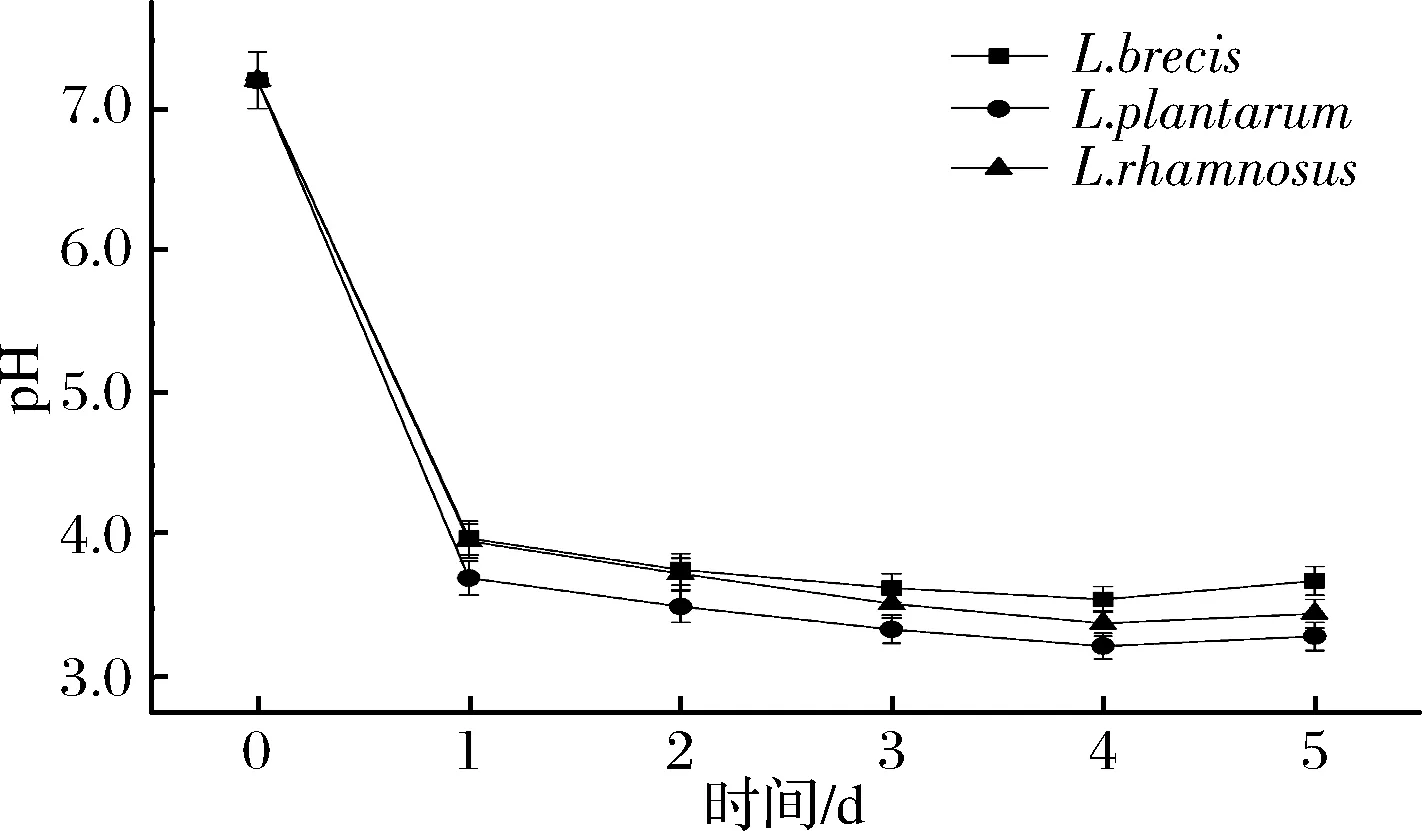

圖2 無培養液中pH值的變化Fig.2 The change of pH in culture medium broth without
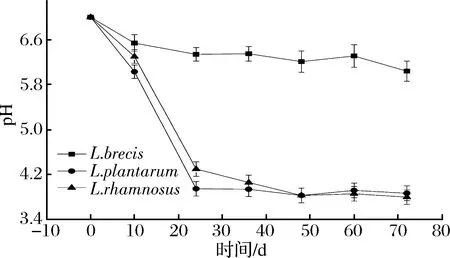
圖3 有培養液中pH值的變化Fig.3 The change of pH in culture medium broth with
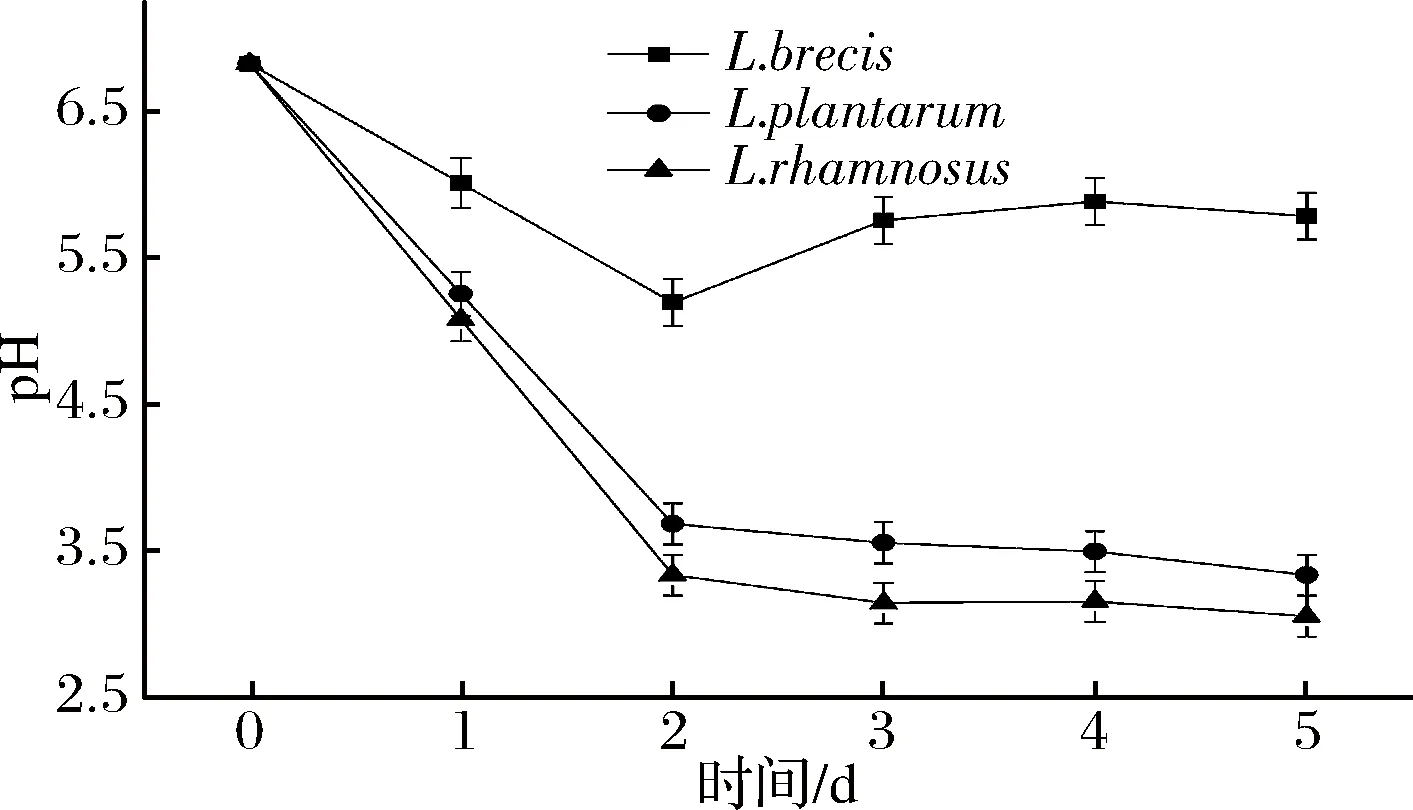
圖4 蔬菜發酵液的pH變化Fig.4 The change of pH in vegetable fermented solution
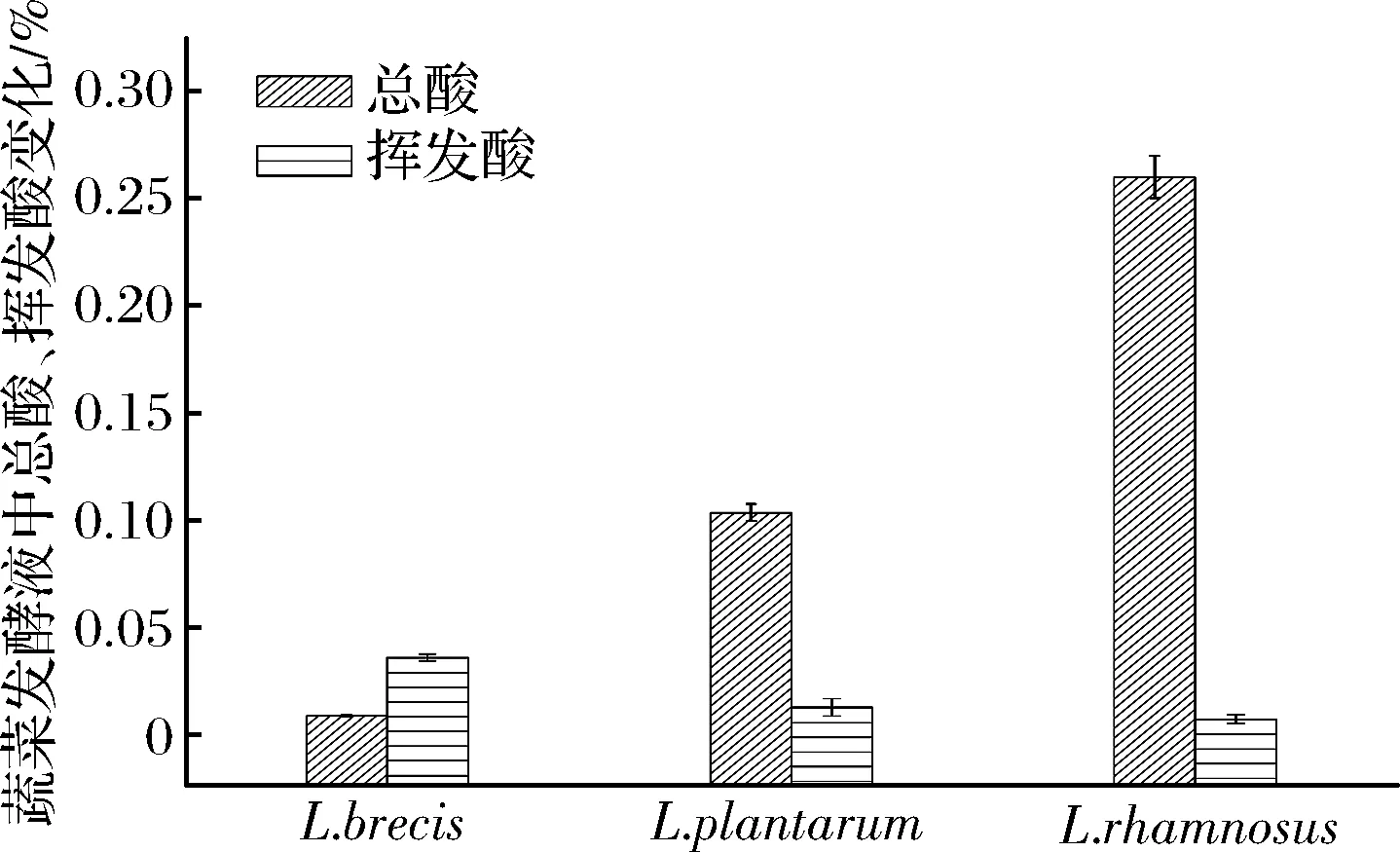
圖5 蔬菜發酵液的總酸、揮發酸含量Fig.5 The change of total acid and volatile acid content of vegetable fermented solution
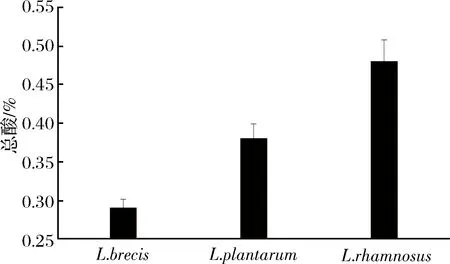
圖6 發酵菜的總酸含量Fig.6 The change of total acid of vegetable fermented solution
2.2 乳酸菌基因分析
2.2.1 亞硝酸鹽還原酶基因的深入分析
基于NCBI數據庫,本研究對L.rhamnosus、L.plantarum、L.brevis所有可獲得的基因組進行了分析,通過基因組功能注釋查找nir基因,并利用BLASTP比對參考序列(NiR蛋白序列),發現L.brevis與L.plantarum菌株中存在nir基因,而L.rhamnosus菌株中未發現該基因。
進一步的多序列比對結果可看出(圖7),除L.plantarum外,其余5株乳桿菌的亞硝酸鹽還原酶的蛋白序列的相似性很高(相似性在80%以上),只在蛋白序列的前端與末端存在較高比例的差異氨基酸。蛋白質的保守結構域分析(CDD分析)也表明,乳桿菌(除L.plantarum外)的亞硝酸鹽還原酶均包含1個NiR_SIR_ferr超家族結構域,該結構域為亞硝酸鹽/亞硫酸鹽還原酶的保守結構域家族。而且,該結構域的氨基酸序列(占全酶序列長的86.9%)在6株乳桿菌中是高度保守的,與多序列比對的同源性較高的區域相一致,這表明在歷史進化中NiR_SIR_ferr超家族結構域的穩定遺傳決定了亞硝酸鹽還原酶的穩定存在與功能實現。

圖7 L. plantarum、L. brevis與其他乳桿菌的亞硝酸鹽還原酶蛋白序列的多序列比對與結構域分析Fig.7 The multiple sequence alignment of the nitrite reductase protein sequence and the domain analysis of L. plantarum, L. brevis and other Lactobacillus
2.2.2 系統發育分析
如圖8-A所示,基于6株乳桿菌(包括L.rhamnosus)的16S rDNA序列構建的系統發育樹分化為2個明顯的大分枝,L.rhamnosus與L.bifermentans、L.coryniformis與L.composti在同一大分枝上,L.plantarum、L.brevis與L.similis在同一大分枝上,而同一大分枝上的菌株的親緣關系更近,也即L.plantarum與L.brevis親緣關系更近。
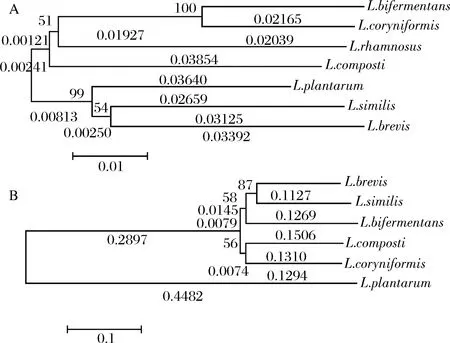
A-16S rDNA序列; B-系統發育樹圖8 基于L. plantarum、L. brevis、L. rhamnosus與其他乳桿菌的16S rDNA序列與亞硝酸鹽還原酶蛋白序列構建的系統發育樹Fig.8 The phylogenetic tree built based on Lactobacillus plantarum, Lactobacillus brevis, Lactobacillus rhamnosus and other Lactobacillus 16S rDNA sequences and the nitrite reductase protein sequence注:系統發育樹由MEGA5構建,采用Neighbor-Joining方法,重復500次建樹,進化距離由p-distance方法運算獲得。
然而,基于亞硝酸鹽還原酶蛋白序列構建的系統發育樹上(圖8-B,不包括L.rhamnosus),L.plantarum所在的進化分枝獨立于其他菌株,這與16S rDNA序列構建的系統發育樹明顯不同。而且,由圖8-B可知,L.plantarum的亞硝酸鹽還原酶基因在歷史進化中最先分化,且在長期進化過程中形成了獨特的亞硝酸鹽還原酶基因結構。
2.3 乳酸菌發酵降解亞硝酸鹽的能力
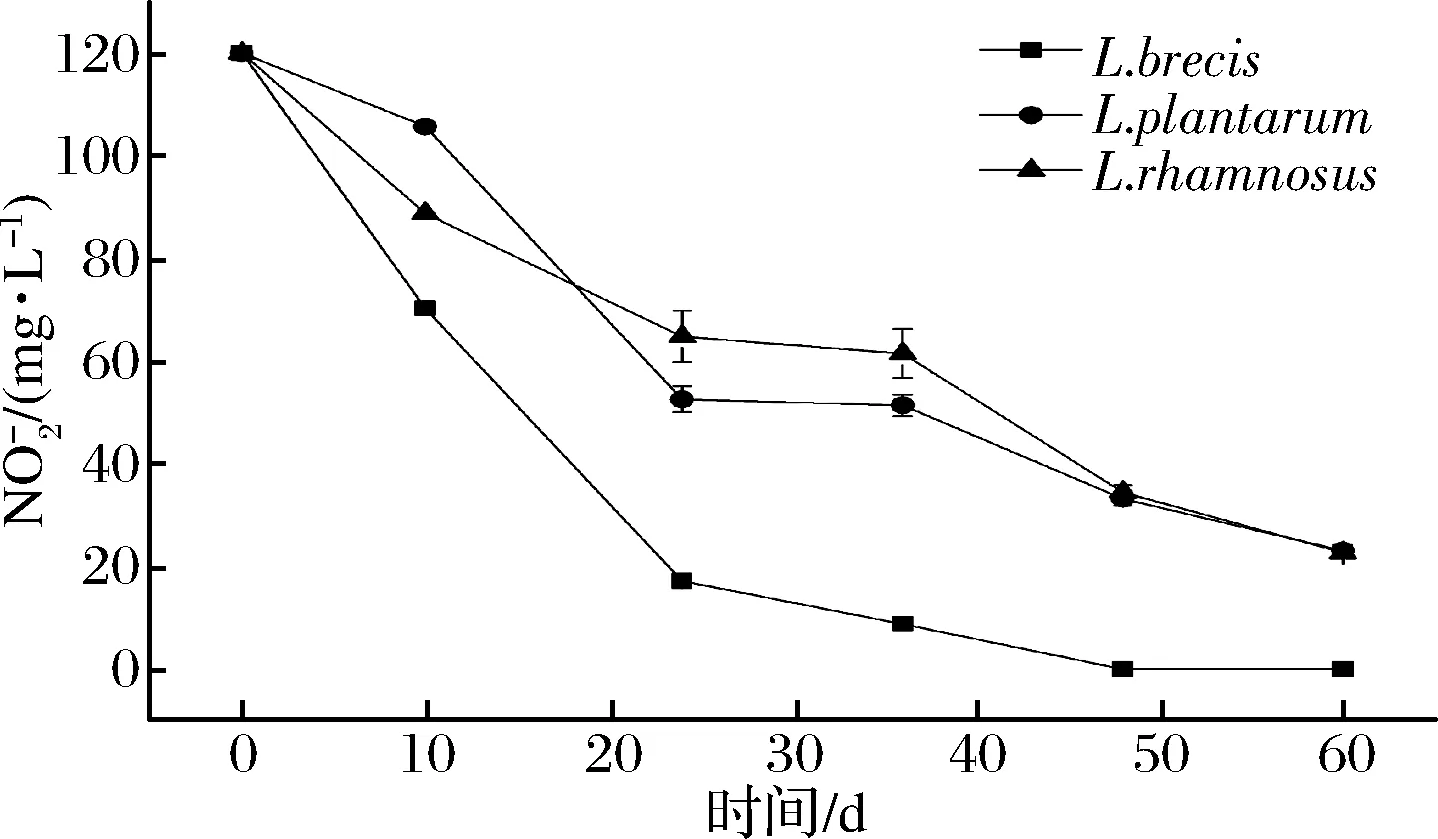
圖9 培養液含量變化Fig.9 The change of content of in culture solution
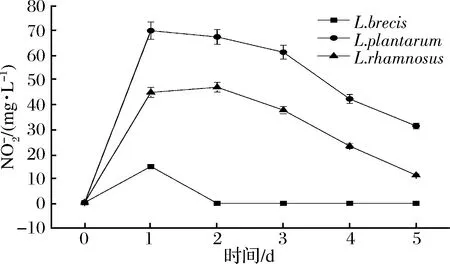
圖10 蔬菜發酵液中含量變化Fig.10 The change of content of in vegetable fermented solution
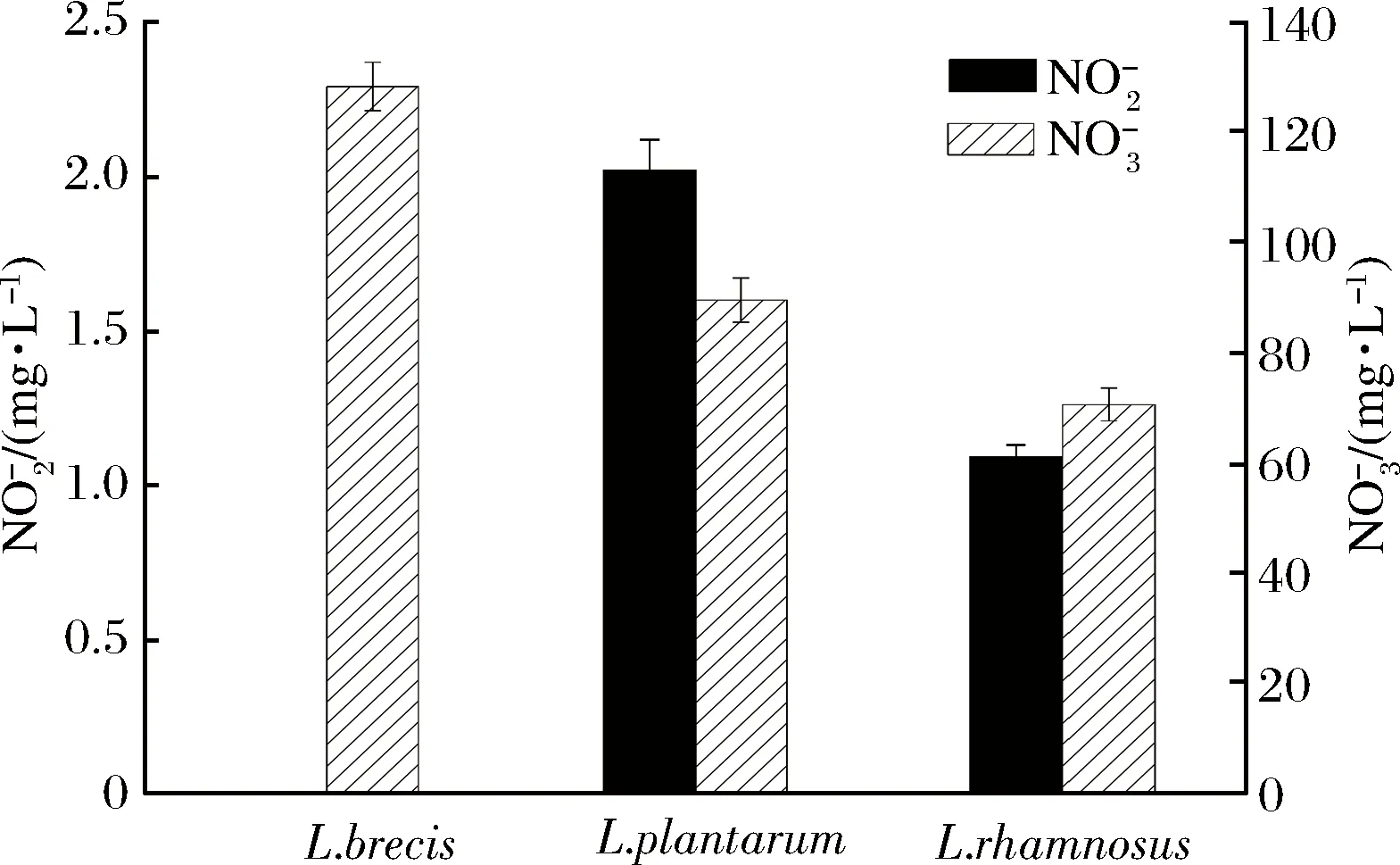
圖11 發酵菜中殘留量Fig.11 Residual of fermented vegetable
3 討論
張慶芳等[19]推導出了乳酸菌對亞硝酸鹽的降解分為NiR降解和酸降解 2個階段。在發酵的前期,培養液pH>4.5時,乳酸菌對亞硝酸鹽降解以NiR降解為主;發酵后期,由于乳酸菌本身產生酸,使培養液pH值降低,pH<4.0后,亞硝酸鹽的降解主要以酸降解為主。
本實驗室及其他學者[33-37]均發現L.rhamnosus有降解亞硝酸鹽的能力。但經詳細搜索基因庫發現,菌株L.rhamnosus的亞硝酸鹽還原酶基因在網上數據庫中無序列信息。可能原因如下:(1)L.rhamnosus有nir基因,但國內外無相關研究。見圖9,在亞硝酸鹽純培養液發酵前期(非酸降解階段)有大量亞硝酸鹽被降解;由圖10和圖11可知,蔬菜原料中有大量硝酸鹽被分解使得發酵環境中有大量亞硝酸鹽生成,隨后亞硝酸鹽被大量降解。(2)L.rhamnosus沒有nir基因。如圖9亞硝酸鹽在前期(pH>4.5)又有降解,可能是此菌產生的乳酸菌素[38]等其他物質作用(如乳酸菌素最適作用pH為2~8,涵蓋了NiR降解階段);但此菌降解亞硝酸鹽能力較強(見圖10),L.rhamnosus是正型發酵,見發酵產酸特性3.1的分析,其產酸能力較強,且降解亞硝酸鹽可能是氮代謝途徑[39-42](如果有亞硝酸鹽還原酶),不能使其發酵環境的pH值回調,使其快速降到4.0以下,進入酸降解階段。
