茯苓菌固態(tài)發(fā)酵小麥基質(zhì)的成分變化
2019-04-24 07:45:38李佳歡金文松張燎原胡開輝
食品與生物技術學報 2019年12期
關鍵詞:質(zhì)量
李佳歡 , 林 輝 , 金文松 , 張燎原 , 胡開輝 *
(1.福建農(nóng)林大學 生命科學學院,福建 福州 350002;2.福建農(nóng)林大學(古田)菌業(yè)研究院,福建 寧德352200)
茯苓(Poria cocos(Schw.)Wolf)作為一種藥用真菌,目前已報道的生物活性物質(zhì)包括多糖類、甾體類、蛋白質(zhì)、四環(huán)三萜類、三環(huán)二萜類等[1-3]。現(xiàn)代藥學研究證實,茯苓具有抗腫瘤、抗氧化、增強免疫功能等多種生物活性[4-6]。現(xiàn)階段,人們對茯苓的利用不僅局限于子實體,還可以通過其菌絲體進行液態(tài)或固態(tài)發(fā)酵,提取其發(fā)酵產(chǎn)物。
谷物因具有豐富的營養(yǎng)成分,如淀粉、蛋白質(zhì)、植物脂肪、微量元素等,可供給微生物生長所需的營養(yǎng)成分,是很好的發(fā)酵基質(zhì)。營養(yǎng)學分析表明,谷物在經(jīng)發(fā)酵后,其營養(yǎng)成分將會更加豐富。文獻[7]的研究表明,經(jīng)自然發(fā)酵后的豇豆等豆科植物,其賴氨酸、蘇氨酸等必需氨基酸質(zhì)量分數(shù)明顯提升。文獻[8]的研究中指出,燕麥、大豆2種混合谷物經(jīng)灰樹花孔菌進行固態(tài)發(fā)酵后,其總?cè)啤⒖偠喾拥瓤寡趸钚猿煞志兴嵘宄杂苫芰︼@著增強。文獻[9]對桑黃發(fā)酵的薏米、稻米展開研究,發(fā)現(xiàn)兩種谷物經(jīng)發(fā)酵后,其抗氧化能力明顯增強。
在微生物的作用下,谷物的營養(yǎng)組成發(fā)生了變化,同時發(fā)酵中產(chǎn)生的代謝產(chǎn)物,一定程度上提升了谷物的附加值。但在已有的研究中鮮見以茯苓菌為菌種對谷物進行發(fā)酵的報道,更未有研究對發(fā)酵過程中的酶活及各有效物質(zhì)的變化展開研究。本研究采用無毒保健的茯苓菌對小麥進行發(fā)酵,初步分析了發(fā)酵過程中茯苓菌關鍵酶活性質(zhì)的變化,并探討了酶活變化與產(chǎn)物營養(yǎng)成分變化之間的關系,以期為茯苓發(fā)酵口服液、飲料等功能性產(chǎn)品的開發(fā)提供借鑒。
1 材料與方法
1.1 原料與試劑
小麥購自福州農(nóng)貿(mào)市場;茯苓菌種由福建農(nóng)林大學微生物工程實驗室提供;葡萄糖購自Sigma試劑公司;其他試劑皆為分析純試劑。
1.2 儀器與設備
Agilent1100高效液相色譜儀,美國 Agilent公司產(chǎn)品;BS110S型電子天平,賽多利斯科學儀器(北京)有限公司產(chǎn)品;LDZX-50KBS型立式壓力蒸汽滅菌鍋,上海申安醫(yī)療器械廠制造;DHG-9203型鼓風干燥機,上海精宏公司產(chǎn)品;LRH-250型恒溫培養(yǎng)箱,寧波江南儀器廠制造;UV1100型紫外分光光度計,上海美譜達儀器有限公司產(chǎn)品;5804R型高速冷凍離心機,德國Eppendorf公司產(chǎn)品;PB10型pH計,德國賽多利斯集團產(chǎn)品。
1.3 實驗方法
1.3.1 培養(yǎng)基 PDA加富固體培養(yǎng)基:土豆(去皮)200 g、葡萄糖 20 g、蛋白胨3 g、酵母提取粉 3 g、硫酸鎂 1.5 g、磷酸二氫鉀 1.5 g、維生素 B10.1 g、瓊脂 20 g,加水定容至1 000 mL,pH自然,115℃滅菌30 min。
小麥固體培養(yǎng)基:去除小麥中雜質(zhì)與空心小麥顆粒,用蒸餾水沖洗2~3次,去除灰塵、麥麩等雜質(zhì),加蒸餾水至剛好浸沒小麥,于40℃水浴2 h,將小麥撈出,按料液比1∶1.2加入蒸餾水煮至小麥飽滿全熟無白心,注意小麥不能裂開,以避免營養(yǎng)散失。將煮好的小麥粒分裝于栽培瓶中,每瓶110 g,壓平,封口膜封口后121℃滅菌120 min。
1.3.2 發(fā)酵與樣品收集 將滅菌后的小麥固體培養(yǎng)基冷卻至室溫,取活化好的茯苓菌種按5 mL/hg的接種量接至小麥栽培料,置于26℃恒溫培養(yǎng)箱中培養(yǎng)。接種后每隔5 d取1次樣,將所得樣品混勻后置于60℃鼓風干燥機中干燥48 h。將干燥后的發(fā)酵小麥經(jīng)中草藥粉碎機粉碎后,過60目篩,于干燥封閉環(huán)境下保存以供后續(xù)營養(yǎng)成分測定實驗。未接種的小麥經(jīng)過上述處理后作為對照,所有實驗設定3組平行。
1.3.3 提取液的制備 將發(fā)酵小麥取出攪拌均勻后,稱取10 g于研缽中,研磨后移至100 mL三角瓶中,加入50 mL雙蒸水,于150 r/min,25℃恒溫振蕩搖床培養(yǎng)90 min,經(jīng)紗布過濾后,8 000 r/min離心15 min,取上清液即為提取液,用于相關酶活、草酸和pH值的測定。
1.4 測定方法
1.4.1 酶活力的測定方法 淀粉酶活力測定在文獻[10]的方法上進行改良,采用淀粉-碘化鉀顯色法。定義60℃、pH 6.0條件下,1 min液化1 μg可溶性淀粉所需酶量為1個酶活力單位“U”。
木聚糖酶活力測定方法在文獻[11]等方法上進行改良,采用3,5-二硝基水楊酸(DNS)法。定義50℃、pH=5.2條件下,1 min生成 1 μg D-木糖所需酶量為1個酶活力單位“U”。
羧甲基纖維素酶的測定方法在文獻[12]的方法上進行改良,采用3,5-二硝基水楊酸(DNS)法。定義50 ℃、pH 4.6的條件下,1 min生成 1 μg葡萄糖(GLU)所需酶量為1個酶活力單位“U”。
1.4.2 茯苓菌發(fā)酵小麥營養(yǎng)成分變化
1)pH值測定:用pH計對樣品粗酶提取液進行測定。
2)草酸測定方法:本研究采用HPLC法來測定小麥中草酸的質(zhì)量分數(shù)。取粗酶液0.4 mL,加入0.2 mL酸性乙醇,在振蕩器上混合1 min,8 000 r/min離心10 min,上清液經(jīng)0.22 μm濾膜過濾。色譜分離柱:SB-Aq C18柱;流動相:0.025 mol/L磷酸二氫鉀和四丁基氫氧化銨溶液(磷酸調(diào)pH2.0);紫外檢測波長:220 nm;進樣量 2 μL;流動相速度:流速 0.6 mL/min;柱溫30℃。以草酸標品質(zhì)量濃度(mg/mL)為X軸與峰面積的值(Y軸)繪制標準曲線。所得標曲的線性回歸方程為Y=3 039.1X+5.096,R2=0.997 7。
3)總糖測定:參考文獻[13]的方法測定小麥當中的總糖質(zhì)量分數(shù),采用葡萄糖溶液進行標準曲線的測定,根據(jù)葡萄糖質(zhì)量(X軸)與OD值(Y軸)繪制標準曲線。所得標準曲線的線性回歸方程為:

4)還原糖質(zhì)量分數(shù)測定:參考文獻[14-15]的方法測定小麥當中的還原糖質(zhì)量分數(shù),采用DNS法進行還原糖測定。以葡萄糖溶液進行標準曲線的測定,根據(jù)葡萄糖質(zhì)量(X軸)與OD值(Y軸)繪制標準曲線。所得標準曲線的線性回歸方程為:
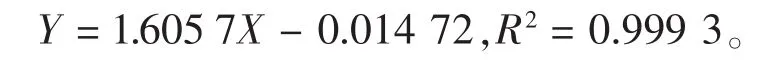
5)多糖測定:參考文獻[16]的方法提取小麥粉中多糖,用苯酚硫酸法測定其吸光值,將結果代入葡萄糖標準曲線計算其多糖質(zhì)量分數(shù)。
6)總?cè)瀑|(zhì)量分數(shù)測定:參考文獻[17]的方法對小麥中的總?cè)疲捎孟悴萑?冰醋酸測定樣品中三萜質(zhì)量分數(shù)。以熊果酸作為標品進行標準曲線的測定,根據(jù)熊果酸質(zhì)量(X軸)與OD值(Y軸)繪制標準曲線,所得標準曲線的線性回歸方程為:
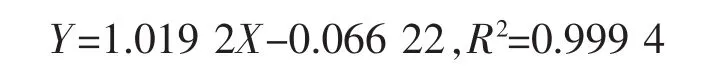
7)水溶性三萜酸測定:取10 g栽培料至三角瓶,加入50 mL的雙蒸水,60℃水浴2 h,每隔半小時搖晃混勻,8 000 r/min離心15 min,取上清液2 mL于烘干箱60℃烘干,烘干后用1 mL雙蒸水溶解,添加 1 mL乙酸乙酯(質(zhì)量分數(shù)99.5%)進行萃取2遍;合并上層液烘干后使用香草醛-冰醋酸測定樣品中三萜酸質(zhì)量分數(shù)。
1.5 數(shù)據(jù)處理
實驗數(shù)據(jù)用SPSS 20.0軟件進行統(tǒng)計學分析,用EXCEL軟件進行繪圖。
2 結果與分析
2.1 茯苓發(fā)酵小麥過程中菌絲生長變化情況
圖1表示的是不同培養(yǎng)時間下,茯苓菌在小麥培養(yǎng)基上的長勢情況,可以看出茯苓菌絲呈白色,菌絲較為纖細,生長速度快,培養(yǎng)6 d即可滿瓶。隨著培養(yǎng)時間的延長,菌絲密度逐漸升高且呈白色,充斥麥粒之間的間隙,培養(yǎng)至30 d后菌絲密度不再發(fā)生明顯變化。

圖1 茯苓栽培過程的生長情況Fig.1 Growth of the P.cocos
2.2 茯苓固態(tài)發(fā)酵過程中酶活力變化情況
茯苓栽培過程中纖維素酶、木聚糖酶和淀粉酶的活力變化如圖2所示。
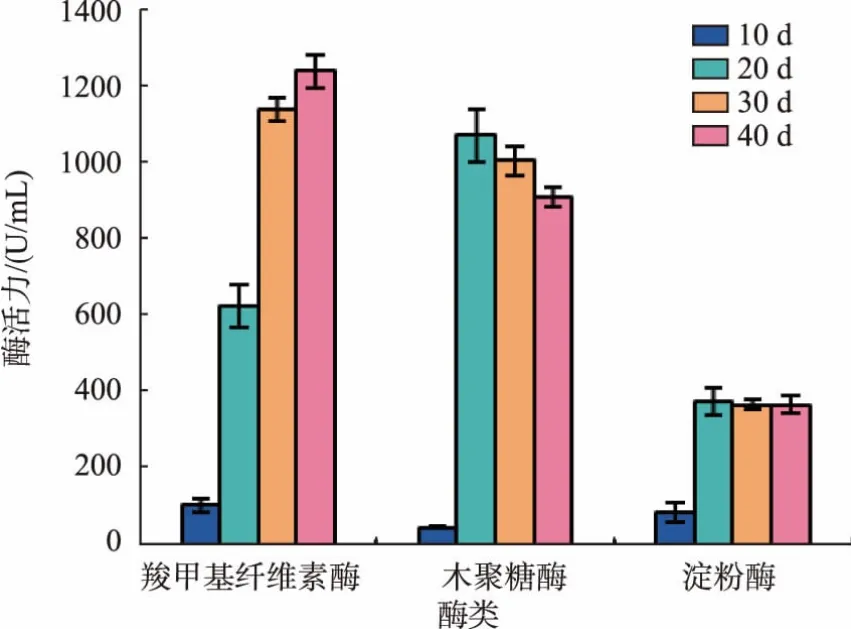
圖2 茯苓固態(tài)發(fā)酵過程纖維素酶、木聚糖酶和淀粉酶活力Fig.2 Activity variation of cellulose,xylanase and amylase produced by P.cocos during the process of solid-state fermentation
小麥基質(zhì)中含有大量的淀粉、纖維素、木聚糖等,這些物質(zhì)可以在相關酶的作用下,分解成單糖以供食用菌的生長所需。通過分析發(fā)酵過程中相關酶活力的變化,觀察茯苓菌的生長情況及其對小麥培養(yǎng)基中營養(yǎng)成分的利用情況。
淀粉為小麥中主要貯藏物質(zhì),質(zhì)量分數(shù)約為53%~70%。在茯苓菌發(fā)酵小麥的過程中,淀粉酶的活力對菌絲生長起著重要作用。由圖2可知,隨著發(fā)酵時間的增加,淀粉酶活力不斷增強,在20 d時活力達最大值247.1 U/mL,隨后基本維持在240 U/mL左右,持續(xù)降解淀粉以供茯苓菌絲生長所需。其可能原因為小麥中的淀粉質(zhì)量分數(shù)一直較高,故淀粉酶作為茯苓菌分泌的一種誘導酶,其活力將保持著相對穩(wěn)定的狀態(tài)。
纖維素是麥麩中的主要成分之一,其質(zhì)量分數(shù)約占其干物質(zhì)的35%~40%[18]。茯苓菌若要利用小麥胚乳中的營養(yǎng)成分,首先要具有降解麩皮中纖維素的能力,因此茯苓菌纖維素酶是茯苓菌能否在小麥基質(zhì)中生長的關鍵酶之一。從圖2可以看出,酶的活力在培養(yǎng)10 d時為100 U/mL,隨茯苓菌發(fā)酵時間的延長,纖維素酶活力以每隔10 d 500 U/mL的速度提升,培養(yǎng)30 d時纖維素酶活達1 150 U/mL,后酶活提升趨勢減緩,40 d時其活力為1 250 U/mL左右。說明茯苓菌可以很好的降解小麥胚乳中的纖維素,以供菌絲生長和代謝所需。
小麥中可溶性多糖(非淀粉)的質(zhì)量分數(shù)可達6%以上,而其主要成分為阿拉伯木聚糖,是機體無法消化利用的成分之一,因此將降低其能量的利用效率[19],而茯苓菌分泌的木聚糖酶可降解小麥中的木聚糖。由圖2可知,木聚糖酶活力在10~20 d時顯著提高,由54 U/mL提升至1 080 U/mL,之后隨著培養(yǎng)時間的延長,木聚糖酶活力逐漸降低,在40 d時酶活力為920 U/mL。
2.3 發(fā)酵產(chǎn)物總糖、還原糖及多糖的變化
由圖3可知,小麥發(fā)酵前總糖質(zhì)量分數(shù)高達80%以上,但經(jīng)茯苓菌發(fā)酵后,小麥中的總糖質(zhì)量分數(shù)逐漸降低。培養(yǎng)前10 d總糖降低速度較慢,但10~20 d期間,總糖被迅速消耗,20 d時栽培料中的總糖質(zhì)量分數(shù)僅剩57.2%,隨后總糖下降速率減緩。通過相關性分析發(fā)現(xiàn),總糖的變化與纖維素酶、木聚糖酶及淀粉酶的變化趨勢呈現(xiàn)顯著的負相關性(r值分別為-0.961、-0.925和-0.958), 結合發(fā)酵過程中的酶活力變化可知,培養(yǎng)前10 d,由于茯苓菌菌絲酶活力較弱,對營養(yǎng)的利用率較低,總糖降低速率較低。發(fā)酵10~20 d期間,茯苓菌的代謝較旺盛,生長速度加快,淀粉酶、羧甲基纖維素酶及木聚糖酶的活力在該階段的活力大幅提升(P<0.01)。在3種酶的作用下,小麥中的淀粉、纖維素及木聚糖被大量消耗,總糖質(zhì)量分數(shù)下降速度加快,第20天時產(chǎn)物中的總糖質(zhì)量分數(shù)下降了43.6%。

圖3 茯苓固態(tài)發(fā)酵過程總糖、還原糖、多糖質(zhì)量分數(shù)變化Fig.3 Changesoftotalsugar、reducing sugar and polysaccharidecontents during the solid-state fermentation of P.cocos
茯苓菌在發(fā)酵小麥的過程中,不能直接利用基質(zhì)中的淀粉、纖維素等物質(zhì),而需要通過相應酶系的作用,將其降解為還原糖后再進行利用,所以還原糖質(zhì)量分數(shù)的變化可以在一定程度上反映真菌對碳水化合物的有效利用率[20]。未發(fā)酵小麥的還原糖質(zhì)量分數(shù)僅為0.389%,發(fā)酵過程中小麥還原糖質(zhì)量分數(shù)呈整體上升趨勢,發(fā)酵35 d時的還原糖質(zhì)量分數(shù)高達27.40%,為未發(fā)酵小麥的70倍。通過相關性分析發(fā)現(xiàn),總糖的變化與纖維素酶、木聚糖酶及淀粉酶的變化趨勢呈現(xiàn)正相關性 (r值分別為0.986、0.734 和 0.804)。 發(fā)酵 0~35 d 期間,發(fā)酵產(chǎn)物中的還原糖質(zhì)量分數(shù)不斷上升,可能是由于在發(fā)酵過程中產(chǎn)生的淀粉酶、纖維素酶、木聚糖酶等多酶體系的作用下,碳水化合物轉(zhuǎn)化為還原糖以供菌絲生長利用,而在35 d之后,還原糖質(zhì)量分數(shù)明顯下降,提示茯苓菌菌絲對淀粉、纖維素等碳水化合物的利用率降低。
茯苓發(fā)酵小麥多糖的質(zhì)量分數(shù)在培養(yǎng)0~20 d內(nèi)大幅上升,由8.72%上升至26.83%。多糖質(zhì)量分數(shù)在培養(yǎng)20 d后開始下降,至40 d時下降為14.96%。可能是因為發(fā)酵前期,在相關酶系的作用下,小麥基質(zhì)中的碳水化合物被降解為單糖、還原糖和多糖,茯苓菌首先利用單糖和還原糖,多糖積累下來,其質(zhì)量分數(shù)逐漸提升。隨著茯苓菌代謝逐漸旺盛,其他糖類不能供給茯苓菌絲的正常生長,茯苓菌開始降解小麥基質(zhì)中的多糖供能。多糖的變化情況其與淀粉酶之間存在著一定的正相關性(r=-0.468)。
2.4 發(fā)酵產(chǎn)物pH值及草酸的變化情況
由圖4可知,茯苓菌發(fā)酵小麥的pH值隨培養(yǎng)時間增加而呈下降趨勢,后期趨于平緩。發(fā)酵前期,茯苓菌生長進入對數(shù)期,由于其生長速度快,在利用小麥淀粉為碳源生長時,產(chǎn)生了大量草酸、月桂酸、茯苓酸等有機酸,pH顯著下降,在15 d左右pH值降至3.70左右后逐漸趨于平穩(wěn),進入穩(wěn)定期,此時茯苓菌生物量基本保持穩(wěn)定,有利于產(chǎn)物的合成。而隨著培養(yǎng)時間的延長,草酸質(zhì)量分數(shù)有所提高,但在培養(yǎng)的25 d內(nèi)草酸質(zhì)量分數(shù)變化不大,質(zhì)量分數(shù)維持在1.2 g/kg左右,而培養(yǎng)25 d以后,產(chǎn)物中的草酸質(zhì)量分數(shù)迅速提高。

圖4 茯苓固態(tài)發(fā)酵過程pH值及草酸質(zhì)量分數(shù)變化Fig.4 Changes of pH and oxalic acid during the process of solid-state fermentation of P.cocos
2.5 發(fā)酵產(chǎn)物三萜質(zhì)量分數(shù)的變化
三萜是萜類物質(zhì)中的一種,是真菌和植物在生長過程中產(chǎn)生的一種次級代謝產(chǎn)物。具有抗腫瘤、免疫調(diào)節(jié)、醛固酮拮抗等功效,是一種極具潛力的功能性物質(zhì)[21]。然而僅通過日常飲食攝入,無法滿足人體對三萜的需求。因此,通過食用菌對谷物發(fā)酵,既改良了谷物原本的營養(yǎng)結構,同時可制成富含三萜類物質(zhì)的功能性食品。由圖5可知,發(fā)酵過程中總?cè)婆c水溶性三萜酸變化趨勢相同,隨著培養(yǎng)時間的延長,總?cè)婆c水溶性三萜酸質(zhì)量分數(shù)逐漸提高,在培養(yǎng)至25 d時達到最高值,分別為1.6 g/kg與0.13 g/kg,但隨著培養(yǎng)時間的繼續(xù)延長,兩者的質(zhì)量分數(shù)略有降低。研究指出,茯苓子實體、菌絲體三萜類化合物種類并無差異,只是其質(zhì)量分數(shù)有所不同。本研究得到的茯苓發(fā)酵小麥,采用全麥小麥培養(yǎng)基,通過茯苓菌進行發(fā)酵,其菌絲的生長極大地提升了小麥中的三萜物質(zhì)質(zhì)量分數(shù),因此,可以以茯苓發(fā)酵后小麥為原料,開發(fā)富含三萜類物質(zhì)的功能性食品。
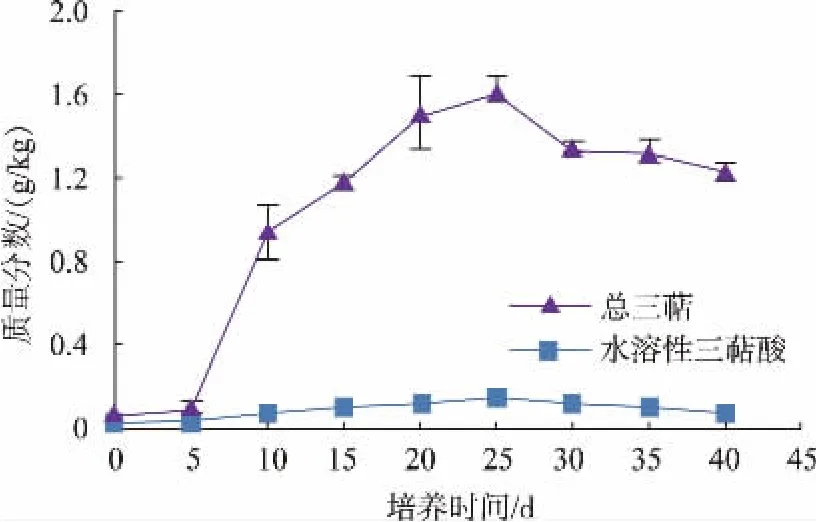
圖5 茯苓固態(tài)發(fā)酵過程三萜質(zhì)量分數(shù)變化Fig.5 Changes oftriterpenes contents during the process of the solid-state fermentation of P.cocos
2.6 討論
茯苓菌作為一種木腐菌,在生長過程中可以分泌多種酶,將大分子營養(yǎng)物質(zhì)降解為可供其利用的小分子物質(zhì)。研究菌絲生長過程中的關鍵酶的活性,可以反映出菌絲的生長發(fā)育情況及其對基質(zhì)的生物轉(zhuǎn)化率[22]。本文對茯苓菌發(fā)酵小麥的過程中的酶活分析結果表明,纖維素酶活力隨培養(yǎng)時間延長不斷上升,40 d時達1 280.5 U/mL,這與文獻[23]在研究茯苓菌發(fā)酵木屑培養(yǎng)基中纖維素酶的變化趨勢不同,其原因可能是所采用的發(fā)酵基質(zhì)不同;文獻[24]對杏鮑菇固態(tài)發(fā)酵過程中木聚糖酶的變化情況的研究與本研究所得結果一致,在茯苓菌發(fā)酵小麥的過程中,木聚糖酶活力在20 d時達最大值,為1 073.2 U/mL,隨后略有下降,可能原因是發(fā)酵后期,小麥中木聚糖質(zhì)量分數(shù)下降,導致其酶活力逐漸降低;淀粉酶活力在發(fā)酵20 d時達最高值,約為494.2 U/mL,隨后基本保持穩(wěn)定,這一結果與文獻[25]對銀耳與香灰菌混合培養(yǎng)過程中的變化趨勢一致。由于小麥胚乳中含有大量淀粉,淀粉酶可能作為一種誘導酶存在,在淀粉量維持穩(wěn)定時,其酶活性變化不大。
3 結 語
茯苓菌固態(tài)發(fā)酵的過程中,發(fā)酵產(chǎn)物中的總糖、還原糖、多糖、三萜類物質(zhì)等質(zhì)量分數(shù)都有明顯變化。與未經(jīng)發(fā)酵的小麥相比,茯苓菌發(fā)酵小麥中的總糖質(zhì)量分數(shù)明顯下降,減少了53%;還原糖質(zhì)量分數(shù)顯著上升,由發(fā)酵初期的0.389%上升至27.40%;多糖質(zhì)量分數(shù)由8.72%上升至14.96%,發(fā)酵過程中最高可達26.83%;總?cè)婆c水溶性三萜的質(zhì)量分數(shù)逐漸提高,在培養(yǎng)至第25天時達到最高值,分別高達1.6 g/kg與0.13 g/kg。
通過對茯苓菌發(fā)酵小麥過程中茯苓菌酶活變化和基質(zhì)中營養(yǎng)成份變化的分析,可以看出茯苓菌在發(fā)酵的過程中,通過纖維素酶、木聚糖酶及淀粉酶等酶系的共同作用可將大分子物質(zhì)降解,改善小麥原有的營養(yǎng)結構。由于固態(tài)發(fā)酵方式工藝簡單、原料成本低,又因小麥本身的可食性,在進行后期加工時不需采取將菌絲分離的步驟,極大地簡化了加工流程。綜上所述,可將固態(tài)發(fā)酵技術與傳統(tǒng)谷物加工結合,開發(fā)具有高營養(yǎng)價值的谷物產(chǎn)品,既能促進傳統(tǒng)谷物的應用,又提升了其營養(yǎng)價值,其市場價值可期。
猜你喜歡
中學生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數(shù)理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數(shù)理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
