釀酒酵母萌發突變株的篩選
2019-04-24 07:45:36邱盼盼高曉冬中西秀樹
食品與生物技術學報 2019年12期
邱盼盼, 高曉冬, 中西秀樹
(江南大學 生物工程學院,江蘇 無錫 214122)
酵母細胞在缺乏氮源且遇到非發酵性碳源的情況下,酵母細胞就會進入減數分裂進行孢子生殖,將單倍體的染色體分到4個不同的子細胞中,形成孢子[1]。而孢子在遇到營養物質如葡萄糖時,開始萌發、發生形變,出芽長成正常的營養細胞[2-3]。孢子萌發是孢子經過休眠,非分裂性孢子的生長和進入有絲分裂的過程,而控制細胞進入有絲分裂的系統是個有序的選擇性系統,只有在遇到適合的情況才會退出休眠期進入有絲分裂[4],該選擇性系統涉及一系列蛋白質,影響到轉錄、蛋白質的磷酸化、催化子和抑制子的穩定性[5]。關于萌發機制則至今還未被詳細解析,因此本實驗對萌發機制的研究有理論意義。
孢子外層壁結構由里到外依次是甘露糖蛋白(mannoprotein),β-葡 聚糖 (β-glucan), 殼 聚糖(chitosan),以及二酪氨酸(dityrosine)層[6-7],而孢子最外層的二酪氨酸層對其乙醚抗性具有重要作用[8-9],而正常的營養細胞對乙醚缺乏抗性,本研究利用這一特性篩選萌發缺陷型突變株。
本研究室首次利用孢子作為微膠囊[10],建立2種新型固定化酶方法。其中一個方法是以孢子為載體[11],利用二酪氨酸的阻擋作用,將酵母中分泌型的酶固定在孢子壁上,其特點是固定化酶的穩定性高,能抵抗較高的溫度和培養環境水解酶的降解作用。另一種方法是將外層二酪氨酸層去除,使殼聚糖層暴露在孢子最外層形成殼聚糖球。利用殼聚糖層上的氨基,結合有效的化學方法將需要固定的外源酶共價交聯在殼聚糖層上,具有重復利用率高,熱穩定性好等特點。該殼聚糖球還具有吸附性強的特點,可去除廢水中的金屬離子和帶負電荷的離子,達到凈化環境的目的。但是上述2種方法的不足之處在于孢子遇到營養物質即開始萌發,不利于酶的固定化,為保證其重復利用性,本研究對釀酒酵母進行化學誘變,經篩選獲得若干株具有萌發缺陷的突變株。
1 材料與方法
1.1 材料
1.1.1 菌株 出發菌株釀酒酵母(Saccharomyces cerevisiae)AN120由本實驗室保藏和提供。
1.1.2 培養基 YPAD培養基(g/dL):2葡萄糖,2蛋白胨,1酵母提取物,0.003腺嘌呤,(固體培養基加2瓊脂);預產孢液體培養基(YPACe):2蛋白胨,1酵母提取物,2醋酸鉀;產孢培養基(KOAC):2醋酸鉀,(固體培養基加2瓊脂);萌發液體培養基YPA(g/dL):2蛋白胨,1酵母提取物,0.003腺嘌呤;選擇性培養基(SD)(g/dL):0.67不含氨基酸的酵母氮源,2瓊脂,2葡萄糖,2不含特定氨基酸的氨基酸混合物;大腸桿菌培養基(LB)(g/dL):0.5 酵母提取物,1胰蛋白胨,1氯化鈉,2瓊脂。
1.1.3 化學突變試劑和緩沖液 本實驗采用甲基磺酸乙酯(EMS)[12]對酵母細胞進行突變,再使用5 g/dL滅菌的硫代硫酸鈉(Sigma)解除突變后的藥物毒性,緩沖液使用滅菌的0.1 mol/L磷酸鈉緩沖液,pH7.0。
1.1.4 儀器與設備 (Pipet Lite XLS)移液槍、(CF16RXⅡ)臺式高速冷凍離心機,購于德國Eppendorf;(BA210)光學顯微鏡,購于美國 GE 公司;(CV334)超聲破碎儀,購于南京新辰生物科技公司。
1.2 方法
1.2.1 突變以及用乙醚篩選突變株 突變參照Fred Winston[12]的方法進行誘變,每 3.0×108個細胞使用150 μL的突變化學試劑EMS在30℃震蕩處理1 h,然后加入100 mL 5 g/dL sodium thiosfate進行解毒處理。
乙醚篩選:產孢后,將孢子懸浮在液體YPAD培養基中于37℃搖床內萌發14 h,滴加乙醚使其終體積分數13%,在30℃搖床中處理45 min,稀釋1 000倍后涂布在50個固體YPAD培養基上置于25℃恒溫箱內4 d以進行后續實驗。
1.2.2 子囊孢子的制備以及萌發步驟 在YPAD培養基中過夜培養細胞,上述液體預產孢培養基YPACe培養24 h后,在產孢培養基KOAC中培養1~2 d,收集孢子,加入2 g/dL lyticase,置于30℃搖床中2~3 h,收集孢子,用去離子水洗滌2次,溶于5 mL體積分數0.5%TritonX-100,使用超聲破碎儀進行破壁處理5 min,用去離子水洗滌2次。在光學顯微鏡下可見97%以上的細胞均為單個的孢子。萌發的步驟如下:將孢子置于YPA液體培養基于30℃下30 min,再加入葡萄糖至總質量濃度為2 g/dL。萌發率的計算則由每100個細胞計已萌發孢子數,測3次取平均值。
1.2.3 四分體孢子的分離 將新鮮的孢子,置于100μL的去離子水中,加0.5μL的lyticase置于30℃下15 min,將YPAD平板在邊緣處劃線,取10 μL的處理液置于一個線內的YPAD上,在操作臺內吹干處理液,將切孢的針用乙醇處理10 min,在顯微鏡下利用針尖將四分體的孢子分離開來置于該YPAD平板上。由于沒有添加裂解酶的四分體子囊孢子不能通過針尖取到,因此后續部分實驗處理子囊孢子時先將其分開再放置在一起。
2 結果與討論
2.1 突變方法的確立
突變流程見圖1,首先對細胞進行化學試劑突變處理,在細胞量回補后,經過正常的產孢,在高溫下37℃下萌發,使得沒有高溫萌發缺陷的孢子萌發成正常的營養細胞,而有缺陷的孢子依然保持孢子的結構,經過一定濃度的乙醚處理,將萌發后的營養細胞殺死,而保留對乙醚有抗性的孢子,然后置于25℃下,缺陷孢子開始萌發即得到萌發缺陷初選菌株。本實驗由于完全不能萌發的突變株不能被篩選到,因此本實驗假設突變株在高溫下不能萌發而在低溫下能萌發。其中測試乙醚篩選條件時,經過一系列不同濃度乙醚測試,發現處理以13%終體積分數的乙醚在30℃下45 min能殺死所有的萌發細胞,而保留一定數量的孢子存活。本實驗獲取3×108個酵母細胞,經過EMS誘變后,致死率約為50%~70%,最后經過乙醚篩選得到300株左右菌株。實驗通過對突變條件的測試,確立了通過化學試劑對釀酒酵母進行突變,結合利用乙醚篩選突變株的方法,成功獲得了具有萌發缺陷的突變株。
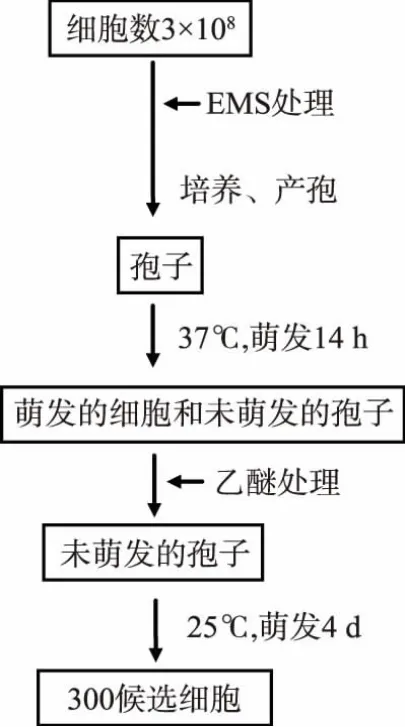
圖1 突變流程Fig.1 Mutation procedure
篩選最終得到約300株初選菌株,為排除生長缺陷菌株,將此300株菌株置于37℃下培養,其中103株可以生長。而為方便研究萌發的缺陷,本文主要針對能產孢的菌株以方便后續的研究,故此排除產孢有缺陷或者單倍體菌株,最后得到32株菌株能產孢的雙倍體菌株。為進一步確認這32株菌株的確有萌發缺陷,對這些菌株再進行萌發處理,發現有明顯萌發缺陷的只有4株,分別命名為AN120-m5、AN120-m8、AN120-m10、AN120-m17。
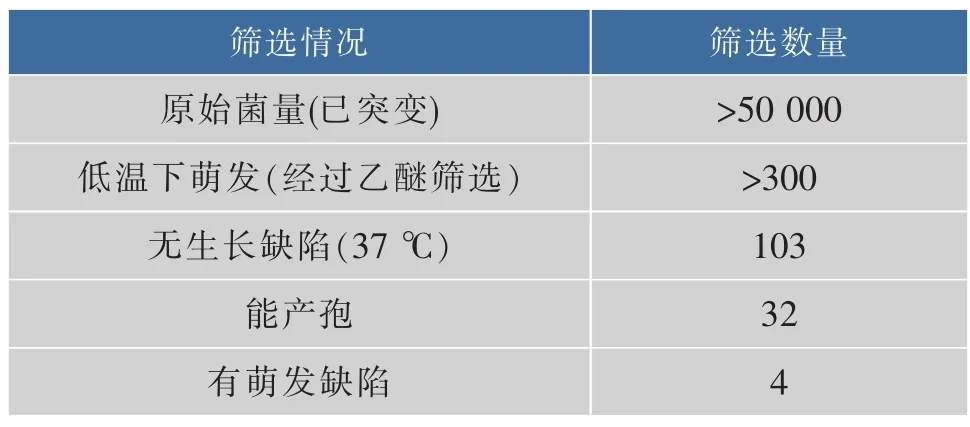
表1 篩選量Table 1 Screening volume
2.2 突變株驗證
上述實驗已證明,4株突變株均可以正常產孢,為進一步確認這4株突變株是在萌發過程中有缺陷,并非生長缺陷突變株,對固體培養基亦做了生長缺陷測試。將野生型和突變株在37℃下培養,由圖 2 可看出 AN120-m8,AN120-m10,AN120-m17相對于野生型來說沒有明顯的生長缺陷,而AN120-m5則表示為輕微生長劣勢,但是能夠在37℃下生長。實驗表明,4株突變株都能在37℃下生長。
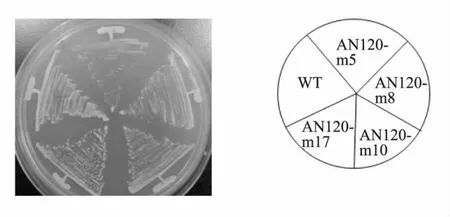
圖2 突變株37℃生長狀況Fig.2 Growth of mutants on YPAD plate in 37℃
為確認突變株在萌發過程中有缺陷,測試突變株在液體培養基內的缺陷進行驗證。如圖3(a)所示,經過8 h后,野生型孢子在37℃和25℃下均能正常萌發,而該4株突變株在25℃下和37℃下都有明顯的萌發缺陷,而尤其特殊的是AN120-m10突變株在25℃下和37℃下萌發率都極低,說明該突變株的萌發缺陷非常嚴重。之后,對突變株在固體培養基的缺陷進行驗證。將突變株單個孢子置于固體培養基上,將來自同一子囊的四個孢子放在一行,將6組單孢子組在固體YPAD于25℃中培養3~4 d后,4株突變體在固體培養基上萌發狀況如圖3(b)所示,能直觀的看到4株突變株在固體培養基上的萌發缺陷更為嚴重。而各突變株在37℃下除AN120-m17全不萌發,且AN120-m17萌發率也極低。并且野生型的單孢子在固體培養基上萌發只需2 d,而突變型則需要4 d。顯著觀察到的是AN120-m5的單個孢子在固體培養基上于任何溫度下都完全不萌發。結合上述實驗數據表明,4株突變株在產孢正常,也沒有生長缺陷的情況下,確實是在萌發過程中有缺陷,其孢子萌發時間較野生型更長。

圖3 驗證突變株在液體和固體培養基中的萌發率Fig.3 Germination efficiency of mutants in liquid medium and on solid medium
可見突變體AN120-m5的單個孢子在固體培養基中完全不能萌發,但是在液體培養基中,卻能夠萌發,考慮到在液體培養基中孢子的聚集性,于是將其子囊孢子的4個孢子緊緊貼近置于固體培養基YPAD上,發現萌發效率和單個孢子相比有所提高,如圖4,這可能是因為在AN120-m5中多個孢子聚集在一起,某種物質的生成能誘發萌發的發生,而導致萌發率提高。

圖4 AN120-m5 4個緊湊挨近的子囊孢子在固體培養基上的萌發Fig.4 Germination on solid plate of tetrad spores of AN120-m5
2.3 萌發缺陷穩定性驗證
為確保突變株的缺陷穩定性,驗證是否單個基因突變發生在突變株中,對突變株的孢子進行分離,將其與自身突變株的單倍體進行交配,獲得自交雙倍體。測驗結果由圖5可知,突變體AN120-m8、AN120-m10和AN120-m17的自交后代AN120-m8C、AN120-m3C、AN120-m4D,在 37℃下萌發率為38%、14%、43%。而由于AN120-m5突變株不能得到單倍體,所以不能交配得到自交雙倍體。實驗顯示,在自交后,萌發缺陷依然存在,說明突變性狀是穩定的。

圖5 自交雙倍體萌發率Fig.5 Germination efficiency of self-crossed mutant diploid cells
為了進一步驗證萌發缺陷的穩定性,在和野生型單倍體雜交三代后,測試在液體培養基內的萌發率,由圖6可知,同一個平板上切割的雜交雙倍體的孢子,為清楚確認生長良好的菌落確是來自于野生型,取生長良好菌落和野生型單倍體交配,發現其和野生型孢子萌發情況相同。實驗證明,萌發狀況良好的單菌落是來自野生型,而萌發狀況不良的單菌落則是來自突變型。由圖6可以明顯看到,每個四分體的2個突變型孢子在25℃下經過三代雜交后萌發情況始終較野生型的2個孢子弱,呈現出萌發缺陷的性狀。由圖7可知AN120-m8、AN120-m10和AN120-m17的第三代雜交雙倍體AN120-G38、AN120-G310、AN120-G317在 37 ℃萌發率分別為45%、48%和53%,較原始菌株有所提高,但是始終低于野生型。實驗進一步證明,萌發缺陷穩定存在突變型菌株中,不隨著產孢過程中遺傳物質的分離3次后消失,顯著表現為野生型與突變型孢子的生長優良狀況始終呈現出2∶2的狀態,進一步證明突變株是單基因突變菌株的可能性很大。

圖6 第三代雜交雙倍體切割孢子Fig.6 Dissection of the third gerneration hybrid diploid cells

圖7 第三代雜交雙倍體萌發率Fig.7 Germination of the third gerneration hybrid diploid cells
2.4 討論
本實驗室研究利用孢子作為載體進行酶的固定化,將酶固定在孢子壁外層二酪氨酸層內以及利用化學方法將酶共價交聯在去除二酪氨酸層的殼聚糖球體上,得到新型固定化酶型孢子。但是經過固定化的孢子其中一個缺陷是孢子遇到營養物質將萌發,為了確保固定化酶的重復使用性,同時利于研究萌發機制,本研究通過化學試劑對釀酒酵母細胞進行誘變,旨在獲取萌發缺陷菌株。本實驗篩選方法的特征在于利用孢子對乙醚的抗性,保護了在37℃下未萌發的突變型孢子,再在25℃下得以萌發,經過進一步篩選,成功地從3×108個細胞中篩選到約300株初選菌株,并且通過各種驗證,最終獲得了4株目的菌株。由于上述突變株在液體培養基中有嚴重的萌發缺陷,初步表明符合微膠囊的應用條件。
有研究者用切孢技術將四倍體孢子分離[13],在本研究中,使用顯微鏡和針尖將孢子分離獲得突變體單倍體,不僅直觀看到在固體培養基上突變株的萌發缺陷更為明顯,并且明顯觀察到AN120-m5突變體單個孢子在固體培養基上完全不能萌發,這對于后續篩選突變基因具有重要意義。而該突變體在液體培養基中能萌發,但是其單個孢子不能在固體培養基上萌發,并且多個孢子聚集在一起時,其四分體孢子在固體培養基上萌發率提高,這可能是由于孢子分離的狀態影響了萌發的發生,該突變體的孢子可能只以子囊的形式萌發,其具體的原因待進一步探究。
為驗證突變株萌發缺陷的穩定性,從而確認突變的來源是否來自單個基因上。在同時測試雜交雙倍體和自交雙倍體的萌發率后,實驗證明,在與野生型單倍體雜交三代后,其野生型孢子和突變型孢子生長優劣狀況始終呈現2∶2的比例,缺陷依然存在,不隨著產孢后遺傳物質的分離而消失,證明缺陷穩定存在突變株內,所以4株突變株是單基因突變菌株有很大的可能性。4株突變株性狀各有不同,而且特別顯著的是AN120-m5的性狀和其余3株有很大差異,至少說明經過后續篩選我們可以獲得2個以上的突變基因。
3 結 語
本研究成功利用甲基磺酸乙酯進行突變處理,并利用孢子對乙醚的抵抗性確定了一種篩選萌發突變株的方法,并以此找到了4株在固體培養基和液體培養基同時具有萌發缺陷的突變株,并且萌發缺陷穩定存在。由于單基因突變的可能性很大,后續實驗利用確定找到發生突變的基因,從而達到敲除突變基因獲得完全不萌發的突變菌株,并研究用于工業固定化酶的工程菌株以及為萌發機制。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
