調(diào)控黑曲霉葡萄糖轉(zhuǎn)運(yùn)系統(tǒng)增強(qiáng)檸檬酸的合成
2019-04-24 07:45:32李江華堵國成
食品與生物技術(shù)學(xué)報 2019年12期
殷 嫻 , 李江華 , 劉 龍 , 堵國成 , 陳 堅 *
(1.江南大學(xué) 工業(yè)生物技術(shù)教育部重點實驗室,江蘇 無錫 214122;2.江南大學(xué) 生物工程學(xué)院,江蘇 無錫214122)
檸檬酸 (citrate)是最具商業(yè)價值的有機(jī)酸之一,年產(chǎn)量達(dá)到170萬噸。黑曲霉(Aspergillus niger)是主要的檸檬酸生產(chǎn)菌[1],然而根據(jù)Alvarez-Vasquez的模型,檸檬酸生產(chǎn)強(qiáng)度還可以有5倍的提高空間[2]。目前黑曲霉的代謝工程改造策略圍繞中心代謝通路和呼吸鏈展開[3],表達(dá)糖酵解途徑[4-5]和TCA循環(huán)[6-8]的關(guān)鍵酶以及交替氧化酶[9]均無法提升檸檬酸產(chǎn)量,僅加強(qiáng)rTCA循環(huán)[10]對檸檬酸合成有促進(jìn)作用。在檸檬酸工業(yè)生產(chǎn)中,原料為淀粉,淀粉經(jīng)過淀粉酶液化后過濾,得到清液和混液,通過清液和混液進(jìn)行勾兌得到種子和發(fā)酵培養(yǎng)基,此時培養(yǎng)基中約一半的碳源為葡萄糖,一半碳源以多聚葡萄糖的形式存在,而工業(yè)生產(chǎn)菌在發(fā)酵初期合成大量糖化酶,將多聚葡萄糖分解為葡萄糖,因此,在發(fā)酵前期培養(yǎng)基中碳源已經(jīng)基本以葡萄糖的形式存在,所以檸檬酸發(fā)酵對碳源的吸收實際上就是對葡萄糖的吸收。Torres的研究發(fā)現(xiàn)黑曲霉中存在兩類葡萄糖轉(zhuǎn)運(yùn)蛋白,一為高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白,Km值約為260 μmol/L,另一類為低親和力的葡萄糖轉(zhuǎn)運(yùn)蛋白,Km值約為3.67 mmol/L[11]。低親和力葡萄糖轉(zhuǎn)運(yùn)蛋白可以提供檸檬酸發(fā)酵足夠的代謝通量,但該系統(tǒng)僅在葡萄糖濃度高于50 g/L時起作用[11]。葡萄糖轉(zhuǎn)運(yùn)是檸檬酸發(fā)酵的第一步,調(diào)整葡萄糖轉(zhuǎn)運(yùn)系統(tǒng)對檸檬酸發(fā)酵的影響值得研究。目前黑曲霉中已經(jīng)鑒定到一個高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白mstA[12],該蛋白質(zhì)在產(chǎn)酶黑曲霉CBS513.88和產(chǎn)酸原始菌株ATCC1015中存在,而在黑曲霉檸檬酸生產(chǎn)菌株H915-1中不存在,因此,H915-1中的高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白需要進(jìn)行鑒定。
在真菌中存在很多假定的葡萄糖轉(zhuǎn)運(yùn)蛋白,它們歸類于主要協(xié)同轉(zhuǎn)運(yùn)蛋白超家族(major facilitator superfamily,MFS)[13-14],可以是單向轉(zhuǎn)運(yùn)蛋白或 H+同向轉(zhuǎn)運(yùn)蛋白,有些可以轉(zhuǎn)運(yùn)多種底物。釀酒酵母(Saccharomyces cerevisiae)的葡萄糖轉(zhuǎn)運(yùn)目前研究得最透徹,含有至少20個轉(zhuǎn)運(yùn)蛋白參與葡萄糖的轉(zhuǎn)運(yùn)[15],一些轉(zhuǎn)運(yùn)蛋白具有信號轉(zhuǎn)導(dǎo)功能[16-17]。在構(gòu)巢曲霉(A.nidulans)基因組中預(yù)測到了17個己糖轉(zhuǎn)運(yùn)蛋白[18],白念珠菌(Candida albicans)基因組中含有20個假定的葡萄糖轉(zhuǎn)運(yùn)蛋白[19]。在真菌中存在多種葡萄糖轉(zhuǎn)運(yùn)蛋白,說明特定條件下轉(zhuǎn)運(yùn)系統(tǒng)是多個基因差異性表達(dá)的結(jié)果。每一個轉(zhuǎn)運(yùn)蛋白有其特定的底物、親和力和轉(zhuǎn)運(yùn)能力,調(diào)整轉(zhuǎn)運(yùn)蛋白可以使賦予菌體營養(yǎng)攝入的靈活性,使之適應(yīng)多樣的營養(yǎng)環(huán)境。
黑曲霉H915-1基因組中預(yù)測到21個葡萄糖轉(zhuǎn)運(yùn)蛋白,其中1個低親和力葡萄糖轉(zhuǎn)運(yùn)蛋白在整個發(fā)酵過程中高表達(dá),5個高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白低表達(dá)(GEO號GSE74544)。本研究對一個假定的葡萄糖轉(zhuǎn)運(yùn)蛋白的功能進(jìn)行鑒定,最終通過過表達(dá)轉(zhuǎn)運(yùn)蛋白提升了檸檬酸產(chǎn)量。
1 材料與方法
1.1 菌株和培養(yǎng)基
大腸桿菌DH5α用作載體構(gòu)建的宿主菌。黑曲霉H915-1為江蘇國信協(xié)聯(lián)能源有限公司 (檸檬酸產(chǎn)量居中國第三位)的檸檬酸生產(chǎn)菌株,作為出發(fā)菌株。
大腸桿菌轉(zhuǎn)化子利用含100 μg/mL氨芐青霉素的LB培養(yǎng)基進(jìn)行篩選,黑曲霉轉(zhuǎn)化子利用含150 μg/mL潮霉素的ME培養(yǎng)基 (3%麥芽提取物,0.5%蛋白胨)進(jìn)行篩選。檸檬酸發(fā)酵培養(yǎng)基為玉米清液和玉米混液的混合物,含15%總糖和0.08%總氮。不同葡萄糖濃度的發(fā)酵實驗使用合成培養(yǎng)基 (3 g/L NH4NO3,0.1 g/L KH2PO4,1 g/L MgSO4·7H2O,0.015 g/L CuSO4·5H2O,0.001 67 g/L FeCl3·6H2O,0.008 g/L ZnSO4·7H2O 和 3.74 g/L CaCl2),葡萄糖濃度分別調(diào)整為3%、5%、8%和10%。葡萄糖限制性培養(yǎng)基中的葡萄糖質(zhì)量濃度則分別調(diào)整為0.05、0.1 g/L和0.5 g/L。
1.2 實驗方法
1.2.1 表達(dá)載體的構(gòu)建 共表達(dá)所用表達(dá)框包含hph抗性基因表達(dá)框以及AnHGT1表達(dá)框。載體pUCDT利用Kpn I和Bam HI進(jìn)行酶切。HGT1基因(NCBI序列號 XM_001399160.2) 以 H915-1的cDNA為模板,利用引物gpd-HGT-F和Trp-HGT-R進(jìn)性RT-PCR擴(kuò)增,序列兩端含有20 bp pUCDT同源序列,利用Vazyme One Step Clone Kit進(jìn)行同源重組形成載體pGTH。隨后pGTH利用Spe I和Not I進(jìn)行酶切,并與用相同限制性內(nèi)切酶酶切的pHPH連接,形成pHGTH,包含了2個表達(dá)框。利用引物pMD-F和pMD-R以pHGTH為模板,PCR擴(kuò)增得到共表達(dá)框。所用引物序列見表1。共表達(dá)框示意圖見圖1。
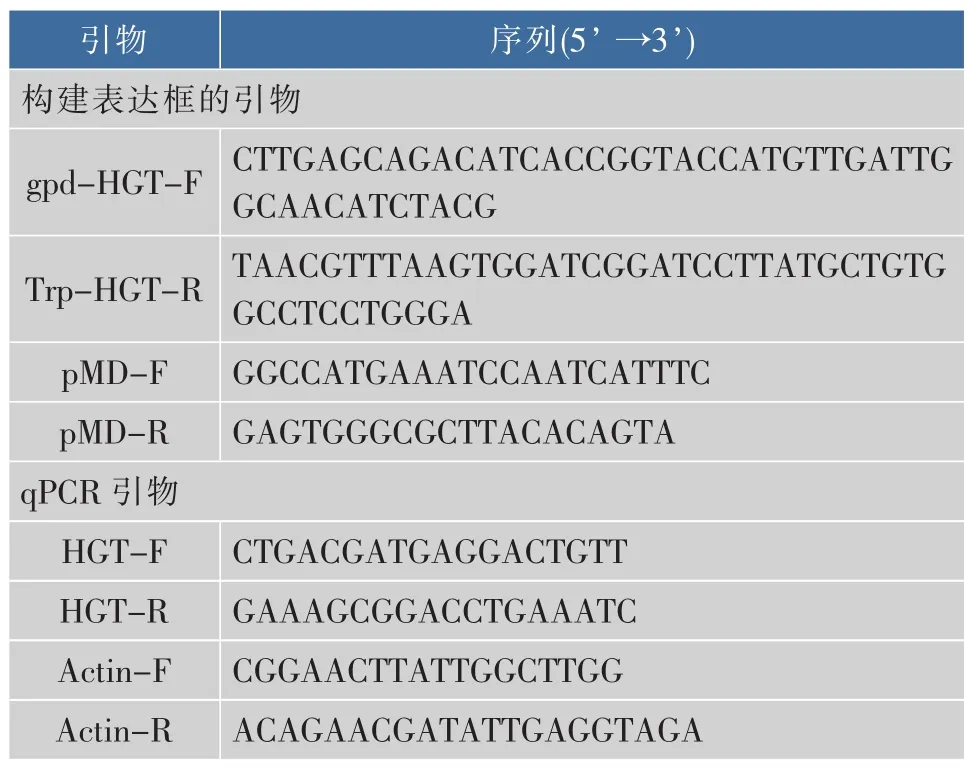
表1 引物序列Table 1 Primer sequences

圖1 HGT1雙元表達(dá)框示意圖Fig.1 Structure of HGT1 binary expression cassette
1.2.2 進(jìn)化樹分析 利用Clustal Omega軟件對DNA序列進(jìn)行比對,利用MEGA5進(jìn)行進(jìn)化樹分析[20]。
1.2.3 跨膜區(qū)域預(yù)測 利用在線軟件TMHMM Server v.2.0 (www.cbs.dtu.dk/services/TMHMM/),對序列跨膜區(qū)域進(jìn)行預(yù)測。
1.2.4 黑曲霉H915-1的轉(zhuǎn)化 通過制備原生質(zhì)體[21],利用PEG介導(dǎo)法進(jìn)行轉(zhuǎn)化[22]。所有的轉(zhuǎn)化子在抗性培養(yǎng)基上經(jīng)過3次單克隆培養(yǎng)進(jìn)行純化,純化后的菌株進(jìn)行qPCR鑒定。
1.2.5 黑曲霉H915-1的檸檬酸搖瓶發(fā)酵 黑曲霉種子培養(yǎng):將黑曲霉孢子懸液以終濃度4×105個/mL接種至種子培養(yǎng)基,在35℃轉(zhuǎn)速250 r/min培養(yǎng)24 h。
搖瓶發(fā)酵:將種子培養(yǎng)液以體積分?jǐn)?shù)10%接種量接種至發(fā)酵培養(yǎng)基以250 r/min、35℃培養(yǎng)至還原糖耗盡,測定檸檬酸產(chǎn)量。
1.2.6 HPLC檢測有機(jī)酸 收集樣品后,利用miracloth過濾去除菌球,溶液在4℃經(jīng)12 000 r/min離心10 min,取上清液經(jīng)過相應(yīng)稀釋后,用0.22 μm濾膜過濾。利用Aminex 87-H有機(jī)酸柱進(jìn)行HPLC檢測,流動相 5 mmol/L H2SO4,流速 0.6 mL/min,溫度35℃,利用紫外檢測器在210 nm處進(jìn)行檢測。
1.2.7 總糖和還原糖的測定 DNS法測定還原糖:配制 0.2、0.4、0.6、0.8、1.0 g/L 的葡萄糖溶液。分別取葡萄糖溶液(或樣品)1.0 mL,與1 mL DNS混勻,沸水浴加熱 15 min,冰浴冷卻后稀釋定容至10 mL,測OD540。
總糖的測定:取5 mL發(fā)酵液至25 mL三角瓶中,加入6 mol/L HCl 5 mL,在沸水浴中加熱25 min進(jìn)行水解,冷卻至室溫。將溶液轉(zhuǎn)移到50 mL燒杯中,調(diào)pH至7~8之間,定容至100 mL,過濾,適當(dāng)稀釋后用DNS法測定還原糖。
還原糖的檢測。利用3,5-二硝基水楊酸(DNS)法檢測還原糖含量。
未分解還原糖含量=總糖含量-還原糖含量。
1.2.8 qPCR檢測檸檬酸發(fā)酵過程HGT1基因的表達(dá) 黑曲霉發(fā)酵8 h和24 h時快速取樣,用miracloth收集菌體,并用無菌去離子水清洗并壓干,置于液氮中凍存。用RNeasy Plant Mini Kit參照說明書進(jìn)行樣品總RNA的抽提,利用Takara公司PrimeScript RT Reagent Kit with gDNA Eraser進(jìn)行反轉(zhuǎn)錄,qPCR所用引物見表1,引物Tm值為55℃,產(chǎn)物長度為120~150 bp,內(nèi)參為actin。誤差線為3個生物學(xué)重復(fù)。
2 結(jié)果與討論
2.1 檸檬酸發(fā)酵過程中葡萄糖吸收速率的變化
對黑曲霉H915-1檸檬酸發(fā)酵的總糖和還原糖進(jìn)行測定。檸檬酸發(fā)酵前,原料淀粉經(jīng)淀粉酶液化后部分分解,初始還原糖質(zhì)量濃度約為9%,未分解多糖在發(fā)酵過程中由黑曲霉自分泌的糖化酶進(jìn)一步降解。因此,發(fā)酵前期的還原糖質(zhì)量濃度增加,為避免糖化酶分泌不足造成培養(yǎng)基中的淀粉持續(xù)分解,發(fā)酵后期的還原糖生成影響糖耗速率檢測。在一組接種種子培養(yǎng)基的同時添加過量糖化酶(糖化酶酶活為 2.5 g/(mL·min),添加量為 40 μL),另一組則不添加糖化酶。2組均在16 h時還原糖含量達(dá)最高值,添加糖化酶后培養(yǎng)基中的未分解多糖在16 h消化完全,未添加糖化酶的培養(yǎng)基則在24 h將未分解多糖完全消化(圖2)。還原糖的消耗速率均在發(fā)酵后期放緩,從80 h的2.09 g/L/h逐步下降104 h的 1.16 g/(L·h), 至116 h糖消耗速率僅為 0.35 g/(L·h),因此需解決糖耗速率放緩的問題。由于黑曲霉產(chǎn)酸階段主要由低親和力的葡萄糖轉(zhuǎn)運(yùn)蛋白承擔(dān)吸收葡萄糖,在發(fā)酵后期,隨著葡萄糖濃度的減少,將逐步過渡到高親和力的葡萄糖轉(zhuǎn)運(yùn)蛋白接管葡萄糖的吸收,因此有必要研究葡萄糖轉(zhuǎn)運(yùn)系統(tǒng)對檸檬酸合成的影響。

圖2 檸檬酸發(fā)酵過程中還原糖、未分解多糖質(zhì)量濃度及檸檬酸質(zhì)量濃度Fig.2 Concentration of reducing sugar,undegraded sugar and citrate titer during fermentation
2.2 葡萄糖轉(zhuǎn)運(yùn)蛋白的進(jìn)化樹分析、多序列比對及跨膜預(yù)測
對黑曲霉H915-1檸檬酸發(fā)酵過程中在轉(zhuǎn)錄水平獲得表達(dá)的糖轉(zhuǎn)運(yùn)蛋白采用進(jìn)化樹分析 (圖3),參考序列為Swiss-Prot數(shù)據(jù)庫所有高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白。ScHXT2為釀酒酵母的高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白[23],并沒有鑒定到與之親緣關(guān)系較近的蛋白。ScSnf3為釀酒酵母的高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白,同時是葡萄糖轉(zhuǎn)運(yùn)的負(fù)調(diào)節(jié)因子,在低葡萄糖濃度下通過抑制RGT1參與誘導(dǎo)HXT2的表達(dá)[15],evm.model.unitig_4.946與之親緣關(guān)系較近。KlHGT1為乳酸克魯維酵母(Kluyveromyces lactis)的高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白,同時可以轉(zhuǎn)運(yùn)半乳糖[24],evm.model.1.1149、evm.model.unitig_0.1770 和 evm.model.unitig_3.1098與之親緣關(guān)系較近;進(jìn)一步對這4條序 列 進(jìn) 行 比 對 ,evm.model.1.1149、evm.model.unitig_0.1770和evm.model.unitig_3.1098與KlHGT1的同源性分別為41.7%、41.0%和43.1%(圖4)。隨后對evm.model.unitig_0.1770進(jìn)行跨膜預(yù)測,發(fā)現(xiàn)該蛋白含有11個跨膜區(qū)域,N端在細(xì)胞膜內(nèi),C端預(yù)測在胞內(nèi)(圖5),對其命名為AnHGT1。
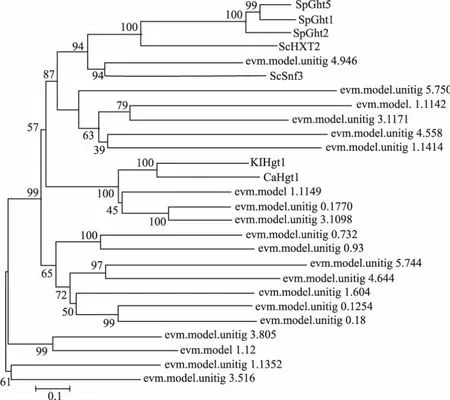
圖3 葡萄糖轉(zhuǎn)運(yùn)蛋白序列的進(jìn)化樹分析Fig.3 Phylogenetic analysis of the sequences of glucose transporters by Neighbour-joining(NJ) method
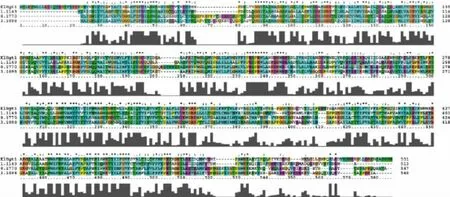
圖4 葡萄糖轉(zhuǎn)運(yùn)蛋白的多重序列比對分析Fig.4 Multiple sequence alignment of glucose transporters
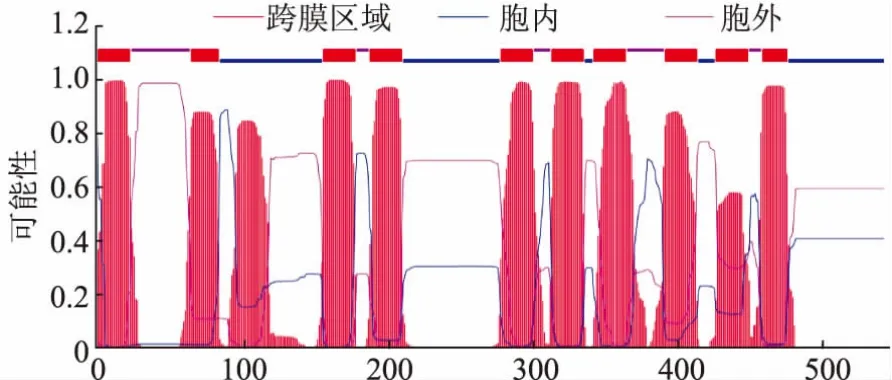
圖5 evm.model.unitig_0.1770的跨膜預(yù)測Fig.5 Transmembrane helices in evm.model.unitig_0.1770
2.3 黑曲霉HGT1轉(zhuǎn)化子的檸檬酸發(fā)酵
克隆黑曲霉自身的HGT1基因并構(gòu)建表達(dá)框以轉(zhuǎn)化黑曲霉H915-1,得到HGT1轉(zhuǎn)化子。對轉(zhuǎn)化子進(jìn)行檸檬酸發(fā)酵,利用qPCR檢測HGT1基因的表達(dá) (圖6),以H915-1發(fā)酵8 h的HGT1表達(dá)量為1,H915-1中的HGT1基因始終弱表達(dá);HGT1轉(zhuǎn)化子的基因在8 h的相對表達(dá)量是H915-1的44.5倍,24 h的相對表達(dá)量則下降為22.4倍,證明HGT1基因在轉(zhuǎn)化子中的表達(dá),同時由于HGT1的啟動子為PgpdA,說明PgpdA在檸檬酸發(fā)酵中啟動基因表達(dá)能力有所減弱。發(fā)酵前期還原糖生成速度放緩,意味著糖化酶的合成量有所降低,由于糖化酶組成型表達(dá),因此與菌體量相耦聯(lián),可能HGT1的大量表達(dá)反而使菌體生長放緩,從發(fā)酵40 h開始,HGT1轉(zhuǎn)化子的還原糖消耗快于H915-1,最終轉(zhuǎn)化子的檸檬酸產(chǎn)量為145.2 g/L,比H915-1增加了14.7%,發(fā)酵時間縮短了 6 h(圖 7(a)),最大比產(chǎn)酸速率提升了29.5%(圖7(b))。由于發(fā)酵中后期,HGT1的表達(dá)量比較于菌體生長階段減弱,可能HGT1的表達(dá)量需要在合適的范圍內(nèi)才有利于菌體利用碳源。

圖6qPCR檢測HGT1基因的表達(dá)Fig.6 qPCR for varification of HGT1 gene expression
進(jìn)一步檢測HGT1轉(zhuǎn)化子在不同濃度碳源下的檸檬酸發(fā)酵,如圖8(a)所示,在復(fù)合培養(yǎng)基中,初始葡萄糖濃度從3%逐步提升至10%,HGT1轉(zhuǎn)化子的檸檬酸產(chǎn)量始終比H915-1提高6%左右,表明HGT1的表達(dá)有利于檸檬酸的合成。利用合成培養(yǎng)基,在發(fā)酵120 h碳源耗盡的情況下,添加30 g/L葡萄糖到培養(yǎng)基中,檢測葡萄糖的消耗,由圖8(b)可知,HGT1轉(zhuǎn)化子對葡萄糖的吸收快于H915-1,轉(zhuǎn)化子完全消耗葡萄糖花費(fèi)了26 h,而H915-1則在38 h時依然可檢測到微量的葡萄糖,證明了HGT1的表達(dá)蛋白具有葡萄糖轉(zhuǎn)運(yùn)的功能,在低濃度碳源條件下促進(jìn)了葡萄糖的吸收。同時,HGT1轉(zhuǎn)化子的檸檬酸產(chǎn)量始終高于H915-1,表明促進(jìn)葡萄糖的轉(zhuǎn)運(yùn)可進(jìn)一步促進(jìn)H915-1合成檸檬酸。
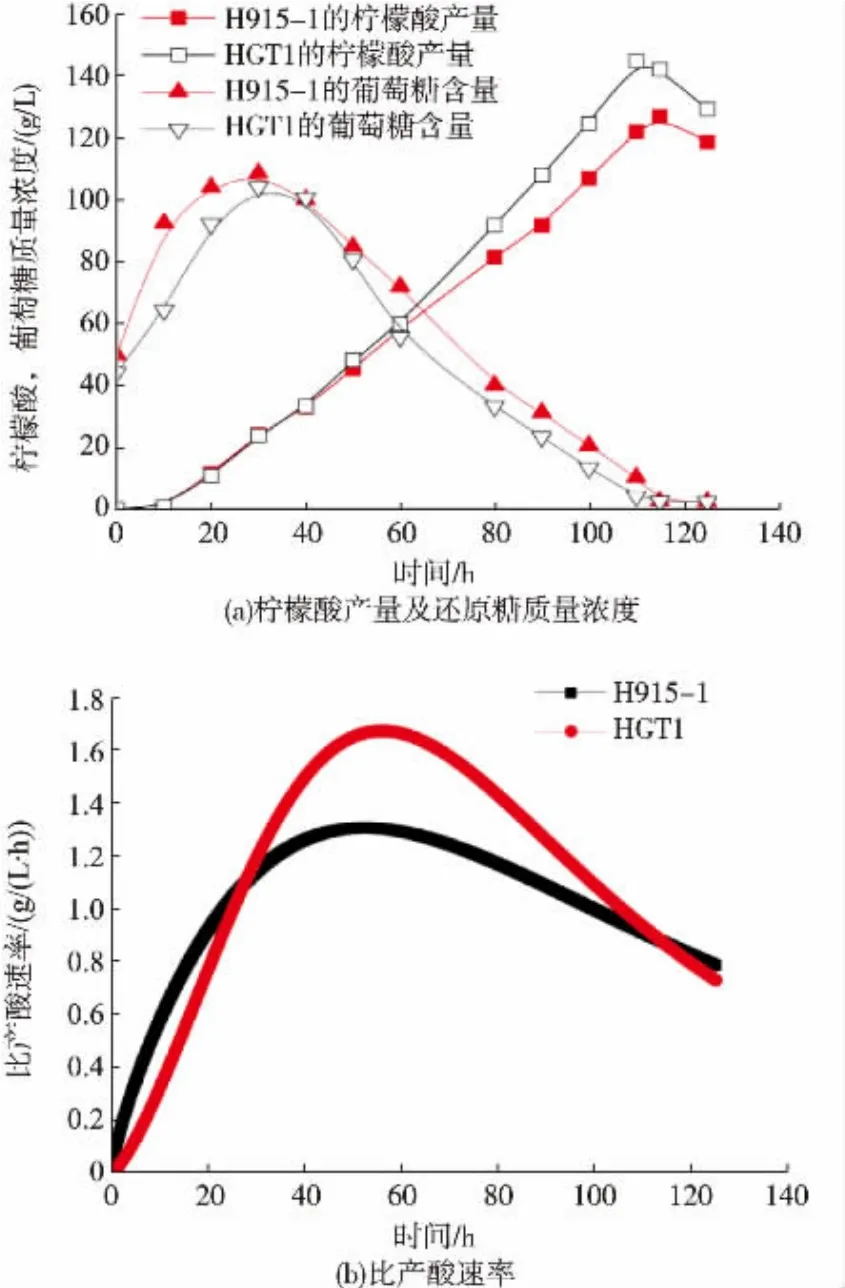
圖7 HGT1轉(zhuǎn)化子的檸檬酸發(fā)酵Fig.7 Citrate fermentation of HGT1 transformants


圖8 不同條件下的檸檬酸發(fā)酵Fig.8 Citrate fermentation under different conditions
2.4 HGT1對黑曲霉在葡萄糖限制性培養(yǎng)基上生長的影響
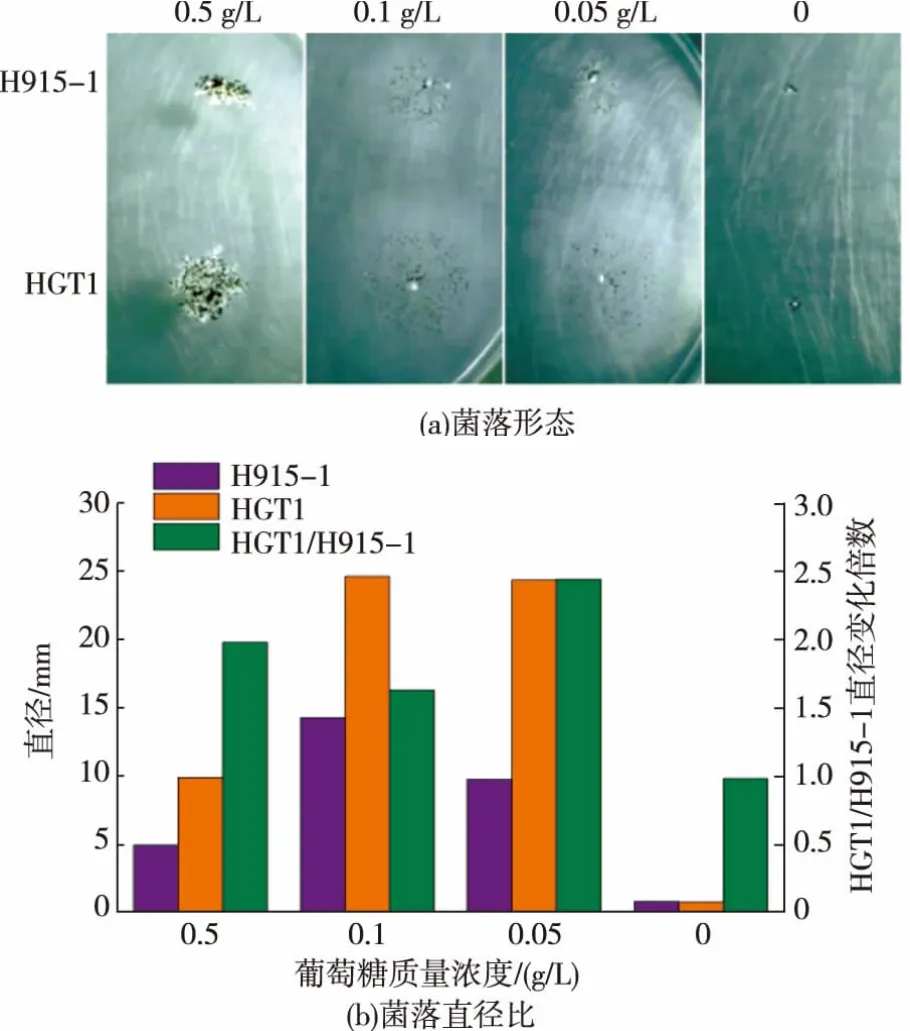
圖9 葡萄糖限制性培養(yǎng)基上菌落的生長Fig.9 Growth of strains in limited glucose media
在平板上涂布稀釋的黑曲霉孢子,在34℃靜置培養(yǎng)24 h至長出單菌落,將單菌落接種于低濃度碳源培養(yǎng)基上,34℃培養(yǎng)4 d,測量菌落直徑,檢測HGT1對黑曲霉生長的影響,由圖9(a)可知,在各質(zhì)量濃度葡萄糖限制性培養(yǎng)基上[25],HGT1轉(zhuǎn)化子的菌落生長強(qiáng)于H915-1。轉(zhuǎn)化子和H915-1的菌絲形態(tài)沒有明顯差異,在0.5 g/L葡萄糖質(zhì)量濃度下形成致密菌落,菌落直徑較小,顯微鏡下顯示菌絲為短粗狀,孢子囊較大;降低碳源質(zhì)量濃度至0.1 g/L導(dǎo)致菌絲成發(fā)散的細(xì)絲狀,并貼在培養(yǎng)基表面生長以吸收貧瘠的碳源,孢子囊細(xì)小,但和菌絲一樣分散生長,使菌落直徑比0.5 g/L葡萄糖時更大。但進(jìn)一步降低葡萄糖質(zhì)量濃度并沒有擴(kuò)大菌落,因為菌體生長受碳源限制而更緩慢。由菌落直徑比(圖9(b))可知,HGT1轉(zhuǎn)化子在各質(zhì)量濃度葡萄糖限制性培養(yǎng)基上生長的菌落直徑比H915-1增加約50%~150%,說明HGT1的組成型表達(dá)有利于菌體在低質(zhì)量濃度葡萄糖培養(yǎng)基上生長,意味著糖轉(zhuǎn)運(yùn)速度的加快,證明HGT1蛋白具有轉(zhuǎn)運(yùn)葡萄糖的功能。
3 結(jié)語
本文作者對黑曲霉H915-1基因組中假定的葡萄糖轉(zhuǎn)運(yùn)蛋白進(jìn)行了研究,通過進(jìn)化樹分析、序列比對分析,獲得與KlHGT1親緣關(guān)系較近的evm.model.unitig_0.1770序列,經(jīng)跨膜預(yù)測該蛋白含有11個跨膜區(qū)域,N端在細(xì)胞膜內(nèi),C端在胞內(nèi),將其命名為AnHGT1。在H915-1中組成型表達(dá)HGT1基因,轉(zhuǎn)化子的檸檬酸產(chǎn)量比對照增加了14.7%,發(fā)酵時間縮短了6 h,最大比產(chǎn)酸速率提升了29.5%。且在不同碳源濃度的培養(yǎng)基中,HGT1轉(zhuǎn)化子的檸檬酸產(chǎn)量均高于H915-1,表明增加葡萄糖吸收速度有利于強(qiáng)化檸檬酸合成。葡萄糖補(bǔ)加實驗表明HGT1轉(zhuǎn)化子完全消耗葡萄糖的時間比H915-1縮短了12 h,證明了HGT1蛋白具有轉(zhuǎn)運(yùn)葡萄糖的功能。在限制性葡萄糖培養(yǎng)基上的生長實驗,發(fā)現(xiàn)HGT1轉(zhuǎn)化子的菌落直徑比對照增加50%~150%,進(jìn)一步證明HGT1具有轉(zhuǎn)運(yùn)葡萄糖的功能。本研究提高了黑曲霉菌株生產(chǎn)檸檬酸的產(chǎn)量,對檸檬酸產(chǎn)業(yè)具有積極意義,并且確定了新的高親和力葡萄糖轉(zhuǎn)運(yùn)蛋白的功能,豐富了對黑曲霉葡萄糖轉(zhuǎn)運(yùn)系統(tǒng)的認(rèn)識,為后續(xù)改造黑曲霉提供了依據(jù)。
