蒙古國傳統發酵乳制品中保加利亞乳桿菌耐藥性的研究
2019-04-24 07:45:32李麗娜郭慧玲任彩霞張和平
食品與生物技術學報 2019年12期
關鍵詞:耐藥
李麗娜 , 郭慧玲 , 任彩霞 , 張和平 *
(1.內蒙古農業大學 乳品生物技術與工程教育部重點實驗室,內蒙古 呼和浩特 010018;2.內蒙古農業大學,奶制品加工農業部重點實驗室,內蒙古 呼和浩特010018)
乳酸菌是一類能夠利用可發酵糖產生50%以上乳酸的革蘭氏陽性細菌的通稱。廣泛地存在于自然界、動植物體、乳制品及發酵食品中,其中多為非致病菌且具有降血壓、改善腸道菌群、降低血液膽固醇等諸多益生功效[1]。在現代醫學中,抗生素是防御細菌感染的主要手段。雖然大多數乳酸菌被公認為是安全的食用型菌種,廣泛的應用于食品和藥品等領域中。但因過度使用和濫用抗生素,致使一些乳酸菌出現抗生素耐藥性,乳酸菌如果通過某種方式獲得耐藥基因,耐藥基因可以與人群、動物、生態系統中的細菌互相傳遞,致使出現耐藥致病菌,一旦將耐藥性傳遞給人類,將會對繼續使用抗生素來治療細菌性疾病造成一定的威脅[2]。世界衛生組織(WHO)確定抗生素耐藥性已經成為21世紀主要的健康性問題之一[3]。早期對細菌耐藥性的研究主要集中在病原微生物上[4-5],而近年來乳酸菌耐藥性的研究逐步成為人們關注的焦點。
乳桿菌屬是乳酸菌類群中最大的一個屬,在腸道正常微生物體系中有著不可或缺的地位,它能夠通過重組腸道菌群平衡來使人類及某些動物宿主恢復并達到健康狀態[6]。保加利亞乳桿菌(Lactobacillus bulgaricus)又稱為德氏乳桿菌保加利亞亞種(lactobacillus delbrueckii subsp.bulgaricus), 具有抗癌抗腫瘤、提高機體免疫力、促進胃腸道蠕動等益生功能[7]。乳酸菌的耐藥性主要包括兩種:固有耐藥性和獲得耐藥性。固有耐藥性是其本身所具有的特性,一般不會發生轉移,所以不必考慮其安全性,獲得性耐藥性是由基因突變或通過外源而獲得的耐藥基因。保加利亞乳桿菌是乳桿菌屬在食品工業中常用的菌種之一,所以有必要在使用前對菌株的表型及基因型進行分析以確保其安全性。
本實驗首先通過標準化的肉湯稀釋法來測定分離自蒙古國傳統發酵乳制品中5株保加利亞乳桿菌對15種抗生素的耐藥性,同時利用PCR技術來檢測菌株中可能攜帶的耐藥基因,以便于為以后菌種的選育及安全性評價提供重要的數據支持。
1 材料與方法
1.1 菌株來源
本實驗選取的5株保加利亞乳桿菌IMAU20289、IMAU20360、IMAU20427、IMAU20514、IMAU20635全部由內蒙古農業大學乳品生物技術與工程教育部重點實驗室菌種保藏庫提供。所有菌株通過16S rDNA序列分析均已鑒定。根據ISO10932/IDF223標準,37℃培養的乳桿菌選取Lactobacillus paracasei ATCC334做質控菌[8],是為了能夠更好的控制藥敏實驗的準確度和精確度。
1.2 培養基與試劑
LSM培養基(包含體積分數90%的ISO-SENSITEST和體積分數 10%的 MRS Broth)、MRS合成培養基(OXOID,CM0359)。
實驗用抗生素:卡那霉素 (KAN)、利奈唑胺(LINE),購買于 Chembase公司;奎奴普丁/達福普汀(QVI/DAL),購買于Bioaustralis公司,慶大霉素(GEN)、鏈霉素(STR)、新霉素(NEO)、四環素(TET)、紅霉素(ERY)、克林霉素(CLI)、氯霉素(CHL)、氨芐西林(AMP)、萬古霉素(VAN)、甲氧芐啶(VAN)、利福平(RIF)和環丙沙星(CIP),均購于 Sigma-Aldrich 公司。
試劑:1 g/dL瓊脂糖凝膠、5×TBE電泳緩沖液貯液、TE 緩沖液、10 g/dL SDS、0.5 mol/L EDTA、5 mol/L NaCl、3 mol/L NaAc、異丙醇、氯仿-異戊醇(24∶1,體積比)、酚-氯仿-異戊醇(25∶24∶1,體積比)、dNTP、rTap 酶、蛋白酶 K、10×PCR Buffer、核酸染料等,以上實驗試劑均購買于北京全是金生物工程有限公司及天根化學試劑公司,本實驗所用的引物由上海桑尼生物科技有限公司設計并合成。
1.3 儀器與設備
試驗所用儀器與設備見表1。
表1 儀器與設備Table 1 Instrument and equipment
1.4 實驗方法
1.4.1 菌株耐藥性表型分析
1)菌株的活化與菌液的制備。將保藏于凍存管中的菌以體積分數1%的接種量接種于MRS液體培養基中進行活化,37℃,培養24 h。后在MRS固體培養基上劃線,37℃下培養48 h,挑取平板上單菌落于滅好菌的5 mL生理鹽水中,直至OD625值在0.16~0.2[8]之間(活菌數約為 3×108CFU/mL),用培養基稀釋500倍和900倍后制成種子液備用。
2)抗生素的制備。每種抗生素均用表2中的溶劑進行溶解,濾菌器過濾,于-80℃冰箱中保存,備用。用LSM稀釋劑將水溶性和水不溶性抗生素溶液連續稀釋至目標質量濃度的2倍和10倍。不同抗生素所需的溶解試劑及質量濃度測定范圍如表2所示。

表2 不同抗生素的質量濃度測定范圍及所需溶劑Table 2 Solvents and range of concentration for different antibiotics
3)菌株MIC的測定。最小抑菌濃度(Minimum Inhibitory Concentration,MIC)是指完全抑制菌株生長時的最小抗生素質量濃度[9]。本實驗采用肉湯稀釋法測定菌株的MIC值,具體的操作步驟參照ISO10932/IDF223[8]標準。
4)乳酸菌耐藥性評價。將測定的MIC值與表3進行比較,基于EFSA頒布的標準[10],如果MIC小于或等于臨界值時定義為敏感(S),MIC大于臨界值時定義為耐藥(R),MIC臨界值的評定標準是由歐盟食品安全局[10](EFSA)和歐盟委員會[11]共同制定的。

表3 保加利亞乳桿菌對15種抗生素耐藥性的評定標準Table 3 Evaluation criteria for the resistance of Lactobacillus bulgaricus against 15 antibiotics(μg/mL)
1.4.2 菌株耐藥性基因型分析
1)菌株DNA的提取。利用CTAB液氮凍融法[12]提取所試菌株基因組的DNA。
2)菌株DNA純度的檢測。取2 μL的DNA用微量紫外分光光度計測其質量濃度及OD260/280值,為了做后續聚合酶鏈式反應的模板測定的OD值應在1.8~2.0間。
3)菌株耐藥基因的檢測。將上一步驟提取的基因組DNA作為模板,50 μL的反應體系如下:DNA模板2 μL、10×PCR Buffer 5 μL(含 Mg2+)、dNTP 4 μL、rTap酶 0.5 μL、Primer F 、Primer R 各 1.5 μL、 去離子水35.5 μL。 PCR擴增程序為:預變性94℃5 min、30個循環:變性94℃1 min、退火1 min、延伸72℃2 min,末端延伸72℃10 min。但不同引物具有不同的退火溫度,實驗中聚合酶鏈式反應中具體所用的引物序列、目的片段長度及退火溫度見參考文獻[13]。
PCR反應結束后,將 PCR產物與 6×DNA Loading Buffer以1∶1體積比的比例均勻混合后,加樣于1 g/dL瓊脂糖凝膠點樣孔中進行電泳,并用凝膠成像儀對膠體進行分析,如果有條帶出現且PCR產物片段長度大小與已知目的片段長度相同則證明菌株中含有耐藥基因,如果無條帶出現則證明菌株中不含有耐藥基因。
2 結果與分析
2.1 菌株耐藥性的表型分析
依據表3保加利亞乳桿菌對抗生素的評定標準可知,5株保加利亞乳桿菌對15種不同抗生素的MIC值測定結果如表4所示。
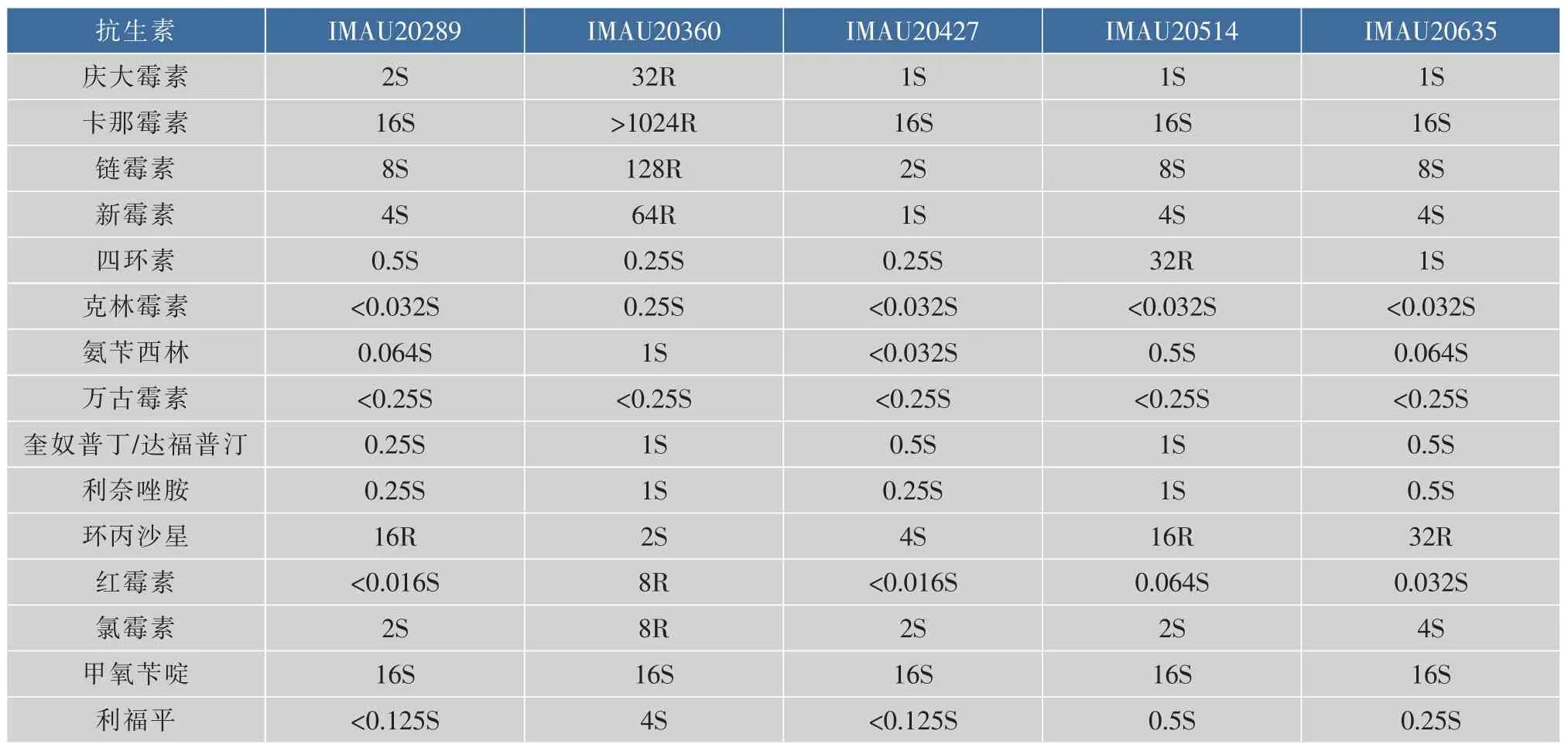
表4 5株保加利亞乳桿菌對15種抗生素的MIC測定結果Table 4 MIC determination results of 15 antibiotics for 5 Lactobacillus bulgaricus (μg/mL)
由表4可知,對于不同的抗生素5株菌均表現為不同的藥物敏感性。相較于其他抗生素,對環丙沙星的耐藥率最高。5株菌全部對利奈唑胺、氨芐西林、克林霉素、利福平、奎奴普丁/達福普汀、甲氧芐啶及萬古霉素敏感,其中對萬古霉素和甲氧芐啶的敏感性十分穩定,MIC值相同且均為<0.25 μg/mL和16 μg/mL。對四環素、卡那霉素、氯霉素、慶大霉素、鏈霉素、新霉素、紅霉素的敏感性次之。特別的菌株IMAU20360對卡那霉素、鏈霉素、紅霉素均耐藥且卡那霉素(>1 024 μg/mL)、鏈霉素(128 μg/mL)和紅霉素(8 μg/mL)的MIC值均達到了所檢測質量濃度范圍的最高值。
D'Aimmo[14]研究了乳桿菌對常見的不同抗生素的耐藥性,結果顯示所有菌株對青霉素、紅霉素、氨芐西林、利福平及克林霉素等抗生素敏感,對多粘菌素B和卡那霉素等耐藥,對鏈霉素、氯霉素、萬古霉素、四環素、新霉素和慶大霉素不同菌株分別表現出不同程度的耐藥性。先前的研究同樣也表明乳桿菌對氨芐西林和紅霉素具有高度敏感性[15]。黃曉敏[16]利用瓊脂稀釋法測定了57株保加利亞乳桿菌的MIC值,其中對克林霉素的耐藥率為0,對紅霉素、青霉素、阿莫西林的耐藥率約為3.5%~12.3%之間,對四環素、慶大霉素的耐藥率為21.1%~26.3%。周寧[17]的測定結果顯示保加利亞乳桿菌對氨基糖苷類抗生素例如:慶大霉素、卡那霉素、鏈霉素完全耐藥,且屬于固有耐藥性,而對大環內酯類抗生素中的紅霉素完全敏感。Li等[18]對19株乳酸菌的耐藥性研究表明對環丙沙星的耐藥率高達68.4%,據相關研究報道因為DNA旋轉酶上gyrA基因上的A亞基,環丙沙星通過作用于A亞基從而抑制DNA的合成和復制,最終致使細菌死亡。本實驗與D'Aimmo、黃曉敏、周寧、Li的研究結果基本一致,但也存在個別差異,例如我們觀察到菌株IMAU20360對糖肽類、大環內酯類、苯丙醇類抗生素耐藥,且在氨基糖苷類抗生素中對慶大霉素的耐藥程度較弱,可能與慶大霉素能通過細胞膜相關,該結果與周寧等人研究的結果不一致。此外我們還發現菌株IMAU20514對四環素耐藥,該發現與黃曉敏報道中的結果也存在差異。這種同種不同菌株間的差異可能是因為多藥轉運蛋白或存在缺陷細胞壁自溶系統等非特異性機制所引起的[19]。因vanX基因能夠編碼D-ala-D-ala二肽酶易使萬古霉素不能與細菌結合,所以通常認為乳桿菌對糖肽類抗生素萬古霉素為固有耐藥性[20]。但也有人認為這并不代表是物種的固有特性,因為在德氏保加利亞乳桿菌、嗜酸乳桿菌、約氏乳桿菌中可能存在可變性[21]。本研究發現所有的菌株對萬古霉素完全敏感,此結果也充分證明了保加利亞乳桿菌對萬古霉素并非是固有耐藥性。
2.2 菌株耐藥性的基因型分析
根據耐藥性分析結果,本實驗利用PCR技術對5株保加利亞乳桿菌的耐藥基因進行了檢測,檢測結果如表5所示。

表5 5株保加利亞乳桿菌中耐藥基因的檢測結果Table 5 Determination of antimicrobial resistance genes from 5 Lactobacillus bulgaricus
由表5所示,根據特異性引物利用PCR技術擴增后發現5株菌中均攜帶耐藥基因,共檢出erm(B)、vanX 、aac(6')-aph(2'')、parC、rpoB 5 種不同的耐藥基因,其中erm(B)、vanX存在于每株菌中。本研究中,菌株的耐藥表型與其基因型呈對應關系,如菌株IMAU20360對慶大霉素和紅霉素耐藥,且相對應的基因型檢測結果發現其攜帶慶大霉素耐藥基因 aac(6')-aph(2'')和紅霉素耐藥基因 erm(B)。在于濤[22]的研究中也在保加利亞乳桿菌中檢測到了慶大霉素耐藥基因 aac(6')-aph(2''),與本實驗研究結果一致。 aac(6')-aph(2'')能夠編碼雙功能修飾酶 AAC(6')-APH(2''),該酶具有磷酸轉移酶和乙酰基轉移酶兩種活性,屬于氨基糖苷類鈍化酶之一,同時影響了大部分氨基糖苷類抗生素使其具有較高的耐藥性[23]。秦宇軒等[24]在對紅霉素敏感的菌株中卻檢測到了紅霉素耐藥基因erm(B),此結果說明敏感的菌株中可能存在耐藥基因。同樣的本研究也在 敏 感 菌 株 IMAU20289、IMAU20427、IMAU20514及IMAU20635中均檢測到了紅霉素耐藥基因erm(B)和萬古霉素耐藥基因vanX,在菌株IMAU20360和菌株IMAU20514中分別檢測到針對環丙沙星的耐藥基因parC和對利福平的耐藥基因rpoB,在RNA的水平上這可能是因為該耐藥基因并未表達,或是因為調控基因表達的核苷酸序列發生了基因突變最終使開放閱讀框不被表達[25]。值得注意的是本研究從3株耐環丙沙星的菌株中均未發現相對應的耐藥基因,在對卡那霉素、四環素、新霉素、鏈霉素及氯霉素表現為耐藥的菌株中并未檢測到與其表型相對應的耐藥基因。Toomey等[26]發現在一些表型對鏈霉素存在耐藥性的菌株中并未檢測到如strA,strB、aadA和aadE等已知的鏈霉素的耐藥基因。這充分表明了耐藥基因的檢出與耐藥性之間并不存在直接關系,耐藥菌株可能由其他相關耐藥基因調控或存在其他的耐藥機制,然而這些存在的耐藥機制,相關的耐藥基因是否會轉移到不同種屬的細菌甚至是致病菌中則需要進一步的深入研究。
3 結語
本實驗對蒙古國傳統發酵乳制品中5株保加利亞乳桿菌的耐藥性進行了初步研究。結果表明,保加利亞乳桿菌對大多數的抗生素敏感,僅有菌株IMAU20360對慶大霉素、卡那霉素、鏈霉素、新霉素、紅霉素及氯霉素表現為耐藥,菌株IMAU20289、IMAU20514、IMAU20635對環丙沙星耐藥。本實驗中耐藥表型與基因型存在一定的差異。所有菌株中共檢測到了5種不同的耐藥基因,在對紅霉素和萬古霉素敏感的菌株中均檢測到了erm(B)和vanX的存在,且在部分耐藥菌株中并未檢測到與其表型相對應的耐藥基因。本實驗說明了乳酸菌可能會是耐藥基因的潛在貯存庫,這些耐藥基因潛在的轉移性可能會造成一定的食品安全隱患。所以有必要加強對乳酸菌的安全性評價并完善乳酸菌安全性評價體系,尤其是對在工業中常用的菌株,在使用前確保其安全性,為其在藥品、食品等應用中的安全性提供理論基礎。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
